

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
The human gastrointestinal tract (GIT) harbors a unique collection of microorganisms, referred to as the gut microbiota. This complex gut ecosystem includes a diverse and dynamic microbial community consisting predominantly of bacteria [1]. There exists a symbiotic and homeostatic association between the gut microbiota and the mucosal and immune cells of the intestine through direct interaction (via microbial adherence) and/or indirectly [23]. The microbiota-host interaction is indirectly mediated by metabolites, mostly derived from the metabolism of the undigestible fiber and amino acids by gut microorganisms in the colon [3]. Fermentation of aromatic amino acids such as tryptophan may be physiologically important as this generates a wide range of bioactive end products [456]. Tryptophan is an essential aromatic amino acid that can be converted by the bacterial enzyme tryptophanase into indole – a metabolite that has been associated with improved transepithelial electrical resistance by enhancing tight junction protein expression in response to pathogens [789]. Subsequently, other bacterial enzymes can convert indole into bioactive derivatives, in particular indole-3-propionic acid (IPA) [101112].
IPA is comprised of an indole ring with a propionic acid group at position 3 and its synthesis is catalyzed by the heterotrimeric enzyme phenyl lactate dehydratase which is expressed by some bacterial species, such as Clostridium sporogenes [13]. The physiological concentrations of this metabolite in the human gut are usually between 1 and 500 μM [714]. However, it has been previously reported that under pathological conditions, such as inflammatory bowel disease, the circulating IPA levels are reduced and can be restored in the remission stages of the disease [14]. Moreover, in subjects with type 2 diabetes (T2D), IPA levels are significantly lower compared to healthy subjects and this is associated with decreased plasma concentrations of glucagon-like peptide-1, a hormone secreted by intestinal enteroendocrine cells that stimulates insulin secretion [1516]. In addition, the anti-inflammatory effects of IPA have been demonstrated in mice models of colitis by a mechanism involving the activation of pregnane X receptor and inhibition of c-Jun N-terminal kinases (JNKs) [1718]. In rodent models of diet-induced obesity and diabetes, IPA supplementation restored intestinal permeability and reduced metabolic endotoxemia, weight gain, and glucose intolerance [111920]. In vitro studies using Caco-2 and T84 cell lines have also shown that IPA exerts barrier protective effects and ameliorates tumor necrosis factor-alpha-induced inflammation [1421]. Hence in vitro, animal, and clinical studies have consistently shown that IPA triggers several protective metabolic effects in diseased states which could be explored in therapeutics.
Nonetheless, studies characterizing the mechanisms of IPA in physiological conditions, particularly in the intestinal milieu are lacking. This is particularly important as it can provide further insights regarding the role of IPA in human health, thus contributing to a better understanding of its potential use as a disease risk biomarker. Therefore, the present study aimed to evaluate the effects of IPA and its precursors (tryptophan, indole, and propionate) at physiological concentrations upon the viability, integrity, differentiation, and proliferation of intestinal cells, using the Caco-2 culture model.
MATERIALS AND METHODS
Materials
4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES), trypsin-ethylenediamine tetraacetic acid (EDTA), NaCl, Tris.NaOH, Tris.HCl, KCl, Na2HPO4, NaH2PO4, CaCl2, MgSO4, MgCl2, sodium butyrate, sulforhodamine B, sodium propionate, 4-nitrophenol (Sigma-Aldrich, Madrid, Spain), IPA, indole, tryptophan (TCI, Lisboa, Portugal), Phosphate buffer solution (PBS) (GRISP, Porto, Portugal), NaOH, acetic acid, Triton X-100, dimethyl sulfoxide (DMSO) (Merck, Darmstadt, Germany), trichloroacetic acid (TCA), p-nitrophenyl phosphate (Applichem, Darmstadt, Germany), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium-bromide (MTT) and sulfonic acid (SA) (Thermo Fisher Scientific, Leiden, The Netherlands).
Caco-2 cell culture
The human epithelial Caco-2 cell line was obtained from the American-Type Culture Collection (ATCC37-HTB, Rockville, MD, USA) and was used between passage numbers 25 and 52. Caco-2 cells were grown in minimum essential medium supplemented with 15% fetal bovine serum (FBS), 25 mM HEPES, 25 mM NaHCO3, 100 units/mL penicillin, 100 μg/mL streptomycin, and 0.25 μg/mL amphotericin B (Sigma-Aldrich). Cells were incubated in a humidified atmosphere of 5% CO2 - 95% air at 37°C. The culture medium was changed every 2–3 days and cells were split every 7 days. For subculturing, cells were removed enzymatically (0.25% trypsin-EDTA, 1 min, 37°C), split 1:4, and seeded in plastic culture dishes (21 cm2; 52 mm; Orange Scientific, Braine-l’Alleud, Belgium). For the determination of cell viability, proliferation, and alkaline phosphatase activity, Caco-2 cells were seeded on 24-well plastic cell culture dishes (1.93 cm2; 16.2 mm; Orange Scientific). For permeability assays, cells were seeded in 12-well Transwell inserts (polycarbonate membrane, 0.4 μm pore size, 12 mm, Corning Costar, Corning, NY, USA), and for RNA extraction cells were seeded on 6-well microplates (9.6 cm2; 34.7 mm; Orange Scientific). Cells were used after 11 days in culture (90–100% confluence), except for proliferation assays, where cells were used after 3 days in culture (50–70% confluence), and alkaline phosphatase assays, where cells were used after 7 days in culture (90–100% confluence).
Effects of IPA and its precursors
The concentration of IPA and its precursors to be tested were chosen based on previously published works [714]. Caco-2 cells were treated for 2, 24, and/or 72 h with IPA or with its precursor’s indole, tryptophan, and propionate at 1, 10, 100, 250, or 500 μM in an FBS-free culture medium. The controls of these compounds were run in the presence of their respective solvents (absolute ethanol or an aqueous solution of 50% acetonitrile and 1% formic acid) and 0.1% concentration was used.
MTT assay
The effect of IPA, indole, tryptophan and propionate on Caco-2 cell viability was assessed by the MTT assay [22]. After treating cells with these compounds, the media was removed and each cell monolayer was further incubated at 37°C for 2 h with Hanks’ buffer (137 mM NaCl, 5 mM KCl, 0.8 mM MgSO4, 1.0 mM MgCl2, 0.33 mM Na2HPO4, 0.44 mM NaH2PO4, 0.25 mM CaCl2, 0.15 mM Tris.HCl, and 1.0 mM sodium butyrate, pH 7.4) containing 5 mg/mL MTT. After this incubation period, the MTT solution was carefully aspirated, and cells were solubilized in 200 μL DMSO and diluted fourfold. The absorbance corresponding to the purple formazan crystals produced by the capacity of mitochondrial dehydrogenases to reduce MTT was determined at 540 nm and then subtracted by the absorbance at 660 nm, which corresponds to unspecific MTT reduction. Both absorbances were measured using a SpectraMax i3x microplate reader (Molecular Devices, San Jose, CA, USA). Results were expressed in % of control.
Permeability assay
The effect of IPA, indole, tryptophan and propionate on the permeability of the Caco-2 cells monolayer was assessed by quantifying the apical-to-basal passage of fluorescein SA. Due to its ability to cross epithelia exclusively by the paracellular route, SA is considered a biomarker of intestinal permeability and also integrity [23]. Caco-2 cell monolayers were treated on the apical compartment with both IPA or its precursors and 100 mg/mL SA in an FBS-free medium. After that, the fluorescence of SA (excitation wavelength at 485 nm and emission wavelength at 530 nm) was measured on the basal compartment using a SpectraMax i3x microplate reader (Molecular Devices). Data are presented in % of control.
Ecto-alkaline phosphatase activity
The effect of IPA and its precursors on Caco-2 cell differentiation was determined by the quantification of ecto-alkaline phosphatase (ecto-ALP) activity [24], as described by Calhau et al. [25]. Briefly, after the treatment of undifferentiated cells with these compounds, the media was removed, and cells were washed once with PBS. Subsequently, each cell monolayer was incubated for 45 min at 37°C with 200 μL of Hanks’ buffer containing 5 mM of the ALP substrate p-nitrophenyl phosphate. After this period, 150 μL of this solution was collected and mixed with 600 μL of ice-cold 0.02 M NaOH. After remaining for 5 min on ice and then 10 min at room temperature, the absorbance was read at 410 nm using a SpectraMax i3x microplate reader (Molecular Devices). Ecto-ALP activity was expressed as U ALP/min/mg of protein. The protein content of the cell monolayers was determined by an EZQTM protein quantification kit (Thermo Fisher Scientific) according to the manufacturer’s instructions.
Sulforhodamine B assay
The effect of IPA, indole, tryptophan and propionate on Caco-2 cell proliferation was assessed by the sulforhodamine B assay, as described by Gonçalves et al. [26]. After treating the subconfluent cells with these compounds, 37.5 μL of ice-cold TCA was added to the medium of each well to fix cells. Following an incubation period of 1 h at 4°C protected from light, the solution was carefully aspirated, and the cells were washed five times with distilled water to remove the remaining TCA. Then, the cells were air-dried for 10 min and stained with 0.4% sulforhodamine B (dissolved in 1% acetic acid) for 15 min at room temperature. The sulforhodamine B solution was then aspirated and cells were rinsed four times with 1% acetic acid to remove the residual dye. After that, each cell monolayer was air-dried again for 30 min, solubilized with 375 μL of 10 mM Tris.NaOH solution (pH = 10.5) and orbitally shaken for 5 min at 35 rpm. After a 4-fold dilution in Tris.NaOH, the absorbance corresponding to the number of cells was determined at 540 nm using a SpectraMax i3x microplate reader (Molecular Devices). Results were expressed in % of control.
RNA extraction and real-time quantitative reverse transcription polymerase chain reaction (qRT-PCR)
After treating the Caco-2 cells with IPA and its precursors, total RNA was extracted from the cells using the Nzyol Isolation Reagent and DNase I (NZY Tech, Lisbon, Portugal), according to the manufacturer’s instructions. For cDNA synthesis, 25 ng/mL of total RNA was reverse transcribed using the NZY First-Strand cDNA Synthesis Kit, following the manufacturer’s instructions. Each individual real-time qRT-PCR was performed using: 3.6 μL of PCR water (Roche, Indianapolis, IN, USA), 0.2 μL of each primer at a final concentration of 0.2 μM (Sigma, St. Louis, MO, USA) and 5.0 μL of FastStart SYBR Green Master (Roche). Thermocycling conditions were as follows: denaturation (95°C for 10 min), amplification and quantification (95°C for 10 s, annealing temperature [AT] for 10 s and 72°C for 10 s) repeated 45 times, and a melting curve program (95°C for 10 s, [AT + 10]°C for 60 s and 97°C with a heating rate of 0.1°C/s and continuous fluorescence measurement). The primer pairs used for amplification, their sequences, and annealing temperatures are described in Table 1. PCR was carried out in duplicate for each gene using a LightCycler® 96 device (Roche, Nutley, NJ, USA). Data were analyzed using the LightCycler® 96 analysis software. Gene expression was normalized for the housekeeping gene hypoxanthine guanine phosphoribosyltransferase.
Table 1
Primer sequences and annealing temperatures used for quantitative reverse transcription polymerase chain reaction
Statistical analysis
Statistical analysis was performed using GraphPad Prism V8.0 (GraphPad Software, Inc., San Diego, CA, USA). Data are shown as mean ± SEM. “n” represents the number of replicates of at least two different days of experiments. The statistical significance of the difference between the compound and its respective control was assessed by the Student’s t-test and the statistical difference between three or more groups was evaluated by the one-way ANOVA test, followed by the Bonferroni test. Differences were considered to be significant when P < 0.05.
RESULTS
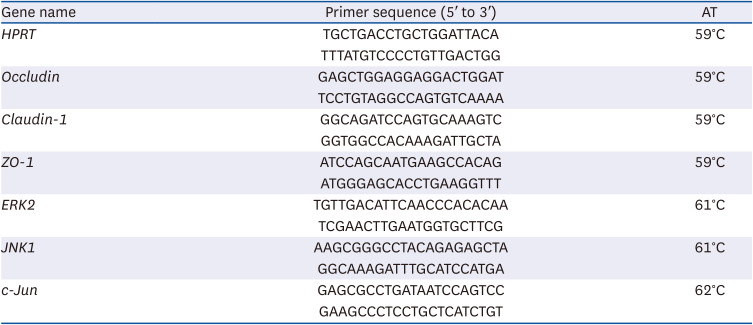
IPA and its precursors were not cytotoxic to Caco-2 cells
First, the effect of IPA and its precursors (indole, tryptophan, and propionate) on Caco-2 cell cytotoxicity using the MTT assay was analyzed. The results showed that the concentrations of IPA typically found in the intestinal lumen (1–500 μM) [714], did not induce Caco-2 cells cytotoxicity after treatment periods of 2, 24, and 72 h (Fig. 1A-C). Similar results were obtained when the cells were treated with IPA precursors at 1 to 500 μM and for 2, 24, and 72 h (Fig. 1A-C). In the following experiments (monolayer integrity), cells were treated with IPA and its precursors at 500 μM for 24 h.
Fig. 1
Effect of IPA, indole, tryptophan, and propionate at 1, 10, 100, 250, and 500 μM on the viability of Caco-2 cells, following 2 (A), 24 (B), and 72 h treatment periods. Cell viability was determined by the MTT assay. Results are shown as % of control. Data are expressed as mean ± SEM (n = 8).
Ctrl, control; IPA, indole-3-propionic acid; ACN, acetonitrile; FA, formic acid.
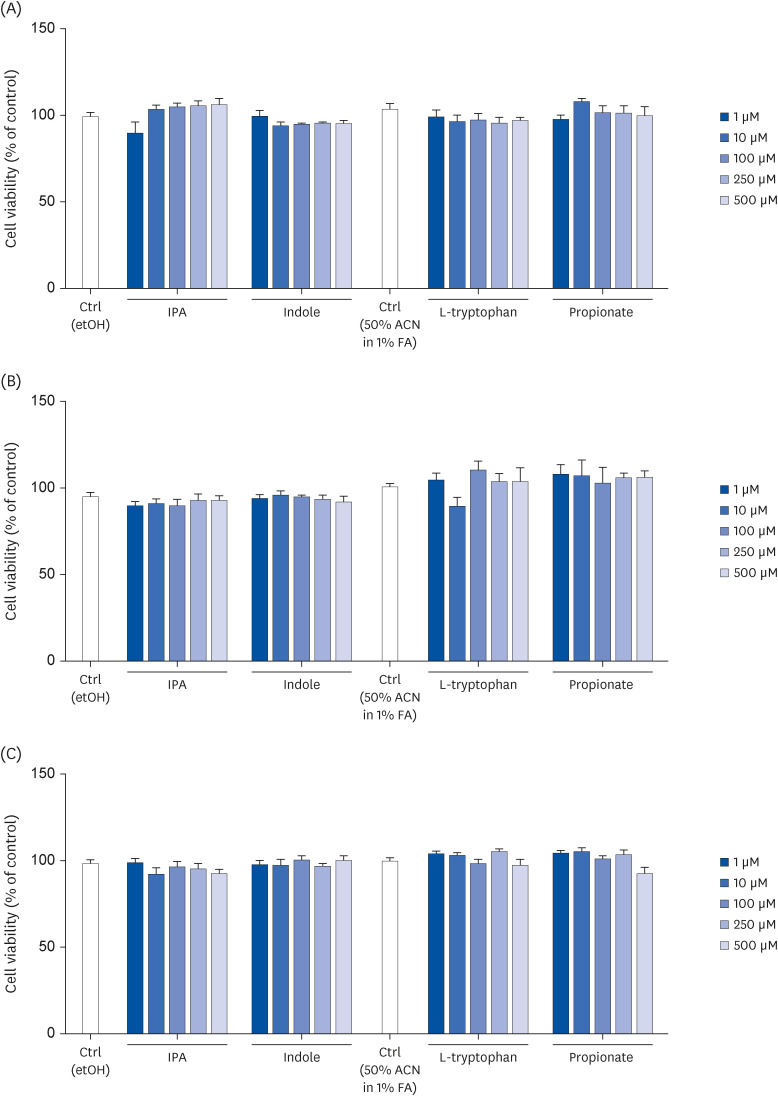
Tryptophan and propionate increased the monolayer integrity of Caco-2 cells
To assess if IPA and its precursors change the monolayer integrity of Caco-2 cells, the apical to the basolateral passage of fluorescein SA, after 24 h of treatment, was measured. When compared to control conditions, tryptophan (P = 0.0281) and propionate (P = 0.0465) significantly decreased SA passage suggesting that both compounds increase cell monolayer integrity (Fig. 2).
Fig. 2
Effect of IPA, indole, tryptophan, and propionate at 500 μM on Caco-2 cells monolayer integrity after a 24 h treatment period. The cell monolayer integrity was determined by assessing the passage of sulfonic acid from the apical to the basolateral compartment. Results are shown as % of control. Data are expressed as mean ± SEM (n = 4).
Ctrl, control; etOH, ethanol; IPA, indole-3-propionic acid; ACN, acetonitrile; FA, formic acid.
*P < 0.05 vs. controls.
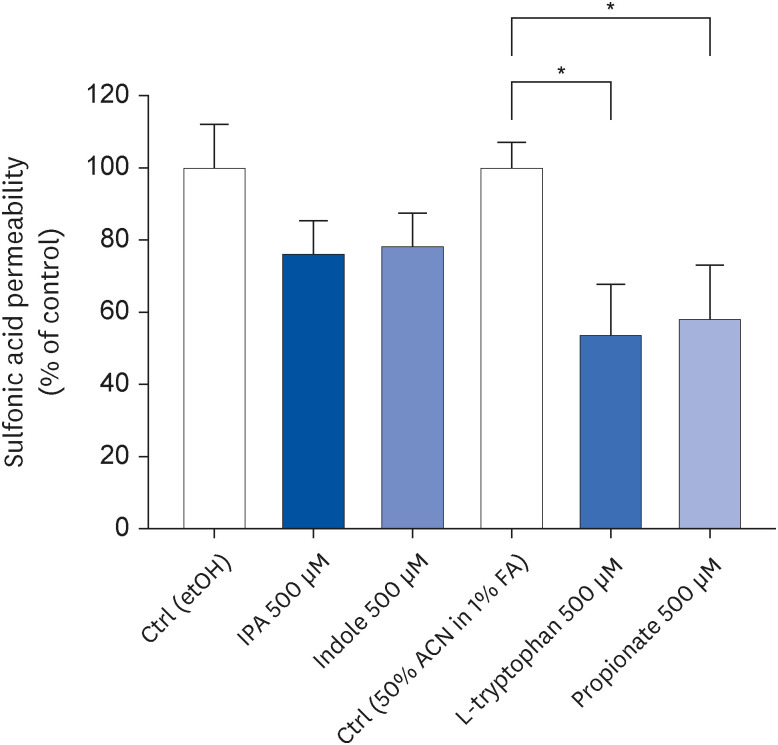
Tryptophan and propionate decreased claudin-1 expression
To assess if IPA precursors-induced changes in monolayer integrity were related to tight junction proteins, the gene expression of claudin-1, occludin, and zonula occludens-1 (ZO-1) was analyzed by qRT-PCR. After 24 h, tryptophan and propionate decreased claudin-1 expression 1.93-fold (P = 0.0021) and 2.49-fold (P = 0.0009), respectively, when compared to control conditions (Fig. 3A). Occludin and ZO-1 expression were not altered by these two compounds (Fig. 3A and B). Neither IPA nor indole affected tight junction gene expression (Fig. 3A-C).
Fig. 3
Effect of IPA, indole, tryptophan, and propionate at 500 μM on claudin-1 (A), occludin (B), and ZO-1 (C) gene expression in Caco-2 cells after 24 h. Relative expression of these genes was evaluated by quantitative reverse transcription polymerase chain reaction. Results obtained were normalized for the housekeeping gene HPRT and data are expressed as mean ± SEM (n = 3–4).
AU, arbitrary units; HPRT, hypoxanthine guanine phosphoribosyltransferase; Ctrl, control; etOH, ethanol; IPA, indole-3-propionic acid; ACN, acetonitrile; FA, formic acid; ZO-1, zonula occludens-1.
**P < 0.01 and ***P < 0.001 vs. controls.
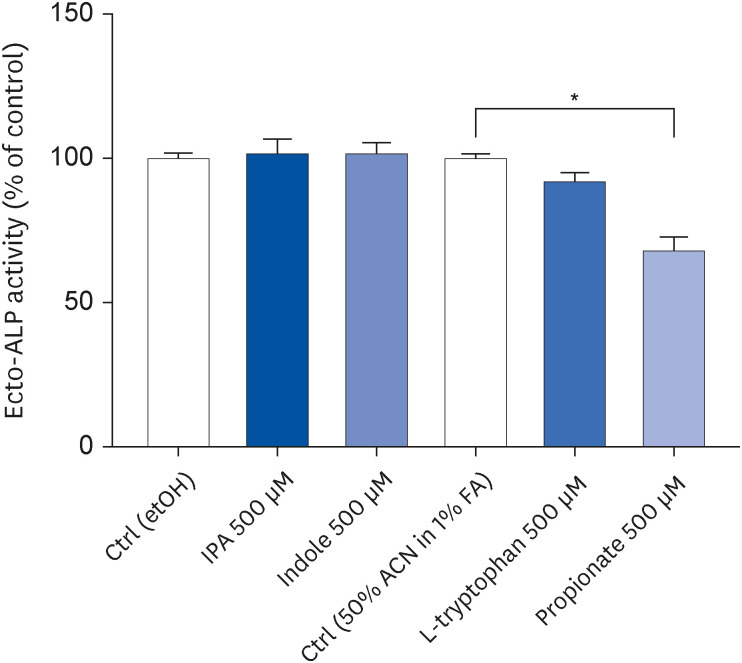
Propionate decreased Caco-2 cell differentiation
In the next step, the effect of IPA and its precursors on the differentiation of Caco-2 cells was evaluated by measuring ecto-ALP activity. As shown in Fig. 4, this enzyme’s activity decreased after treating confluent cells during the last 72 h of differentiation with 500 μM propionate (P = 0.0138) but not with 500 μM IPA and its other precursors.
Fig. 4
Effect of IPA, indole, tryptophan, and propionate at 500 μM during the last 72 h of Caco-2 cell differentiation. Cell differentiation was determined by measuring ecto-ALP activity. Results are shown as % of control. Data are expressed as mean ± SEM (n = 8).
etOH, ethanol; ALP, alkaline phosphatase; Ctrl, control; IPA, indole-3-propionic acid; ACN, acetonitrile; FA, formic acid.
*P < 0.05 vs. controls.
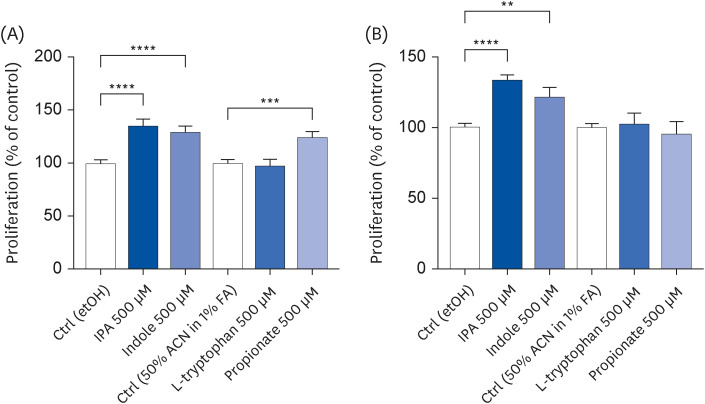
IPA-induced Caco-2 cell proliferation
The effect of IPA and its precursors on the proliferative capacity of Caco-2 cells was evaluated using a sulforhodamine B assay. After a 2 h treatment, IPA, indole, and propionate increased the cell proliferation rate by 35% (P < 0.0001), 29% (P < 0.0001), and 25% (P = 0.0003), respectively, compared to control conditions (Fig. 5A). An effect of similar magnitude was also observed for IPA and indole after a 24 h treatment (increase of 34% [P < 0.0001] and 22% [P = 0.0021], respectively) (Fig. 5B). On the other hand, the effect of propionate on cell proliferation was no longer observed after 24 h (Fig. 5B). Tryptophan did not alter the proliferation of Caco-2 cells (Fig. 5A and B).
Fig. 5
Effect of IPA, indole, tryptophan, and propionate at 500 μM on Caco-2 cell proliferation after a 2 h (A) and 24 h (B) treatment period. Cell proliferation was determined by sulforhodamine B assay. Results are shown as % of control. Data are expressed as mean ± SEM (n = 11–24).
Ctrl, control; etOH, ethanol; IPA, indole-3-propionic acid; ACN, acetonitrile; FA, formic acid.
**P < 0.01, ***P < 0.001, and ****P < 0.0001 vs. controls.
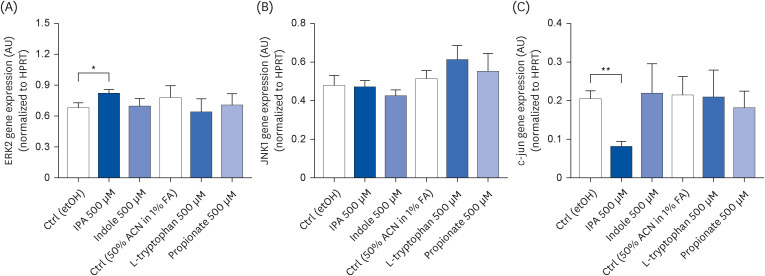
IPA-induced changes in extracellular signal-regulated kinase 2 (ERK2) and c-Jun expression
To confirm if the IPA-induced changes in the proliferative capacity of Caco-2 cells were associated with the modulation of the signaling pathways controlling cell proliferation, the gene expression of ERK2, c-Jun N-terminal protein kinase 1 (JNK1) and c-Jun was analyzed by qRT-PCR. As shown in Fig. 6A and C, after a 24 h treatment, IPA (500 μM) increased ERK2 expression by 1.20-fold (P = 0.0319) and decreased c-Jun expression (P =0.0021) 2.48-fold when compared with control conditions. These effects were not accompanied by changes in JNK1 expression (Fig. 6B). None of the IPA precursors affected the gene expression of ERK2, JNK1, and c-Jun (Fig. 6A-C).
Fig. 6
Effect of IPA, indole, tryptophan, and propionate at 500 μM on Caco-2 cells expression of ERK2 (A), JNK1 (B), and c-jun (C) after 24 h. Relative expression of these genes was evaluated by qRT-PCR. Results obtained were normalized for the housekeeping gene HPRT and data are expressed as mean ± SEM (n = 4).
ERK2, extracellular signal-regulated kinase 2; AU, arbitrary units; HPRT, hypoxanthine guanine phosphoribosyltransferase; Ctrl, control; etOH, ethanol; IPA, indole-3-propionic acid; ACN, acetonitrile; FA, formic acid; JNK1, c-Jun N-terminal kinase.
*P < 0.05 and **P < 0.01 vs. controls.
DISCUSSION
The GIT is an important habitat of a diverse and highly dynamic microbial community known as the gut microbiota [1]. This community is mainly composed of bacteria and is highly influenced by the environment, especially diet [27]. While a low-fiber diet decreases the abundance of commensal bacteria, contributing to the degradation of the epithelial mucus and increased inflammation, a diet rich in fiber has the opposite effect and increases the synthesis of beneficial metabolites [27]. The gut microbiota is involved in critical physiological host functions, including the degradation of indigestible macronutrients and complementing the activity of the host’s gut enzymes [5]. The products of protein digestion, peptides, and amino acids can be used by bacteria for de novo biosynthesis of proteins, or, importantly, be metabolized into several small bioactive compounds [6]. The latter are essential regulators of signal transduction pathways, barrier function, and inflammation in host intestinal epithelial cells [628]. Tryptophan-derived metabolites have recently attracted a great deal of interest. Most of this essential amino acid is metabolized by intestinal cells through the kynurenine and serotonin pathways [4]. The remaining tryptophan is further metabolized, through the indole pathway, in the large bowel by the gut microbiota into indole and its derivatives, with some of them acting as aryl hydrocarbon receptor (AhR) ligands [4]. This is the case with IPA, which has been linked with a myriad of health benefits, including antioxidant, anti-inflammatory and neuroprotective effects, and has even been associated with a decreased risk of T2D [4729]. Although some metabolic effects of this bioactive compound have been reported, particularly in pathological conditions [162030313233], there remains a need for detailed mechanistic insights regarding its interaction with the host in intestinal physiology. Taking this into consideration, the present work adds information to this field by assessing the effects of IPA and its precursors (tryptophan, indole, and propionate) at physiological concentrations upon the viability, integrity, differentiation, and proliferation of Caco-2 cells. This cell model was chosen due to its wide application in the study of the mechanisms and effects of metabolites of gut microbiota on the intestinal epithelial barrier function [34].
In the present study, it was demonstrated that IPA induced Caco-2 cell proliferation and this effect was associated with an increase and a decrease in ERK2 and c-Jun expression, respectively. ERK2 belongs to the mitogen-activated protein kinase family that is involved in cell proliferation [35], whereas c-Jun, a transcription factor regulated by JNK phosphorylation, is mainly related to apoptotic pathways [36]. These results are in agreement with previous studies demonstrating that, in the absence of growth factors, AhR ligands (including IPA) may activate ERK1/2 thereby increasing the cell proliferation and survival of hepatocytes and immune cells [37]. Although indole also increased the cell proliferation rate of Caco-2 cells, its effect was lower than that of IPA and did not involve ERK signaling pathway. This may be explained by the fact that indole is not considered an AhR ligand, and that it might stimulate cell proliferation through a different pathway [38]. Also, both IPA and indole appear to have no effect on the viability and differentiation of Caco-2 cells. Although the absence of effect of IPA on cell viability observed in this study is in accordance with previous research [39], to our knowledge this is the first time that the effect of IPA and indole in Caco-2 cell differentiation has been reported. Both IPA and indole did not appear to alter cellular integrity in Caco-2 cells. This minor impact of IPA and indole in cellular integrity compared to previous studies can be justified by a difference in the experimental conditions. While previous studies were performed after an inflammatory stimulus, such as lipopolysaccharides or high-fat diets [9143132], in the present study, experiments were performed under physiologic conditions, demonstrating a discrepant effect of IPA and indole in normal versus pathologic settings.
As for the other tested compounds, tryptophan did not show any effect on the viability, proliferation, and differentiation of Caco-2 cells. Previous studies using different cell lines corroborate the lack of effect of tryptophan on cell proliferative capacity [4041]. Concerning differentiation, a recent study published by Pham et al. [42] has shown that tryptophan induces the differentiation of bone marrow-derived mesenchymal stromal cells. However, the ability of these cells to maintain a permanent undifferentiated phenotype challenges the comparison with the findings obtained in the present work. Regarding propionate, this short-chain fatty acid did not affect cell viability but increased cell proliferation (after 2 h) and differentiation (after 72 h). The evidence concerning this result shows that propionate may exhibit different effects in healthy and disease conditions, increasing cell proliferation in a healthy colonic mucosa and decreasing it in an inflamed one [4344]. However, in the present work, no effect was observed on cell proliferation after a 24 h treatment. Fu et al., [45] even observed that after treating Caco-2 cells with propionate for 48 h, proliferation decreased and differentiation rate increased. These discrepant results regarding the proliferative capacity of propionate might be explained by the activation of G protein-coupled receptor 43 (GPR43), which has been demonstrated to significantly reduce Yes-associated protein 1 (YAP1)-mediated phosphorylation in a time-dependent manner[46]. Apart from the differences in cell properties between propionate and tryptophan, both compounds effectively increased cell integrity. Indeed, tryptophan has previously been shown to improve intestinal barrier function under physiologic [47] and lipopolysaccharide-induced inflammatory settings [48]. On the other hand, propionate appears to show conflicting results on Caco-2 cell integrity under normal and inflammatory conditions, i.e. either protective or non-protective barrier effects [4950]. These dual and opposite effects may be related to the lack of robust studies that specifically detect the effect of propionate without the confounding effect of other short-chain fatty acids, especially butyrate, which accounts for most of the beneficial effects seen with microbiota-related metabolites [51]. To understand the impact of tryptophan and propionate on cell monolayer integrity, the expression profile of tight junction proteins, particularly occludin, claudins, and zonula occludens, was assessed. The results indicate that tryptophan and propionate reduced claudin-1 gene expression. Previous studies have shown that claudin-1 expression is regulated by cytokines [52], which may explain why this protein is overexpressed in some inflammatory conditions [5354] and reflects a possible feedback loop to counteract the impact on gut integrity. Thus, we hypothesize that since experiments were performed in the absence of an inflammatory stimulus and tryptophan and propionate induce anti-inflammatory effects [5556], claudin-1 is not essential to maintain epithelial integrity. Nonetheless, the lack of effect on occludin and ZO-1 expression after tryptophan and propionate treatment was also observed by Chen et al.(in Caco-2 cells) [48] and Zhang et al. (in intestinal segments of pigs) [57], respectively.
In conclusion, the results of this study suggested that the pro-proliferative effect of IPA in intestinal epithelial cells depended primarily on the IPA molecule rather than on its precursor molecules: tryptophan, indole, or propionate. Notwithstanding this, the physiological levels of the IPA precursors also appear to have an important role in maintaining the homeostasis of intestinal cells and barrier function. Therefore, gut microbiota composed of IPA-producing bacteria will certainly contribute to improving intestinal health.
 XML Download
XML Download