

 PDF
PDF Citation
Citation Print
Print



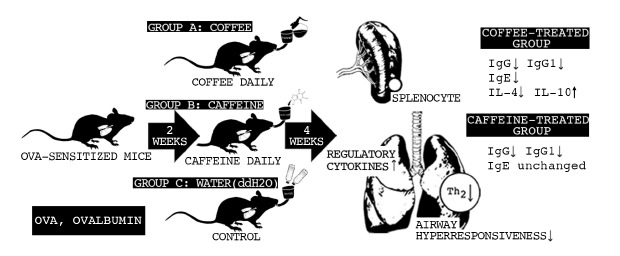
INTRODUCTION
Coffee is one of the most popular beverages consumed worldwide. As a complex chemical mixture composed of more than 1,000 components [1], coffee possess many health benefits [2], including reducing the risks of various cancers [345678], liver and gallstone diseases [391011], metabolic syndrome [12], coronary morbidity [1314], depression [1516], hyperuricemia [17], and kidney stones [18].
Coffee consumption is related with suppressed allergic reaction by metabolites associated with anti-inflammatory and immunomodulatory activities [19]. Caffeine (1, 3, 7-trimethylxanthine), the most well-known bioactive component contained in coffee, is an adenosine receptor antagonist and a phosphodiesterase inhibitor [20]. Like theophylline, it belongs to the group of methylxanthines, which is used as a bronchodilator in the clinical setting. Due to the industrialized environment and air pollution, airway hypersensitivity is becoming a rising concern worldwide. According to current human meta-analysis studies, caffeine improves airway function in asthmatic patients [19], and poses anti-inflammatory effects which influence both innate and adaptive immunity [21]. Additionally, caffeine is a pharmaceutical product either in tablet or intravenous form, prescribed for particular medical indications such as apnea of prematurity. Based on available data, how caffeine influences specific cytokines associated with allergic reactions of the respiratory system have not been clarified. Furthermore, whether coffee enhances immunomodulatory effects due to its natural form as a chemical compound containing other bioactive substances besides caffeine is unknown. We hypothesize that coffee can effectively reduce airway hypersensitivity via anti-inflammatory and immunomodulatory pathways. Most studies regarding health benefits associated with caffeine or coffee ingestion are based on observational studies. In this study, we investigate the effect of coffee and caffeine on airway hyperresponsiveness and allergic reactions through analysis of cytokine assays in ovalbumin (OVA)-sensitized mice. Through our experiment findings, we present the cytokine profile and demonstrate the direct impact of coffee on the respiratory system, in hopes of encouraging a novel and effective treatment for airway hypersensitivity.
MATERIALS AND METHODS
Animals
Six-to-eight-week-old female BALB/c mice were purchased from the National Animal Center (Taipei, Taiwan). They were fed with a standard diet (LabDiet®; PMI Feeds, St. Louis, MO, USA) in the Animal House of National Defense Medical Center. The experiments were repeated at least twice. All protocols of animal experiments were approved by the Institutional Animal Care and Use Committee of National Defense Medical Center (approval number: IACUC-12-052).
Preparation of coffee and caffeine
Coffee was obtained from City Café of Uni-President Enterprises Corporation in Taiwan. The coffee product is ready-made Coffee arabica beans, medium roast, containing 0.625 milligram (mg) caffeine per milliliter (mL). The chemical caffeine (Sigma, St. Louis, MO, USA) was diluted to the same concentration. The mice were gavaged with coffee or caffeine at the same dosage of 0.3125 mg. Gavage of 0.5 mL liquid approximates the gastric volume of the mice.
Experiment protocols
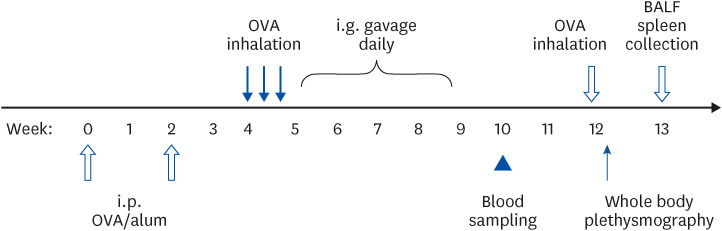
Groups of 6 BALB/c mice were sensitized intraperitoneally (i.p.) with 10 microgram (µg) OVA adsorbed onto 4 mg alum and suspended in 0.5 mL of phosphate-buffered saline (PBS) twice at a 2-week interval. Two weeks after the last sensitization, the mice were inhaled with 3% OVA aerosol (300 mg OVA in 10 mL PBS) for 15 min. OVA aerosol exposure was repeated 3 times within a week. One week after the first OVA exposure, the mice were treated with 0.5 mL coffee or caffeine both containing 0.3125 mg caffeine by using intragastric gavage once daily for 4 weeks. Control mice received ddH2O only. The mice were bled from the tail vein just a week following the final gavage. Mice were exposed to OVA aerosol again 3 weeks after the final gavage. Airway responsiveness detection was performed a day after the final OVA inhalation. Spleen sampling and bronchoalveolar lavage (BAL) collection were carried out a week later. These protocols are summarized in Fig. 1.
Fig. 1
Experimental protocol. BALB/c mice were i.p. sensitized twice with OVA in alum during a 2 weeks interval. Two weeks after the last sensitization, the mice were inhaled with OVA aerosol 3 times within 1 week. One week after the first OVA inhalation, the mice were i.g. gavage with coffee, caffeine or ddH2O. Blood were collected 1 week after the final of gavage. Mice were exposed to OVA aerosol again 3 weeks after the final gavage. Airway responsiveness were detected 1 day after the final OVA inhalation. Spleen and BAL lavage were collected 1 week later.
BAL, bronchoalveolar lavage; OVA, ovalbumin; i.p., intraperitoneally; i.g., intragastrically; BALF, bronchoalveolar lavage fluid.
Assessment of antibody response
Antibody levels were examined by enzyme-linked immunosorbent assays (ELISA) [22]. In brief, wells of plates (96 wells; Nunc, Kamstrup, Roskilde, Denmark) were coated with 100 μL OVA (100 μg/mL) in 0.05 M carbonate buffer, pH 9.6, stored overnight at 4°C. Sites not bound by OVA were blocked with 3% skimmed milk in PBS-Tween 20 for an hour. Serum samples (1/30–1/1,000) and standards (pooled hyperimmune sera after monthly sensitization with OVA emulsified in the complete Freund’s adjuvant for 4 months) in serial triple dilution were added in duplicates, following incubation for 5 h in room temperature. After washing, 100 μL of horseradish peroxidase-conjugated goat anti-mouse immunoglobulin (Ig) G (1/4,000; Jackson, West Grove, PA, USA), IgG2a or IgG1 (1/4,000; SBA, Birmingham, AL, USA) were added and the plates were incubated overnight at 4°C. After washing, color was developed by adding orthophenyleldiamine (0.5 mg/mL; Sigma) in citrate-carbonate buffer containing 0.015% hydrogen peroxide, incubated in the dark at room temperature. The color reaction was stopped by 4 N sulfuric acid and read at 492 nm using a reader (SPECTRAmax 250; Molecular Devices, Sunnyvale, CA, USA) and unknowns were interpolated according to the standard curve from serial diluted standards.
Passive cutaneous anaphylaxis
Serum-specific IgE antibody response was assessed in an in vivo animal model [22]. In sum, passive cutaneous anaphylaxis tests were examined in Sprague-Dawley mice. Aliquots of 100 µL of 2-fold dilutions of pooled mice serum samples (1/10–1/640) were intradermally injected. The mice were then challenged 24 h later by an intravenous injection of 2 mg OVA and 5 mg Evans Blue in 1 mL of PBS. The reaction was read 30 min later. The PCA titer was taken as the highest dilution giving a positive reaction of at least 5 mm in diameter and the results were expressed as mean ± SD.
Airway responsiveness
Airway hyperresponsiveness is the hallmark of bronchial asthma [23]. Airway hyperresponsiveness was detected using whole body plethysmography (Buxco, Troy, NY, USA) in unrestrained and conscious mice [24]. Mice were acclimatized in the main chamber for 10 min. After measuring baseline values for 5 min, the mice were subsequently challenged with aerosolized PBS or increasing concentration of methylcholine (6.25, 12.5 and 25 mg/mL; Sigma) for 3 min. The response in terms of enhanced pause (Penh) was recorded for 6 min after each dose. Penh reflects airway resistance and is used to monitor pulmonary dysfunction. Increased Penh was used as an index of in vivo airway obstruction.
Bronchoalveolar lavage fluid (BALF)
Mice were sacrificed by i.p. injection of 20 mg pentobarbitone a week after the final aerosol challenge. The tracheas were cannulated then lavaged twice with 1 mL of PBS. The recovered fluid, BALF, was centrifuged at 600 ×g for 10 min at 4°C. The supernatant was stored at −70°C until examination.
Cytokine assays
One week after the final aerosol challenge, spleen cells from 6 BALB/c mice in each group were sampled and cultured (4 × 106 cells/well) with OVA in duplicate in complete RPMI-1640 medium (1 mg/mL), 10% fetal calf serum and antibiotics in 24-well flat-bottomed microtitre plates (1 mL/well; Costar, Cambridge, MA, USA). The supernatants were harvested after 1–3 days of culture. The levels of cytokines interleukin (IL)-4, IL-10 and interferon-γ (IFN-γ) from the harvested supernatants and rewarmed BALF were measured using appropriate sandwich ELISA kits (eBioscience, San Diego, CA, USA).
Statistics
All experiments were performed in duplicates. Antibody, cytokine titers and Penh were expressed as means ± SD. Data from the 3 groups were analyzed using the Shapiro-Wilk test to determine normal distribution. Normally distributed data are further analyzed using the one-way analysis of variance and post-hoc Scheffé test. Non-normal distributions are analyzed with the non-parametric Mann-Whitney U test.
RESULTS
Allergen-specific antibody response after intragastric gavage
The immunomodulatory effects of coffee on OVA-specific antibody response were studied. Antibody concentration was recorded, without significant differences prior to or after sensitization (data not shown). Systemic antibody responses were examined a week after treatment (Fig. 1).
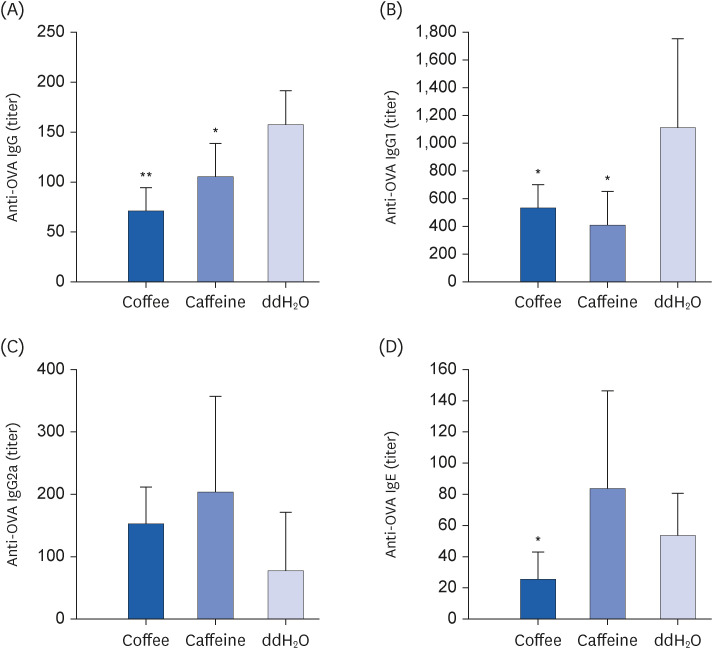
When compared with the control, coffee-gavaged mice had significantly reduced OVA-specific IgG and IgG1 antibody responses (P < 0.05 respectively; Fig. 2A and B). Furthermore, coffee intake effectively attenuated allergen-specific IgE antibody in comparison with the controls (P < 0.05 respectively; Fig. 2D). Interestingly, although caffeine gavage effectively reduced OVA-specific IgG and IgG1 antibody concentrations (P < 0.05 respectively; Fig. 2A and B), IgE antibody was not attenuated in comparison with the control (Fig. 2D). There were no significant differences in specific IgG2a antibody levels between both groups (Fig. 2C).
Fig. 2
Effect of coffee or caffeine intake on specific antibody responses. Groups of 6 BALB/c mice were sensitized with and treated as described in Fig. 1. Serum specific (A) IgG, (B) IgG1, and (C) IgG2a antibody responses (titers) 1 week after the final gavage were assessed by ELISA, and (D) IgE by in vivo passive cutaneous anaphylaxis test. Results are shown as mean ± SD. Significant differences were analyzed by one-way analysis of variance and post-hoc Scheffé test or Mann-Whitney U test.
Ig, immunoglobulin; ELISA, enzyme-linked immunosorbent assay.
*P < 0.05 and **P < 0.005 compared to ddH2O group.
Cytokine profiles of spleen cells
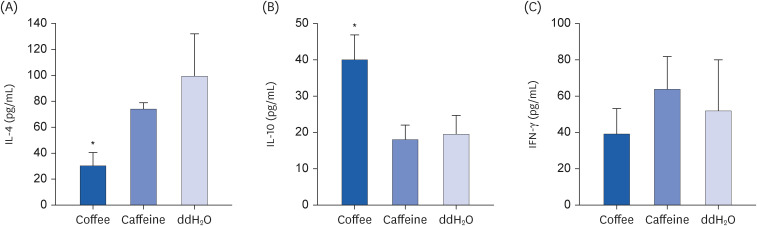
Cytokine profiles after coffee were further studied (Fig. 3). Compared to the control, mice after coffee had significantly reduced IL-4 but enhanced IL-10 concentrations from cultured spleen cells (P < 0.05, respectively; Fig. 3A and B). There was no significant difference in IFN-γ concentration in spleen cells between coffee-treated mice and the controls (Fig. 3C). On the other hand, spleen cells from mice receiving caffeine had mildly attenuated IL-4 concentration in comparison with the controls; however, it had no statistical difference (P > 0.05; Fig. 3A). IL-10 and IFN-γ concentrations from cultured spleen cells were also similar between mice after caffeine and those after ddH2O (P > 0.05, respectively; Fig. 3B and C).
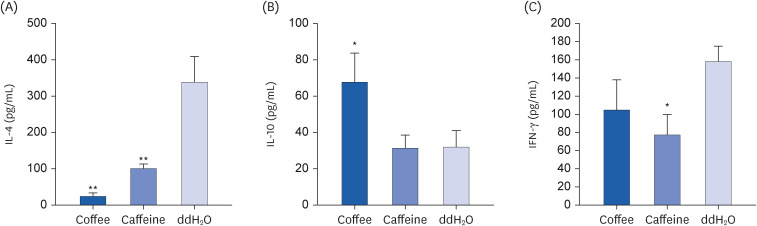
Fig. 3
Effect of coffee or caffeine treatment on cytokine levels in cultured spleen cells. Groups of 6 BALB/c mice were treated as described in Fig. 1. One week after the final aerosol challenge, their spleen cells were collected and cultured for 1 to 3 days. The cytokine levels in the supernatant were measured in duplicate by sandwich ELISA kits. (A) IL-4, (B) IL-10, and (C) IFN-γ. Results are shown as mean ± SD. Significant differences between groups were determined by one-way analysis of variance and post-hoc Scheffé test.
ELISA, enzyme-linked immunosorbent assay; IL, interleukin; IFN, interferon.
*P < 0.05 compared to ddH2O group.
Cytokine profiles in BALF
Cytokine profiles from BALF were collected and examined. Similar to the findings from spleen cells, mice after coffee presented with significantly reduced IL-4 and increased IL-10 concentrations in BALF than the controls (P < 0.05, respectively; Fig. 4A and B). On the other hand, compared with the controls, caffeine-gavaged mice had significantly decreased IL-4 concentration (P < 0.05; Fig. 4A). Nevertheless, BAL IL-10 concentration after caffeine consumption showed no significant difference from the controls (Fig. 4B).
Fig. 4
Effect of coffee or caffeine gavage on cytokine levels of BALF. Groups of 6 BALB/c mice have the same treatment as described in Fig. 3. BALF was collected 1 week after the final aerosol challenge. The cytokine levels in BALF were examined in duplicate by sandwich ELISA kits. Refer to Fig. 1 for details of the experimental protocols. (A) IL-4, (B) IL-10, and (C) IFN-γ. Results are shown as mean ± SD. Significant differences between groups were determined by one-way analysis of variance and post-hoc Scheffé test.
BALF, bronchoalveolar lavage fluid; ELISA, enzyme-linked immunosorbent assay; IL, interleukin; IFN, interferon.
*P < 0.05 and **P < 0.005 compared to ddH2O group.
Airway responsiveness determination
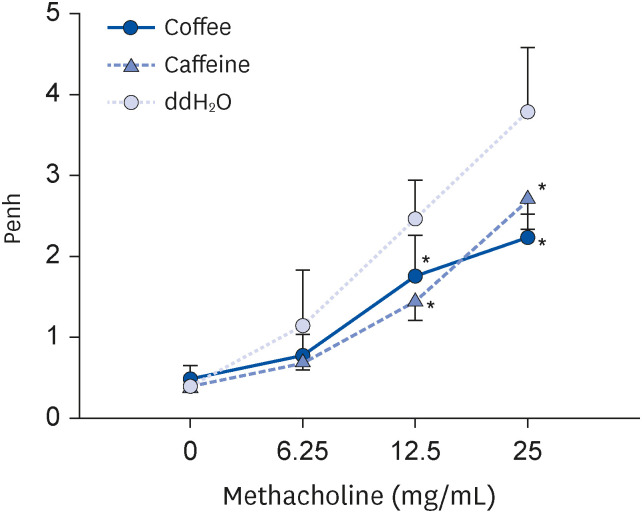
Airway responsiveness after coffee was further examined (Fig. 2). Mice receiving coffee and caffeine both had significantly decreased Penh values in comparison with the control (P < 0.05 respectively; Fig. 5). As shown, airway hyperresponsiveness in mice was effectively reduced after coffee and caffeine.
Fig. 5
Effect of coffee or caffeine therapy on airway hyperresponsiveness. Groups of 6 BALB/c mice were sensitized and treated as described in Fig. 1. One day after the final aerosol challenge, airway responsiveness were measured 1 day after the final aerosolized OVA challenge using a whole-body plesthysmograph. Results are shown as mean ± SD. The concentration of methacholine denotes the concentration required to reach airway hyperresponsiveness. Significant differences between groups were determined by one-way analysis of variance and post-hoc Scheffé test.
OVA, ovalbumin; Penh, enhanced pause.
*P < 0.05 compared to ddH2O group.
DISCUSSION
In this study, we present the therapeutic potential of coffee on asthma and allergic reactions. Coffee-treated mice demonstrated immune profiles which inhibited systemic T helper 2 (Th2) response. Our results demonstrate that airway hyperresponsiveness of OVA-sensitized mice was effectively improved after treatment with coffee or caffeine, which accepts our hypothesis that coffee effectively reduced airway hypersensitivity. This is the first study to demonstrate that coffee intake effectively suppressed Th2 immune reaction and reduced airway hyperresponsiveness in allergen-sensitized mice.
Production of cytokines by T cells after allergen challenge has important regulatory consequences on the inflammatory response in the lung [23]. The pathogenesis of asthma is the result of an augmented IL-4-driven Th2 response, culminating in eosinophil inflammation and airway hyperresponsiveness [23]. IL-4 is a pivotal cytokine in the production of IgE antibody, which also regulates airway eosinophilia [25]. In our study, IL-4 production in spleen cells and serum allergen-specific IgE antibody responses were significantly reduced in the coffee-treated group, demonstrating the anti-inflammatory effect of coffee. In addition, both coffee and caffeine directly affected the respiratory system by expressing significantly decreased IL-4 concentration in BALF compared with the control (P < 0.005). Reduced IL-4 and IFN-γ concentrations as demonstrated by the coffee-treated murine model in our study could potentially reduce airway eosinophilia, leading to improved airway resistance. Additionally, enhanced IL-10 concentration expressed by the coffee group further suppressed the Th2 dominant allergic reaction. To compare, while the immune profile of the caffeine group in BALF displayed significant anti-inflammatory results, immune profile of the spleen cells were only slightly attenuated without statistical significance. Moreover, suppression of Th2 allergic reaction was not emphasized in the caffeine group. While caffeine indeed posed anti-inflammatory properties, the immunomodulatory effect was not as systemic and profound when compared to coffee. Other than caffeine, certain chemical compounds in coffee play an important role in increasing IL-10 while suppressing IL-4 concentration, which enhances the drive towards reducing Th2 allergic reaction. Besides caffeine, which is a methylxanthine that induces bronchodilatation and improves lung function [20], other major bioactive compounds of coffee include phenols (chlorogenic acids and derivatives), trigonelline, nicotinic acid (vitamin B3), diterpenes (cafestol, kahweol), and short-chain carboxylic acids. Of these, chlorogenic acids and derivatives act as antioxidants and showed antiviral activities, which potentially possess anti-inflammatory properties [1]. Further studies are encouraged to investigate potential compounds contained in coffee for allergy and asthma control.
Goto et al. [26] investigated the effects of orally administered coffee on allergic responses using a T cell receptor-transgenic DO11.10 allergic mouse model. Splenocytes from coffee-administered mice showed decreased IL-2 but increased IL-12p40 concentration. The serum antigen specific IgE concentration was also reduced. Similar to our study, they also demonstrated that coffee could prevent allergic reaction.
Coffee may potentially suppress local pulmonary inflammation induced by OVA aerosol. The cytokine profile after long term coffee intake requires further evaluation.
Asthma is characterized by airway hyperresponsiveness, chronic airway inflammation and airflow limitation which result in the clinical expression of airway obstruction [27]. Airway hyperresponsiveness could be documented by decreased bronchial airflow after bronchoprovocation with methacholine or histamine [28].
Caffeine had been shown to improve pulmonary function and to prevent exercise-induced bronchoconstriction in patients with bronchial asthma [29]. It should be withdrawn from the diet for more than 4 h before performing pulmonary function test, as caffeine could lead to misinterpretation of the results. In our study, we confirmed that both coffee and caffeine can attenuate airway hyperresponsiveness, which may be attributed to their bronchodilator effect.
Caffeine not only can reduce airway hyperresponsiveness but also contribute to modulating immunological responses. It suppresses cytokine production, antibody production, chemotaxis, leukocyte proliferation and increases myeloperoxidase production [30]. Caffeine also inhibits antigen-stimulated IL-4 and IL-2 concentrations in mice [31]. Similar to these findings, we demonstrate that IFN-γ and IL-4 concentrations in BALF were suppressed by caffeine intake. These may be the reasons for reduced IL-4 concentration without concomitant elevation of IL-10 in caffeine-treated mice.
The limitations of this study include the lack of histologic assessment to confirm and compare the pathogenesis and severity of airway hyperresponsiveness before and post exposure to coffee.
Coffee intake effectively attenuated OVA induced systemic allergic responses and airway hyperresponsiveness. Besides caffeine, other constituents in coffee may contribute to the immunomodulating effects. Further studies are necessary to analyze the constitutions in coffee, as well as the dosage effect and elucidate their impacts on the immune reaction.
 XML Download
XML Download