

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Exercise-induced fatigue is a phenomenon in which the ability of the body to work is reduced due to excessive exercise. Energy substances are gradually depleted when the body performs long-term, high-intensity exercise. Metabolites accumulate; the electrolyte metabolism is disordered; free radicals are generated in large quantities, resulting in exercise-induced fatigue. The mitochondria, as the center of cellular energy generation, produce excessive reactive oxygen species while oxidatively decomposing energy substances. These substances change the fluidity and permeability of the mitochondrial membrane, so the energy metabolism enzymes are in an abnormal state. The mitochondrial function is disordered; ATP production is reduced; skeletal muscle is fatigued and damaged [1]. Appropriate exogenous antioxidant supplementation can effectively improve the body's ability to generate energy and scavenge oxygen free radicals, protect the mitochondrial function, and relieve exercise-induced fatigue [2]. Current research has focused on compounds containing phenolic hydroxyl groups with strong antioxidant capacity [34].
Resveratrol is a polyphenolic compound found in plants, such as knotweed, peanuts, grapes, and veratrole [5]. Resveratrol has antioxidant, anti-inflammatory, calcium ion regulation, phytoestrogen, and anti-tumor properties [6]. Several studies have found that resveratrol can improve mitochondrial quality by regulating mitochondrial biosynthesis, fusion, fission, and autophagy, thereby preventing and treating metabolic and neurological diseases. In this study, a rat model of exercise-induced fatigue was constructed through moderate- and high-intensity exercise. The effects of supplementing resveratrol on mitochondrial energy metabolism and exercise endurance in rats were investigated.
MATERIALS AND METHODS
Animals and experimental design
The experiment and feeding protocol complied with the national health guidelines and were reviewed and approved by the Experimental Animal Welfare Ethics Committee of Yangzhou University (202012010). Forty-eight Sprague-Dawley male rats (6–8 wk, 234.5 ± 14.5 g) were purchased from the Hangzhou Medical College (SCXK [ZHE] 2019-0002). The rats were raised in a 22–25°C environment with a moderate humidity of 50 ± 5%, 12/12-h light/dark cycle, and free access to food and water.
After one week of adaptive feeding and swimming practice, the rats were randomly divided into four groups (each group, n = 12): the control group (C), resveratrol group (R), exercise group (E), and exercise and resveratrol group (ER). The exercise program referred to previous research [78]. Rats were set to swim for 60 min with 5% of the weight daily and trained six days a week for 6 wk. The exercise time was between 8:00–11:00 a.m. daily. The water temperature was 32 ± 2°C, and the water depth was 1.5 times the body length of a rat.
Administration method
Referring to the studies of Alway et al. [9], Polley et al. [10], and Gocmez et al. [11], the dose of resveratrol administration was determined to be 50 mg/kg by combining human and rat dose conversions. Resveratrol was dissolved in dimethyl sulfoxide (DMSO) and diluted with sterile deionized water to a DMSO concentration of 2% and a resveratrol concentration of 25 mg/mL. Resveratrol was administered by gavage one hour after exercise, six times a week for six weeks. Groups ER and R were gavaged at 50 mg/kg, and groups E and C were gavaged with the same volume of 2% DMSO solution.
Sample collection
Twenty-four hours after their last exercise, the rats were anesthetized, and blood was collected through the abdominal aorta. Gastrocnemius muscle tissue was excised, frozen quickly in liquid nitrogen, and then frozen in a −80°C refrigerator for measurement.
Main reagents and detection methods
Resveratrol (purity ≥ 99%) was purchased from Shanghai McLean Company. Blood urea nitrogen (BUN), creatine kinase (CK), total superoxide dismutase (T-SOD), malondialdehyde (MDA) ATPase, succinate dehydrogenase (SDH), cytochrome-c oxidase (COX), and citrate synthase (CS) activities assay kits were purchased from Nanjing Jiancheng Bioengineering Institute. TRIzol reagent was purchased from Ambion Company, the ReverTra Ace qPCR RT Kit was purchased from TOYOBO Company (Osaka, Japan), and the SYBR Green Master (Rox) kit was purchased from Roche Company (Indianapolis, IN, USA).
Real-time quantitative polymerase chain reaction (RT-qPCR) analysis
The total RNA from the skeletal muscle was extracted with TRIzol and measured using an ultra-micro spectrophotometer. The total RNA was reverse transcribed into cDNA using a ReverTra Ace qPCR RT Kit. qPCR was performed using an SYBR Green Master Mix. The primer sequence and synthesis were provided by Nanjing Ruizhen Biotechnology Co., Ltd. (Nanjing, China). Fold changes of the target gene expression were calculated using the relative quantification method (2−ΔΔCT). The primer sequences of the target gene are exhibited in Table 1 (CT indicates the number of threshold cycles in the RT-PCR experiment).
Table 1
Amplification primer sequences of the target gene

RESULTS
Changes in the indices related to exercise fatigue in each group of rats
As listed in Table 2, compared with group C, the BUN content (P < 0.05) and CK activity (P < 0.05) in the plasma of group E were increased significantly. Compared with group E, the BUN content (P < 0.01) and CK activity (P < 0.05) decreased significantly in the plasma from group ER. This data indicated that exercise intensity caused fatigue in the rats, and a model of exercise-induced fatigue was established. In addition, resveratrol supplementation could reduce the BUN and CK activity significantly in rats and improve their anti-fatigue ability.
Table 2
Changes in BUN, CK, T-SOD, and MDA indices of rats in each group
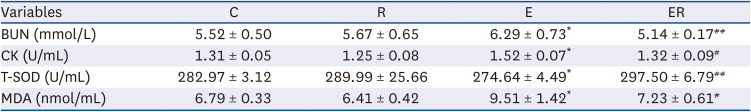
BUN, blood urea nitrogen; CK, creatine kinase; T-SOD, total superoxide dismutase; MDA, malondialdehyde; C, control group; R, resveratrol group; E, exercise group; ER, exercise and resveratrol group.
Note: * indicates P < 0.05 compared to group C; ** indicates P < 0.01 compared to group C; # indicates P < 0.05 compared with group E; ## indicates P < 0.01 compared with group E.
Compared with group C, the T-SOD activity in the skeletal muscle of group E (P < 0.05) was lower, and the MDA content was significantly higher (P < 0.05) (Table 2). T-SOD activity (P < 0.01) was significantly higher, and the MDA content (P < 0.05) was significantly lower in skeletal muscle in group ER compared to group E (Table 2). The results suggest that 5% weight-bearing swimming for 60 min can increase the level of oxidative stress in rats’ skeletal muscle, and resveratrol supplementation can reduce the oxidative stress caused by moderate-and high-intensity exercise, enhance oxygen-free radical scavenging, and improve the fatigue state.
Changes in the mitochondrial function in the rat skeletal muscle in each group
Changes in the ATPase activity in the rat skeletal muscle in each group
The activities of Ca2+-Mg2+-ATPase (P < 0.01) and Na+-K+-ATPase (P < 0.05) were significantly lower in the skeletal muscle of group E than in Group C. Compared with group E, Ca2+-Mg2+-ATPase (P < 0.01) and Na+-K+-ATPase activities (P < 0.05) increased significantly in the skeletal muscle of group ER (Fig. 1). These data suggest that resveratrol supplementation can reduce Ca2+ overload, protect the mitochondrial structure and function, and positively regulate Na+ and K+ operation to maintain the cell membrane integrity.
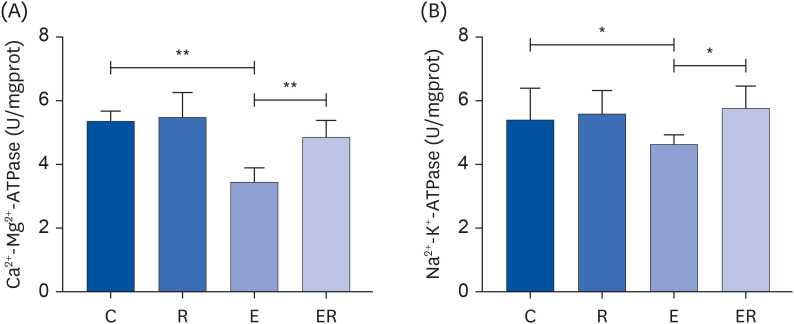
Enzyme changes related to mitochondrial energy metabolism in the skeletal muscle of rats in each group
As shown in Fig. 2A, the SDH activity (P < 0.05) was significantly lower in the skeletal muscle of group E than in group C. The SDH activity (P < 0.05) in group ER was significantly higher than in group E. Hence, resveratrol supplementation can improve the state of mitochondrial damage and enhance mitochondrial energy production.
Fig. 2
Changes in the (A) SDH, (B) CS activity, and (C) COX content in the skeletal muscle.
C, control group; R, resveratrol group; E, exercise group; ER, exercise and resveratrol group; SDH, succinate dehydrogenase; CS, citrate synthase activity; COX, cytochrome-c oxidase.
Note: *P < 0.05.
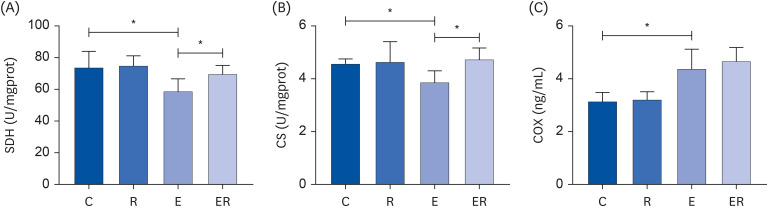
The CS activity (P < 0.05) was significantly lower in the skeletal muscle of rats in group E than in group C. The CS activity (P < 0.05) of group ER was significantly higher than that of group E (Fig. 2B). These data suggest that resveratrol supplementation can improve the CS activity and improve the mitochondrial energy production function.
Furthermore, the COX content (P < 0.05) was significantly higher in the skeletal muscle in group E than in group C. The COX activity was slightly higher in group ER than in group E, but the difference was not statistically significant (Fig. 2C).
Gene expression changes related to mitochondrial biosynthesis in rat skeletal muscle in each group
Sirtuin 1 (SIRT1) mRNA expression was significantly higher in groups E and R than in group C (P < 0.01 and P < 0.01, respectively). The mRNA expression of SIRT1 in group ER was significantly higher than in group E (P < 0.05). These results suggest that exercise and resveratrol supplementation increase SIRT1 gene expression (Fig. 3A).
Fig. 3
mRNA expression level of (A) SIRT1, (B) PGC-1α, and (C) NRF-1 in the skeletal muscle.
C, control group; R, resveratrol group; E, exercise group; ER, exercise and resveratrol group; SIRT1, sirtuin 1; PGC-1α, peroxisome proliferator-activated receptor gamma coactivator-1α; NRF-1, nuclear respiratory factor 1.
Note: *P < 0.05, **P < 0.01.
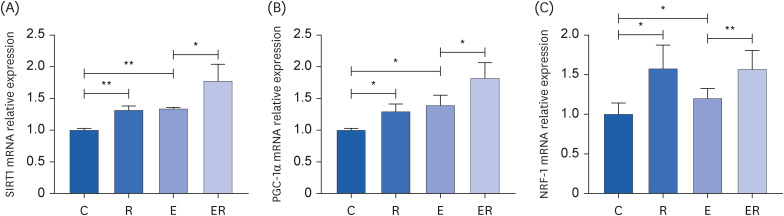
The mRNA expression of peroxisome proliferator-activated receptor gamma coactivator-1α (PGC-1α) was significantly higher in groups E (P < 0.05) and R (P < 0.05) than in group C. The mRNA expression of PGC-1α was significantly higher in group ER (P < 0.05) than in group E (Fig. 3B). These data indicate that exercise and resveratrol supplementation could increase PGC-1α mRNA expression and promote mitochondrial production.
The mRNA expression levels of nuclear respiratory factor 1 (NRF-1) were significantly higher in groups E (P < 0.05) and R (P < 0.05) than in group C. The mRNA expression of NRF-1 (P < 0.01) was significantly higher in group ER than in group E (Fig. 3C). These results suggest that exercise or resveratrol supplementation could increase NRF-1 mRNA expression and promote mitochondrial production.
DISCUSSION
The BUN and plasma CK levels are classic indicators of the body function and fatigue. The present study found that swimming with a 5% weight for 60 min significantly increased the BUN and plasma CK values, and the rats were significantly fatigued. Supplementing with resveratrol could reduce these two indicators significantly and improve exercise-induced fatigue.
During high-intensity exercise, the body produces many free radicals, such as hydroxyl and superoxide anion free radicals, called reactive oxygen species (ROS). ROS production and accumulation can cause oxidative stress injury and fatigue in skeletal muscle and other tissues [1213]. SOD is the primary mitochondrial antioxidant enzyme that can accelerate ROS scavenging [14]. MDA is a product of free radicals attacking unsaturated fatty acids in cells leading to lipid peroxidation, and is a typical indicator reflecting the degree of lipid peroxidation [15]. The MDA and SOD levels are critical for measuring the degree of cellular oxidative stress and the antioxidant capacity [2].
This paper reports that T-SOD activity in the gastrocnemius muscle of group E decreased significantly, and the MDA content increased significantly. The fatigue results from increased oxidative stress in the body after exercise and the excessive production of free radicals. The T-SOD activity was significantly higher, and the MDA content was significantly lower than group E. Hence, supplementing resveratrol after exercise can accelerate free radical scavenging, improve the body’s antioxidant capacity, and reduce lipid peroxidation, thereby relieving exercise-induced fatigue.
ATPase is a critical active protease in biological membranes, among which Ca2+-Mg2+-ATPase and Na+-K+-ATPase play important roles in maintaining transmembrane ion movement, energy metabolism, and information transduction [1617]. During excessive exercise, the increase in oxygen free radicals can damage the ATPase structure when the disulfide sulfhydryl group in the active center is oxidized. Concurrently, the body’s increased metabolic demand leads to insufficient blood and oxygen supply to tissues, weakening mitochondrial oxidative phosphorylation, enhancing the anaerobic metabolism, reducing the ATP synthesis rate, and further decreasing the ATPase activity.
The present experimental results show that Ca2+-Mg2+-ATPase and Na+-K+-ATPase activities in the skeletal muscle of group E were reduced significantly by moderate- and high-intensity exercise. Fernández et al. [18] reported that the activity of Na+-K+-ATPase was reduced significantly after a single exercise session. No study has shown how Ca2+-Mg2+-ATPase is changed after fatigue. In contrast, resveratrol supplementation significantly increased the activities of these two enzymes in group ER. Thus, moderate-to-high-intensity exercise significantly reduces ATPase activity in skeletal muscle, while resveratrol supplementation can improve the degree of mitochondrial oxidative stress, increase the ATPase activity, and promote mitochondrial energy metabolism.
SDH is a part of the mitochondrial respiratory chain complex II, and participates in the citric acid cycle in the mitochondria, oxidizing succinate to fumarate and promoting the generation of ATP [19]. CS is also an essential catalytic enzyme involved in the tricarboxylic acid cycle, and CS activity is often used to indicate the mitochondrial content and functional status [2021]. COX acts as an electron shuttle in the inner mitochondrial membrane, continuously transferring electrons from respiratory complex III to complex VI [22]. When COX is reduced, the mitochondrial respiratory chain is disrupted, electron transport is impaired, and ATP production is reduced. The experiments found that medium- and high-intensity swimming training decreased the number and vitality of SDH, CS, and COX in rats, while resveratrol supplementation could improve these phenomena significantly.
Sun et al. [23] reported that exercise, combined with resveratrol, can increase the ATPase, CS, and COX activities in obese mice. Vincenzi et al. [24] examined a type I citrullinemia rat model. They reported that resveratrol supplementation could inhibit the reduction of SDH and COX in the cerebral cortex induced by citrulline and promote energy metabolism. The present results are consistent with these findings, indicating that resveratrol supplementation can increase the ATPase, CS, and SDH activities in exercise-induced fatigued rats and improve the degree of exercise fatigue in rats.
Mitochondrial biosynthesis is a complex process. PGC-1α is a significant transcriptional activator that regulates the mitochondrial energy metabolism and muscle function. Its specific overexpression in skeletal muscle increases mitochondrial biogenesis, thereby enhancing exercise capacity [25]. SIRT1 is located upstream of PGC-1α and is a sirtuin family member. It participates in various cellular responses in vivo and can regulate mitochondrial biosynthesis [2627]. NRF-1, a transcription factor located downstream of PGC-1α, promotes the transcription of key mitochondrial enzymes and plays a critical role in mitochondrial biogenesis [28]. During exercise, the metabolic changes in the body will activate the partial enzyme system to regulate the phosphorylation and deacetylation of PGC-1α, causing mitochondrial biogenesis and improving the energy metabolism [29].
In recent years, most studies have proven that resveratrol, as a naturally occurring plant polyphenol antioxidant, can alter the mitochondrial energy metabolism by regulating mitochondrial mass. Chuang et al. [30] reported that resveratrol could help regulate mitochondrial biogenesis in an experimental epilepsy model by activating the PGC-1α signaling pathway to increase the NRF-1 and mtDNA levels. Wang et al. [31] showed that resveratrol supplementation could promote the expression of PGC-1α, NRF-,1 COX IV, and other proteins that regulate mitochondrial biogenesis in type 2 diabetic mice, reduce the PTEN-induced kinase 1 (PINK1) and Parkin autophagy-related protein expression, and improved the mitochondrial mass.
Resveratrol has preventive and therapeutic effects on diabetic muscle atrophy [31]. In a study of aging mice, Muhammad and Allam [32] reported that resveratrol supplementation and exercise can promote PGC-1α gene overexpression and improve the degree of cellular aging. Vargas-Ortiz et al. [26] showed that several exercise sessions activate SIRT1 and SIRT3 that, together with the biogenesis and mitochondrial oxidative function, jointly activate ATP production and the mitochondrial antioxidant function. Menzies et al. [29] reported that voluntary exercise and resveratrol treatment induced mitochondrial biogenesis in a SIRT1-independent manner. On the other hand, a SIRT1-dependent synergistic effect was evident when resveratrol and exercise were combined, leading to enhanced translocation of PGC-1α and SIRT1 to the nucleus and stimulation of mitochondrial biogenesis. The study results also showed that exercise and resveratrol supplementation could promote SIRT1, PGC-1α, and NRF-1 gene expression. When exercise was combined with resveratrol, a more apparent synergistic effect appeared, indicating that supplementation with resveratrol could activate the SIRT1-PGC-1α-NRF-1 pathway and promote mitochondrial biogenesis.
Resveratrol supplementation could promote mitochondrial biogenesis by activating the SIRT1-PGC-1α-NRF-1 pathway, increase the activity of enzymes related to the mitochondrial energy metabolism, improve the body’s antioxidant capacity, improve mitochondrial energy metabolism, and promote recovery from exercise-induced fatigue.
 XML Download
XML Download