

 PDF
PDF Citation
Citation Print
Print


Introduction
Injury and aging-related disorders of the cartilage often result in the development of degenerative joint diseases, such as osteoarthritis (OA) (1). OA is characterized by the progressive destruction of articular cartilage, suggesting that chondrocytes play key roles in the pathogenesis of OA. Although chondrocytes have low mitotic activity, they are responsible for the synthesis and turnover of the articular cartilage matrix. Thus, with the onset of OA, chondrocytes undergo multiple changes in their proliferation, viability, and secretory profile (2). As most drugs for OA aim to relieve pain and inflammation, thereis a lack of pharmaceutical therapies that can regenerate cartilage by stimulating the chondrogenesis of endogenous stem cells and increasing growth of the cartilaginous matrix of chondrocytes.
Recent studies have shown that a population of cartilage stem/progenitor cells similar to populations of mesenchymal stem cells (MSCs) is involved in cartilage repair and maintenance of chondrocyte homeostasis (3). These cartilage stem/progenitor cells have been observed in human, equine, and bovine articular cartilages. Phenotypic analyses have shown that cartilage stem/progenitor cells express various MSC-related surface markers, including cluster of differentiation (CD)-29 (integrin β-1), CD44, and CD105 (3). Therefore, a better understanding of the mechanisms involved in the differentiation of MSCs into chondrocytes may enable the screening of small molecules to identify those that promote the chondrogenic differentiation of cartilage stem/progenitor cells and the repair of abnormal cartilage in OA.
Wedelolactone, which is thought to be the active component of the dried aerial part of Eclipta prostrata, exhibits anti-inflammatory activities, including the inhibition of IkappaB (IκB) kinase (4). In a previous study, we showed that wedelolactone promoted the serine/threonine phosphorylation of nucleotide-binding and oligomerization domain-like receptor family pyrin domain-containing 3, which is dependent on protein kinase A signaling, to block inflammasome activation (5). Moreover, recent studies have revealed the effects of wedelolactone on MSC differen-tiation. For example, wedelolactone was reported to inhibit the adipogenic differentiation of human adipose tissue-derived MSCs (6). In addition, Liu et al. (7, 8) have demonstrated that wedelolactone enhances the osteoblastogenesis of MSCs but inhibits their osteoclastogenesis (9). Another study showed that wedelolactone stimulates odontoblast differentiation (10). However, the effects of wedelolactone on the chondrogenic differentiation of MSCs remain largely unknown.
The pharmacological activity of wedelolactone was previously attributed to its inhibition of IκB kinase, caspase-11, and/or androgen receptors (11-13). However, there is emerging evidence that wedelolactone can act indepen-dently of these factors. For example, the anti-tumor effects of wedelolactone may be due to its inhibition of DNA topoisomerase IIα (14). Wedelolactone was also found to have potent G protein-coupled receptor-35 agonist activity (15). Furthermore, recent studies have demonstrated that wedelolactone can target the enhancer of zeste homolog 2 (EZH2)-embryonic ectoderm development complex to promote H3K27 demethylation, indicating that wedelolactone has potential to be used as an epigenetic agent in cancer treatment (16). Thus, the cellular targets of wedelolactone are context-dependent, and the exact mechanisms by which wedelolactone modulates MSC differentiation remain to be identified.
In the study described in this paper, we investigated the effect of wedelolactone on the chondrogenic differentiation of human induced pluripotent stem cell (iPSC)-derived MSCs and rat bone-marrow MSCs (BMSCs). Intriguingly, we found that compared with MSCs exposed to control (wedelolactone-lacking) media, MSCs exposed to wedelolactone-containing media exhibit significantly increased chondrocyte differentiation. We also found that forkhead box O 1 (FOXO1), a transcription factor that plays an important role in stem cell differentiation, is stimulated by wedelolactone during the chondrogenic differentiation of MSCs. Accordingly, we investigated the mechanism by which wedelolactone induces the FOXO1 pathway in the chondrogenic differentiation of MSCs.
Materials and Methods
Cell culture
Human iPSCs (hiPSCs) were obtained according to a reported method by reprogramming somatic cells derived from a patient with OA (17). The hiPSCs were maintained on Matrigel (Corning, USA)-coated dishes in PeproGrow hESC medium (PeproTech, USA) containing 2 μM Y-27632. The medium was changed every day. All cells were cultured at 37℃ in a humidified atmosphere of 5% CO2 and 95% air.
The hiPSCs were then induced to form embryoid bodies (EBs). Briefly, iPSCs were digested and resuspended in EB formation medium and then transferred to an ultra-low adhesion six-well plate (Corning, USA). The differentiation medium was changed every 2 days. After 10 days, the EBs were transferred into 0.1% gelatin-coated culture plates (Corning, USA) and cultured for a further 3 days. After this time, the cells were digested and then passed through a 100-μm nylon mesh. The resulting filtered cells were resuspended in an MSC growth medium comprising Dulbecco’s Modified Eagle Medium (DMEM Gibco, USA) supplemented with 10% fetal bovine serum (FBS; Gibco, USA), MEM Non-Essential Amino Acids (Gibco, USA), 2 ng/ml FGF (PeproTech, USA), and 20 ng/ml epidermal growth factor (PeproTech, USA).
Collection and culture of BMSCs
The tibia and femur of Sprague–Dawley (SD) rats were isolated under sterile conditions and then immersed in cooled phosphate-buffered saline (PBS) and washed to remove attached blood. The bone marrow cavity was washed three times with DMEM containing 10% FBS. The bone marrow cells were cultured at 37℃ in a 5% CO2 atmosphere. After 24 h, the nonadherent cells were gently washed off with PBS, and the MSC medium was added.
Flow cytometric analysis
The hiPSC-derived MSCs or rat BMSCs were harvested using 0.25% trypsin/ethylenediaminetetraacetic acid and resuspended in 100 μl staining medium (2% FBS and 2% N-2-hydroxyethylpiperazine-N’-2-ethanesulfonic acid in PBS). The cells were stained with mouse anti-human CD29 (BD, USA), mouse anti-human CD44 (BD, USA), mouse anti-human CD34 (BD, USA), mouse anti-human CD105 (BD, USA), and mouse anti-human CD45 (BD, USA) antibodies for 30 min at 4℃. Isotype-matched antibody (immuno-globulin G 2b-fluorescein isothiocyanate) was used to determine nonspecific fluorescence. Samples were run on a cytometer (CytoFLEX Beckman Coulter, USA).
Chondrogenic differentiation of MSCs
MSCs were chondrogenically differentiated (18, 19) by first centrifuging 5×105 hiPSC-derived MSCs or rat BMSCs cells at 500×g in 15-ml centrifuge tubes for 10 min to form small pellets. The cell pellets were cultured for 21 days in a chondrogenic differentiation medium comprising DMEM (Gibco, USA) supplemented with 10% FBS, 1:100 Insulin–Transferrin–Selenium (ThemoFisher, USA), 100 nM dexamethasone, 10 ng/ml recombinant transforming growth factor (TGF)-β3 (PeproTech, USA), and 40 μg/ml L-proline (Sigma-Aldrich, USA). The medium was changed every 3 days.
Cartilage defect rat model
We established cartilage defect rat models as previously described (20). Eight-week-old SD rats were purchased from the Experimental Animal Center of Guangzhou University of Chinese Medicine and housed in individually ventilated microisolator cages under a 12-h dark/light cycle. The rats were anesthetized by intraperitoneal injection of ketamine. The skin around the knee was disinfected, and the right knee was exposed for surgery. Next, a defect (1.5 mm in diameter and 1.5 mm in depth) was created in the center of the groove. The rats were divided into two groups: the experimental group, which was implanted with hydrogel mixed with rat BMSCs pretreated with wedelolactone, and a control group, which was implanted with hydrogel mixed with non-pretreated rat BMSCs. After 6 weeks, all rats were sacrificed, and their joint tissues were collected for further analysis.
Transfection of human induced pluripotent stem cell-derived MSCs with short hairpin RNAs (shRNAs)
FOXO1 and EZH2 shRNA sequences (listed in Supplementary Table S1) were designed and cloned into the pLKO-puro lentiviral vector. The constructed shRNA-expressing plasmid was co-transfected with packaging plasmids pVSVg and psPAX2 into human embryonic kidney 293T cells using Lipofectamine 3000 to generate lentiviral particles. The iMSCs were infected with the shRNA lentiviruses to knock down the expression of FOXO1 or EZH2.
Safranin O and Fast Green staining
Five-micrometer slices were made of formalin-fixed paraffin-embedded joint tissues for safranin O and Fast Green staining (Solarbio, China), according to the manufacturer’s instructions. Briefly, the slides were deparaffinized and stained with Weigert’s hematoxylin working solution for 3∼5 min and then washed with water and treated with acid differentiation solution for 15 s. The slides were then rinsed in water for 10 min and subsequently stained with Fast Green staining solution for 5 min. After this time, the slides were rinsed in weak acid solution for 10∼15 s to remove the remaining Fast Green staining solution and then stained with safranin O staining solution for 5 min.
Alcian blue staining and toluidine blue staining of chondrocytes
MSCs were cultured in the chondrogenic differentiation medium for 3 days. Subsequently, the medium was removed, and the cells were washed three times with PBS and then stained with toluidine blue staining solution (Solarbio, China) or Alcian blue staining solution (Solarbio, China) for 30 min. Next, the cells were washed with PBS for 5 min, and the washed stained cells were then observed under a microscope.
Histological staining of chondrogenic pellets
Chondrogenic pellets were collected after 21 days of chondrogenic differentiation. The pellets were fixed in 4% buffered formaldehyde for 6 h and then dehydrated in alcohol. The pellets were subsequently clarified in xylene three times and the pellets were immersed in paraffin for 1 h in a 65℃ oven. The paraffin-embedded pellets were sliced to a thickness of 7 μm for histological staining. The slides were deparaffinized by successive treatment with 200 ml of 100%, 95%, 80%, and 75% ethanol for 4 min each. The slides were then incubated in 3% hydrogen peroxide for 15 min to block endogenous peroxidases. Next, the slides were blocked with goat serum solution at room temperature for 10∼15 min and then incubated with 100 μl of primary antibodies—anti-collagen type II alpha-1 (COL2A1 Abcam, ab34712; 1:500) and anti-aggrecan (ACAN Proteintech, 13880-1-AP; 1:500)—at 4℃ overnight. The slides were subsequently washed three times with PBS and then incubated with an appropriate amount of biotin-labeled goat anti-mouse/rabbit immunoglobulin G (IgG) polymer at room temperature for 15 min. After this time, the slides were washed three times with PBS and then incubated with an appropriate amount of horseradish enzyme-labeled streptomyces ovalbumin working solution (Cell Signaling Technology, USA) for 10∼15 min. Finally, the slides were washed three times with PBS, incubated with 3,3’-diaminobenzidine (Cell Signaling Technology, USA) solution for 5∼8 min, and then observed under a microscope.
Immunofluorescence
MSCs were cultured in the chondrogenic differentiation medium in confocal dishes at 37℃ in a 5% CO2 atmosphere for 3 days. The cells were then fixed with 4% paraformaldehyde (dissolved in PBS, pH 7.4) for 10 min at room temperature. Subsequently, the cells were incubated with PBS (containing 0.1%∼0.25% Triton X-100) for 10 min and then incubated with goat serum at room temperature for 30 min to block the nonspecific binding of antibodies. Thereafter, the cells were incubated with 100 μl of primary antibodies—anti-COL2A1 (Abcam, ab34712; 1:200), anti-ACAN (Proteintech, 13880-1-AP; 1:200), and anti-SXO9 (Abcam, ab185966; 1:200)—at 4℃ overnight. On the second day, the cells were washed three times with PBS and then incubated with secondary antibodies for 1 h at room temperature. 4’,6-Diamidino-2-phenylindole (The-rmoFisher, USA) was used to stain the nuclei, and the stained cells were observed under a confocal microscope.
Real-time reverse transcription–polymerase chain reaction (qRT-PCR) analysis
The total RNA of the cells was extracted used TRIzol reagent (Life Technologies, USA) and then reverse-transcribed into cDNA using HiScript II RT SuperMix for qPCR (Vazyme, China). The final qPCR mix contained 5 μl of 2×ChamQ Universal SYBR qPCR Master Mix (Vazyme, China). The relative expression levels of mRNAs were detected and analyzed using QuantStudio 6 Flex Real-Time PCR Systems (Applied Biosystems, USA). The primer sequences used for qRT-PCR analysis are listed in Supplementary Table S2. The relative mRNA expression level was calculated using the 2-ΔΔCt method.
Western blotting
Cells were washed three times with PBS and then lysed in radio-immunoprecipitation assay lysis buffer (Ther-moFisher, USA) containing phenylmethanesulfonyl fluoride (Beyotime, China) and protease inhibitor cocktail (Millipore, USA) at 4℃ for 30 min. The concentrations of protein were assayed using the PierceTM BCA Protein Assay Kit (ThermoFisher, USA). The proteins were then separated by 10% sodium dodecyl–sulfate polyacrylamide gel electrophoresis and transferred to a polyvinylidene difluoride (PVDF) membrane (Millipore, USA). The PVDF membrane was blocked with 5% skim milk in 1×Tris-buffered saline with 0.1% Tween-20 (TBST) for 1 h and then incubated overnight at 4℃ with primary antibodies: anti-SRY-box transcription factor 9 (SOX9 Abcam, ab185966; 1:1,000), anti-COL2A1 (Abcam, ab34712; 1:500), anti-FOXO1 (Cell Signaling Technology, #2880; 1:1,000), anti-EZH2 (Cell Signaling Technology, #5246; 1:1,000), anti-histone 3 lysine 27 trimethylation (H3K27me3) (Cell Signaling Technology, #9733; 1:1,000), anti-Histone H3 (Cell Signaling Technology, #60932; 1:1,000), anti-EZH2 (Cell Signaling Technology, #5246; 1:1,000), and anti-β-ACTIN (Proteintech, #60008-1-Ig; 1:1,000). The PVDF membrane was washing three times with TBST (10 min each time) and then incubated with horseradish peroxidase (HRP)-linked anti-rabbit IgG (Cell Signaling Technology, #7074; 1:2,000) and HRP-linked anti-mouse IgG (Cell Signaling Technology, #7076; 1:2,000) for 1 h at 37℃. The expression levels of proteins were detected by chemiluminescence (Millipore, USA).
Dual-luciferase reporter assay
The wild-type 3’-untranslated region (UTR; FOXO1-3’-UTR-wt) or a 3’-UTR containing a mutant (FOXO1-3’-UTR-mut) sequence of the miR-1271-5P-binding site was cloned into the psiCheck2 plasmid (Promega). The iMSCs were seeded in a six-well plate and transfected with the reporter plasmid. After 24 h, the cells were infected with a miR-1271-5P mimic or miR-1271-5P inhibitor. The Dual-GloⓇ Luciferase Assay System (Promega Madison, WI) was used to measure reporter activity at 72 h according to the manufacturer’s protocol. Renilla luciferase was used as the internal control to normalize the results.
Chromatin immunoprecipitation (ChIP)–qPCR
The cells were seeded in a 10-cm dish and cross-linked with 1% paraformaldehyde for 10 min, followed by incubation with glycine to terminate the cross-linking. The cells were then lysed on ice and centrifuged at 12,000×g for 10 min. The nuclei of the cells were resuspended in the lysis solution, the chromatin DNA of the cells was sonicated, and 30 μl of protein A-agarose beads were added to the mixture. The resulting mixture was incubated with anti-H3K27me3 antibody, anti-EZH2 antibody, or control IgG for 2 h at 4℃. Subsequently, the antibody–protein–DNA complexs were eluted from the beads. The remaining proteins and RNAs were degraded with protease K and RNase A. The DNA was extracted with phenol–chloroform and purified using a PCR purification kit.
Isobaric tags for relative and absolute quantitation (iTRAQ) and tandem mass tags
Chondrogenic pellets were lysed by heating with an appropriate amount of lysate solution at 95℃ for 10 min. The mixture was centrifuged at 12,000×g for 10 min, and the supernatant was transferred to a new 1.5-ml tube and then treated with dithiothreitol. The resulting protein solution was transferred to a 10 K ultrafiltration tube for centrifugation. The lysate solution was then discarded and replenished. Trypsin was added to the ultrafiltration tube, and the proteins were enzymatically hydrolyzed at 37℃ for 16 h. The enzymatically hydrolyzed polypeptides were concentrated and then labeled with the iTRAQ reagent. The iTRAQ-labeled samples were analyzed by liquid chromatography–tandem mass spectrometry (Thermo Fisher Easy-nlC 1000 Liquid Chromatograph and Thermo Fisher Q Exactive).
Bulk RNA sequencing and analysis
Total RNA was extracted from chondrogenic pellets using TRIzol reagent (Life Technologies, USA). The integrity of the RNA was tested using the Agilent Bioanalyzer 2100. Paired-end reads were generated using the IlluminaⓇ Hi-Seq 2500 platform and mapped to the human genome. DESeq was applied for transcription quantification and differential expression analysis using a cutoff of p<0.05.
Statistical analysis
Bioinformatic analyses—Kyoto Encyclopedia of Genes and Genomes (KEGG) and Gene Set Enrichment Analysis (GSEA)—were performed using the OmicStudio tools at https://www.omicstudio.cn/tool. All statistical analyses of the experimental data were performed using SPSS 10.0. Data are presented as means±standard deviations of three independent experiments. Comparisons between three groups were performed using a one-way analysis of variance, while those between two groups were performed using a two-sided Student’s t-test. For all tests, statistical significance was set at p<0.05.
Results
Wedelolactone promotes the differentiation of hiPSC-derived MSCs to chondrocytes in vitro
Using a modified stepwise protocol, we derived human MSCs from an iPSC line established from a patient with OA (Fig. 1A; Supplementary Fig. S1A). A homogeneous culture of hiPSC-derived MSCs had a fibroblastic spindle-shaped morphology that resembled BMSCs and expressed CD29, CD44, and CD105 but not CD45 or CD34 (Supplementary Fig. S1B). To evaluate the effects of wedelolactone on the chondrogenesis of MSCs, we cultured hiPSC-derived MSCs with and without wedelolactone during chondrogenic differentiation by following a previously reported protocol (18) (Fig. 1A). Chondrogenic pellets were obtained after 21 days of chondrogenic differentiation (Fig. 1B). We observed upregulated levels of gene expression of SOX9, COL2A1, and ACAN in all hiPSC-derived MSC chondrogenic pellets after 21 days of chondrogenic differentiation (Fig. 1C). Moreover, the expression levels of these cartilage-marker genes were increased further after wedelolactone treatment compared with no treatment (Fig. 1C). Immunohistochemical results also showed a significant upregulation of COL2A1 and ACAN after wedelolactone treatment (Fig. 1D). We also observed increased levels of expression of the proteins COL2A1, ACAN, and SOX9 after wedelolactone treatment, compared with no treatment, under two-dimensional culture conditions (Fig. 1E, Supplementary Fig. S1C and S1D). These data indicate that wedelolactone significantly promotes the chondrogenic differentiation of hiPSC-derived MSCs.
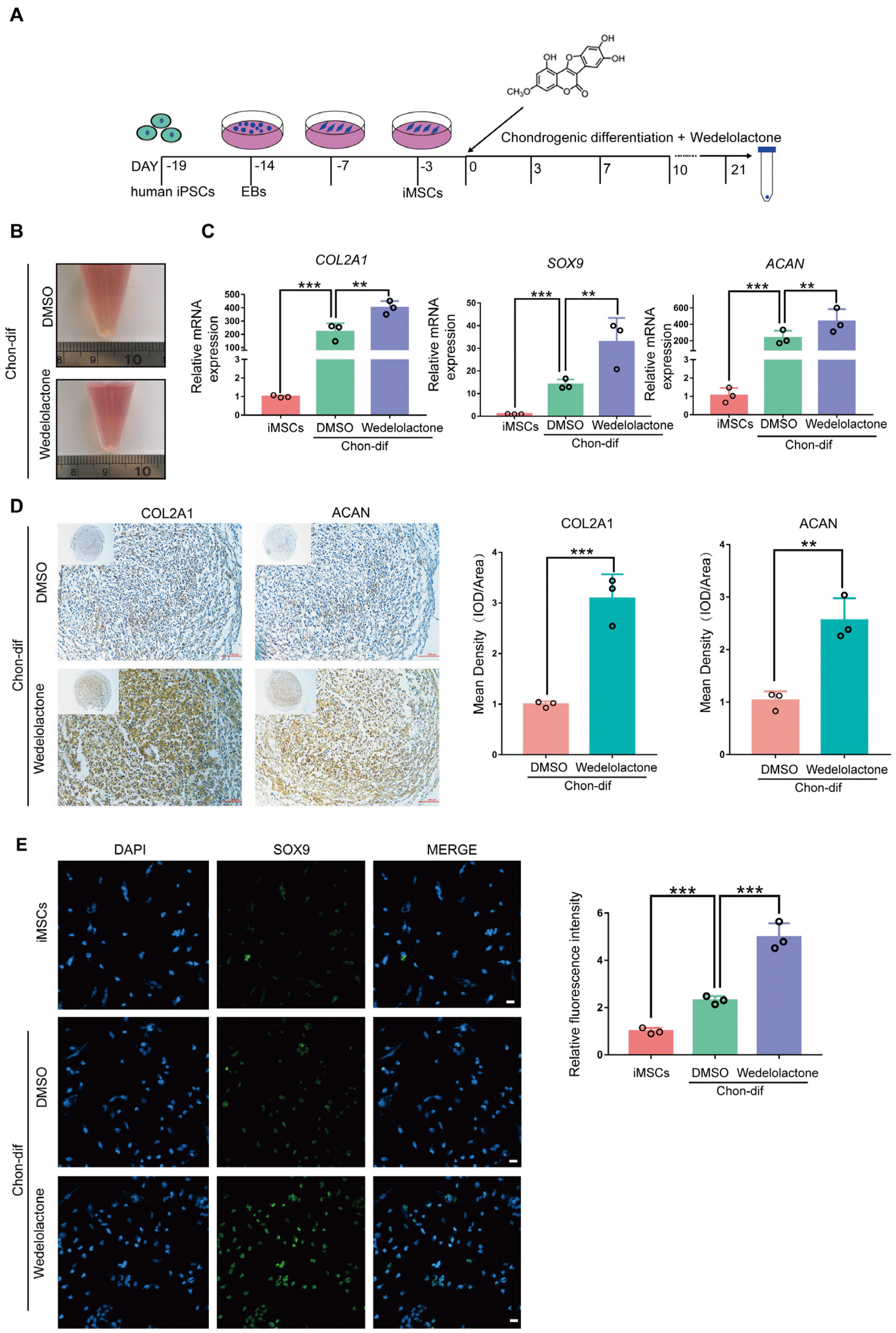
Fig. 1
Wedelolactone promotes the differentiation of hiPSC-derived MSCs to chondrocytes in vitro. (A) Schematic procedure of the chondrogenic differentiation of MSCs induced by human iPSC-derived MSCs. (B) Image of the chondrogenic pellet differentiated from human iPSC-derived MSCs. (C) Gene expression analysis of the chondrogenic differentiation markers (collagen type II alpha-1 [COL2A1], SRY-box transcription factor 9 [SOX9], and aggrecan [ACAN]) in the chondrogenic pellet. (D) Immunohistochemistry image of the chondrogenic pellet stained with COL2A1 and ACAN. Scale bar=100 μm. Mean density was used to quantify the COL2A1 and ACAN contents in the chondrogenic pellet. (E) Immunofluorescence image of iPSC-derived MSCs under chondrogenic differentiation stained with SOX9, COL2A1, and ACAN. Scale bar=100 μm. Relative fluorescence intensity was used to quantify the expression levels of SOX9, COL2A1, and ACAN. Data are expressed as the mean±standard deviation (SD) (n=3). Statistical differences were analyzed by one-way analysis of variance (ANOVA) followed by Dunnett’s test: **p<0.01, ***p<0.001. EB: embryoid body, iMSCs: human iPSC-derived MSCs, Chon-dif: chondrogenic differentiation.
![]()
Wedelolactone promotes the chondrogenic differentiation of rat BMSCs
To further confirm the role of wedelolactone in promoting chondrogenic differentiation, we used another differentiation system, as previously reported (21). Rat BMSCs were induced to differentiate into chondro-cytes using the chondrogenic differentiation medium under three-dimensional culture conditions, with and without wedelolactone (Fig. 2A). Chondrogenic pellets were formed after 21 days of chondrogenic differentiation (Fig. 2B). We observed significantly increased levels of expression of chondrogenic-marker genes (COL2A1, ACAN, and SOX9) after wedelolactone treatment compared with no treatment (Fig. 2C). Wedelo-lactone also upregulated the protein levels of these chondrogenic markers during the chondrogenic differentiation of rat BMSCs (Fig. 2D and 2E).
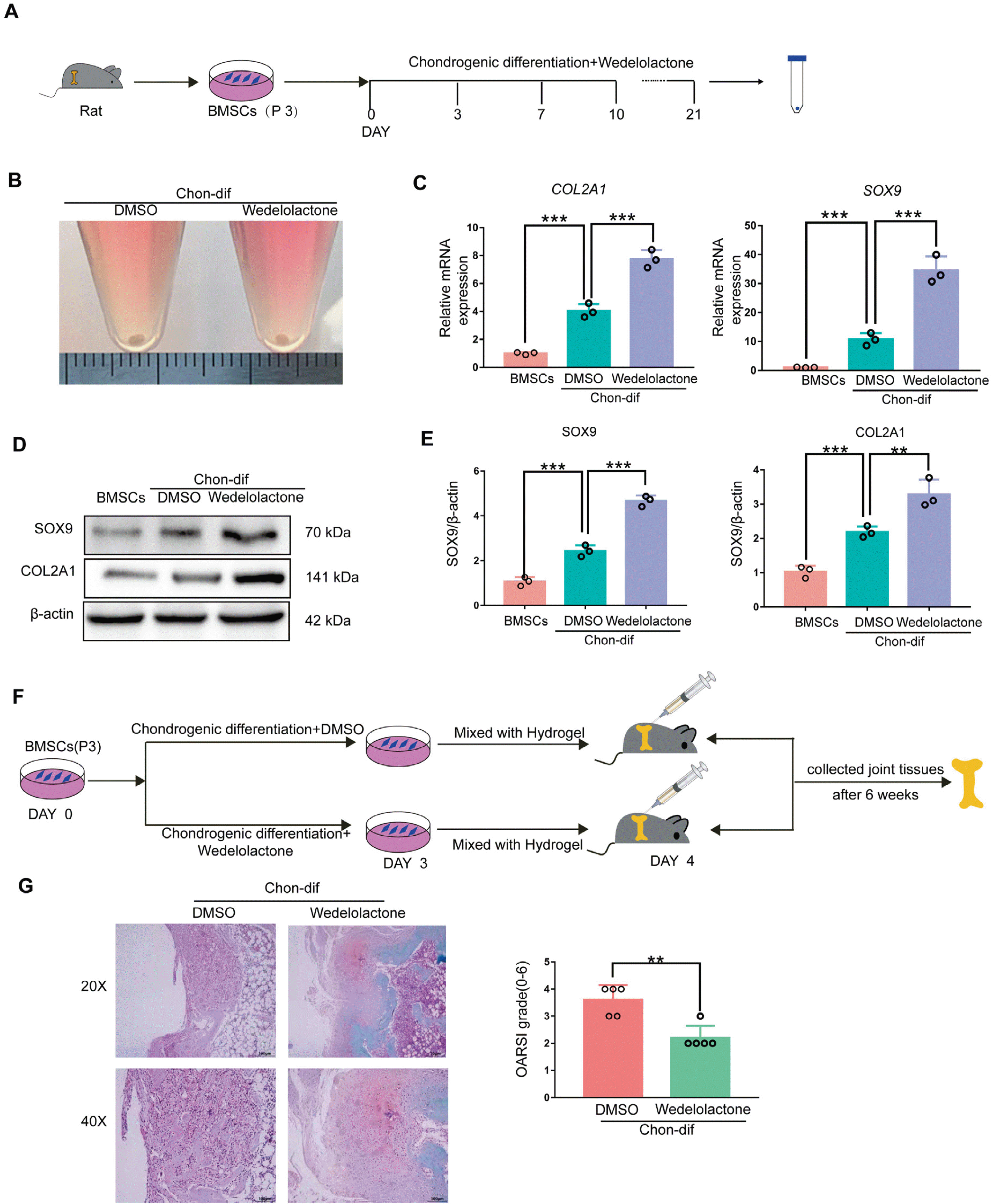
Fig. 2
Wedelolactone promotes the chondrogenic differentiation of rat BMSCs. (A) Schematic procedure of the chondrogenic differen-tiation of BMSCs. (B) Image of the chondrogenic pellet differentiated from BMSCs. (C) Gene expression analysis of the chondrogenic differentiation markers (COL2A1, SOX9, and ACAN) in the chondrogenic pellet. (D) Western blotting analysis of the chondrogenic differentiation markers (COL2A1 and SOX9) in the chondrogenic pellet. (E) Mean gray value was used to quantify the protein expression levels of COL2A1 and SOX9 in the chondrogenic pellet. (F) Schematic of the experimental outline. BMSCs treated with dimethyl sulfoxide (DMSO) or wedelolactone were mixed with the hydrogel and transplanted into the cartilage defect model, and the joint tissues were collected after 6 weeks. (G) Safranin-O fast green stains of joints after 6 weeks (n=5 rat per group). The Osteoarthritis Research Society International scoring system was used to grade the rat cartilage degeneration. Data are expressed as the mean±SD (n=3). Statistical differences were analyzed by one-way ANOVA followed by Dunnett’s test: **p<0.01, ***p<0.001.
![]()
To confirm the ability of wedelolactone to promote cartilage regeneration in vivo, rat BMSCs were induced with or without wedelolactone in chondrogenic differentiation medium for 3 days. The cells were then mixed with hydrogel and injected into the cartilage defect rat models (Fig. 2F). After 6 weeks, the Safranin O and Fast Green staining results of the obtained joint tissues showed that wedelolactone-treated cells had better repaired cartilage defects than non-treated cells (Fig. 2G). In summary, these results indicate that wedelolactone promotes the differentiation of rat BMSCs into chondrocytes in vitro and cartilage repair in vivo.
Wedelolactone promotes the chondrogenic differentiation of MSCs by activating the FOXO pathway
To explore the mechanism by which wedelolactone regulates the chondrogenic differentiation of MSCs, we performed RNA-seq and iTRAQ-based quantitative proteomic analysis of hiPSC-derived MSCs and chondrogenic-differentiated hiPSC-derived MSCs treated with dimethyl sulfoxide or wedelolactone. The iTRAQ analysis results showed that 1,108 proteins were upregulated and 185 proteins were downregulated upon wedelolactone treatment (Fig. 3A). The RNA-seq analysis revealed that 812 genes were upregulated (Supplementary Fig. S2A) and hypertrophic marker genes (MMPs and Aggrecanases) were downregulated (Supplementary Fig. S2B) upon wedelolactone treatment. The KEGG analysis demonstrated that the differentially expressed genes or proteins were enriched in the FOXO signaling pathway (Fig. 3B; Supplementary Fig. S2C). Interestingly, the FOXO signaling pathway factors were upregulated in the wedelolactone-treated group (Fig. 3C). Studies have reported that wedelolactone enhances osteoblastogenesis by regulating the Wnt and extracellular signal-regulated kinase (ERK) pathways (9, 22). However, the GSEA results revealed that wedelolactone has no effect on the WNT, ERK, or NF-κB signaling pathways during chondrogenic differentiation (Supplementary Fig. S2D).
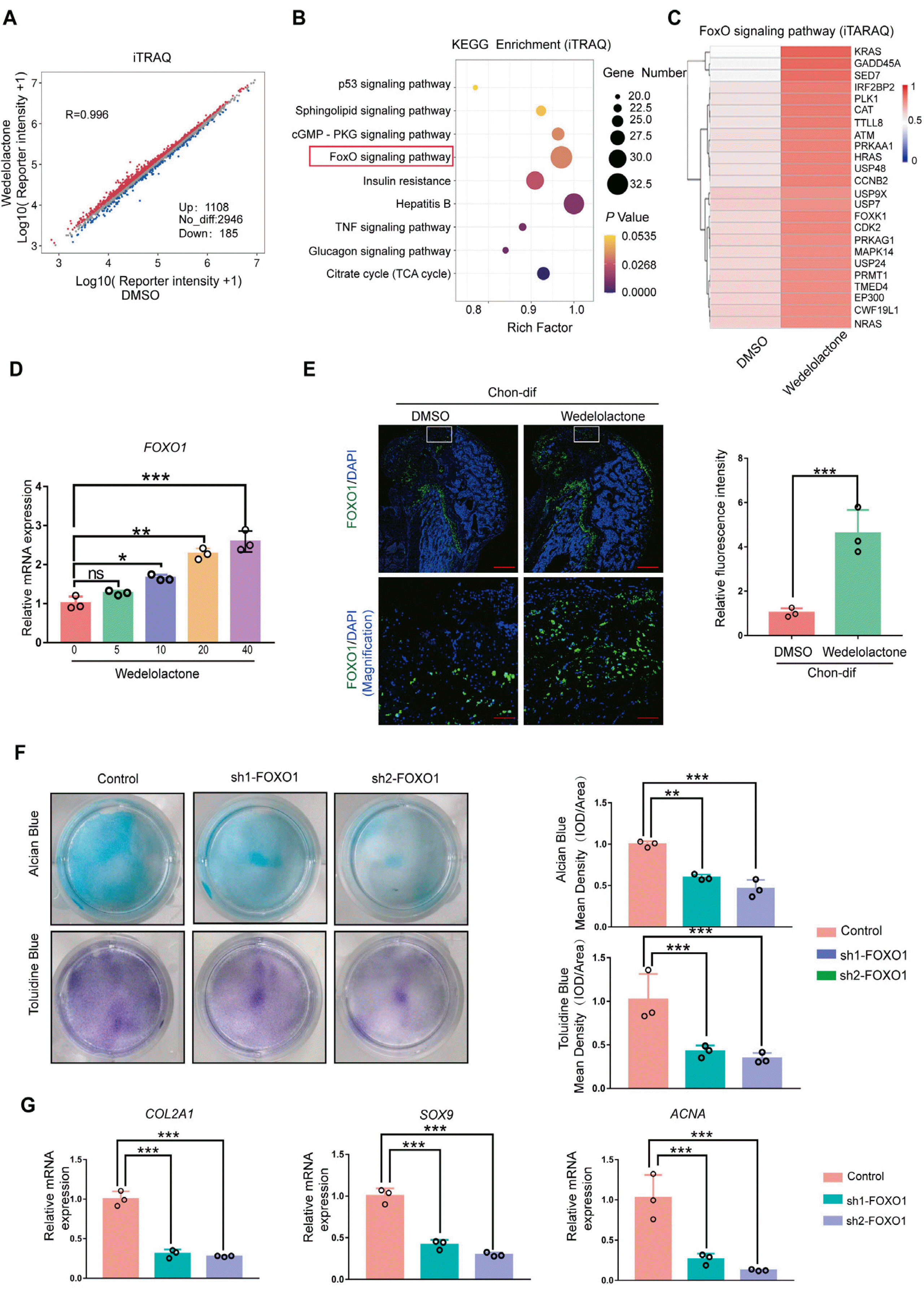
Fig. 3
Wedelolactone promotes the chondrogenic differentiation of MSCs by activating the FOXO pathway. (A) Scatter plot of the quantitation proteomics analysis showing differently expressed proteins in the chondrogenic pellet differentiated from human iPSC-derived MSCs treated with DMSO or wedelolactone. (B) The Kyoto Encyclopedia of Genes and Genomes enrichment analysis of differentially expressed proteins in the chondrogenic pellet differentiated from human iPSC-derived MSCs treated with DMSO or wedelolactone. (C) Heatmap of the quantitation proteomics analysis showing differently expressed proteins associated with FOXO signaling. (D) Gene expression analysis of FOXO1 after induction with different concentrations of wedelolactone. (E) Immunofluorescence staining of FOXO1 in the cartilage defect model after wedelolactone intervention. Scale bar=100 μm. Relative fluorescence intensity was used to quantify the expression of FOXO1. (F) Alcian blue and toluidine blue staining images of the chondrogenic differentiation of human iPSC-derived MSCs after different interventions. Quantification of the mean intensity of alcian blue and toluidine blue staining. (G) Gene expression analysis of the chondrogenic differentiation markers (COL2A1, SOX9, and ACAN) after FOXO1 knockdown. Data are expressed as the mean±SD (n=3). Statistical differences were analyzed by one-way ANOVA followed by Dunnett’s test: *p<0.05, **p<0.01, ***p<0.001.
![]()
Recent research has shown that FOXOs are key factors in stem cell maintenance and differentiation (23). We observed that wedelolactone increased the level of expression of the gene FOXO1 during chondrogenic differentiation in vitro (Fig. 3D). Moreover, in the cartilage defect rat models, wedelolactone-treated rat BMSCs expressed more FOXO1 in vivo than non-treated rat BMSCs (Fig. 3E). To investigate the effects of FOXO1 on chondrogenic differentiation, we silenced FOXO1 in hiPSC-derived MSCs during chondrogenic differentiation. Alcian blue and toluidine blue staining results showed that glycosaminoglycan formation was decreased upon FOXO1 knockdown (Fig. 3F). qRT-PCR results showed that the levels of expression gene chondrogenic-marker genes (COL2A1, ACAN, and SOX9) were significantly decreased in FOXO1-knockdown cells (Fig. 3G). Additionally, AS1842856, a specific FOXO1 inhibitor, significantly decreased glycosaminoglycan formation in iPSC-derived MSCs compared with the control group (Supplementary Fig. S2E). Moreover, the levels of expression of chondrogenic-marker genes were significantly decreased after AS1842856 treatment (Supplementary Fig. S2F). These findings prove that FOXO1 is an important regulatory factor in chondrogenic differentiation.
The promotion of chondrogenic differentiation by wedelolactone is weakened by FOXO1 inhibition
To further confirm that FOXO1 is a key regulatory factor in the wedelolactone-mediated regulation of chondrogenic differentiation, we tested the effects of AS1842856 on wedelolactone-treated cells. Alcian blue staining and toluidine blue staining results showed that AS1842856 significantly decreased glycosaminoglycan formation induced by wedelolactone during chondrogenic differentiation. Furthermore, wedelolactone did not alleviate the inhibitory effect of the FOXO1 inhibitor during chondrogenic differentiation (Fig. 4A). qRT-PCR results showed that the levels of expression of chondrogenic-marker genes (COL2A1, ACAN, and SOX9) were significantly increased after wedelolactone treatment but completely abolished under wedelolactone and FOXO1 inhibitor combination treatment (Fig. 4B). These results indicate that FOXO1 is a key regulatory factor in the wedelolactone-mediated regulation of chondrogenic differentiation.
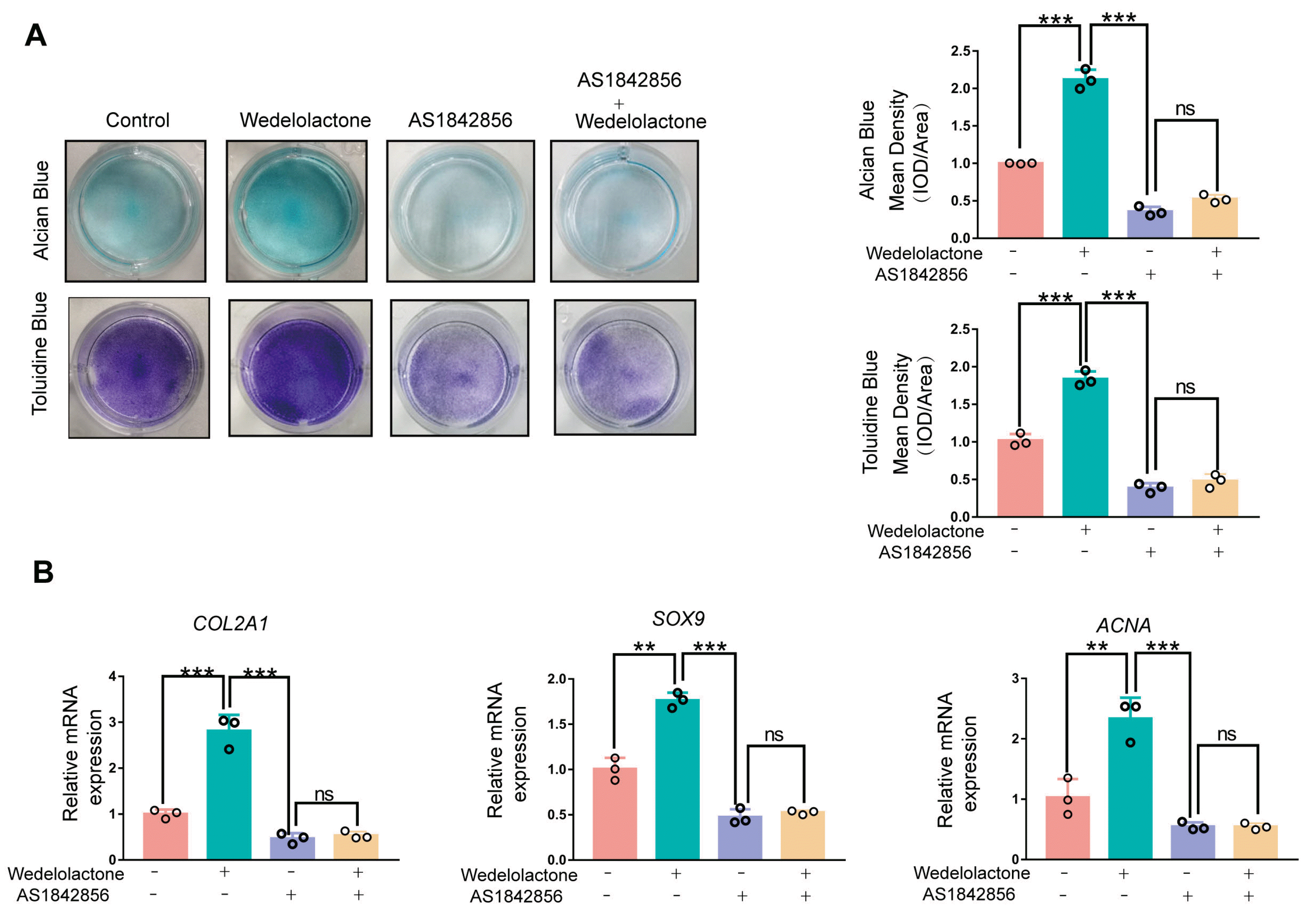
Fig. 4
The promotion of chondrogenic differentiation by wedelolactone is weakened by FOXO1 inhibition. (A) Alcian blue and toluidine blue staining images of the chondrogenic differentiation of human iPSC-derived MSCs after different interventions. Quantification of the mean intensity of alcian blue and toluidine blue staining. (B) Gene expression analysis of the chondrogenic differentiation markers (COL2A1, SOX9, and ACAN) after wedelolactone and FOXO1 inhibitor (GSK126) intervention. Data are expressed as the mean±SD (n=3). Statistical differences were analyzed by one-way ANOVA followed by Dunnett’s test: *p<0.05, **p<0.01, ***p<0.001.
![]()
Wedelolactone decreases EZH2-dependent trimethylation of H3K27 on the promoter region of FOXO1
We next explored the potential mechanisms by which wedelolactone upregulates FOXO1 expression during chondrogenic differentiation. Immunoblot analysis showed that wedelolactone decreased the level of expression of H3K27me3 and increased that of FOXO1 (Supplementary Fig. S3A). Moreover, the ChIP–qPCR results showed that wedelolactone decreased the modification of H3K27me3 on the promoter of FOXO1 in hiPSC-derived MSCs and 293T cells (Fig. 5A). EZH2 is a histone methyltransferase that inhibits gene transcription by trimethylating H3K27. Next, we used GSK126, an effective and highly selective EZH2 methyltransferase inhibitor, to intervene in the trimethylation of H3K27 during chondrogenic differentiation. As expected, GSK126 increased FOXO1 expression at both the mRNA and protein levels (Fig. 5B and 5C).
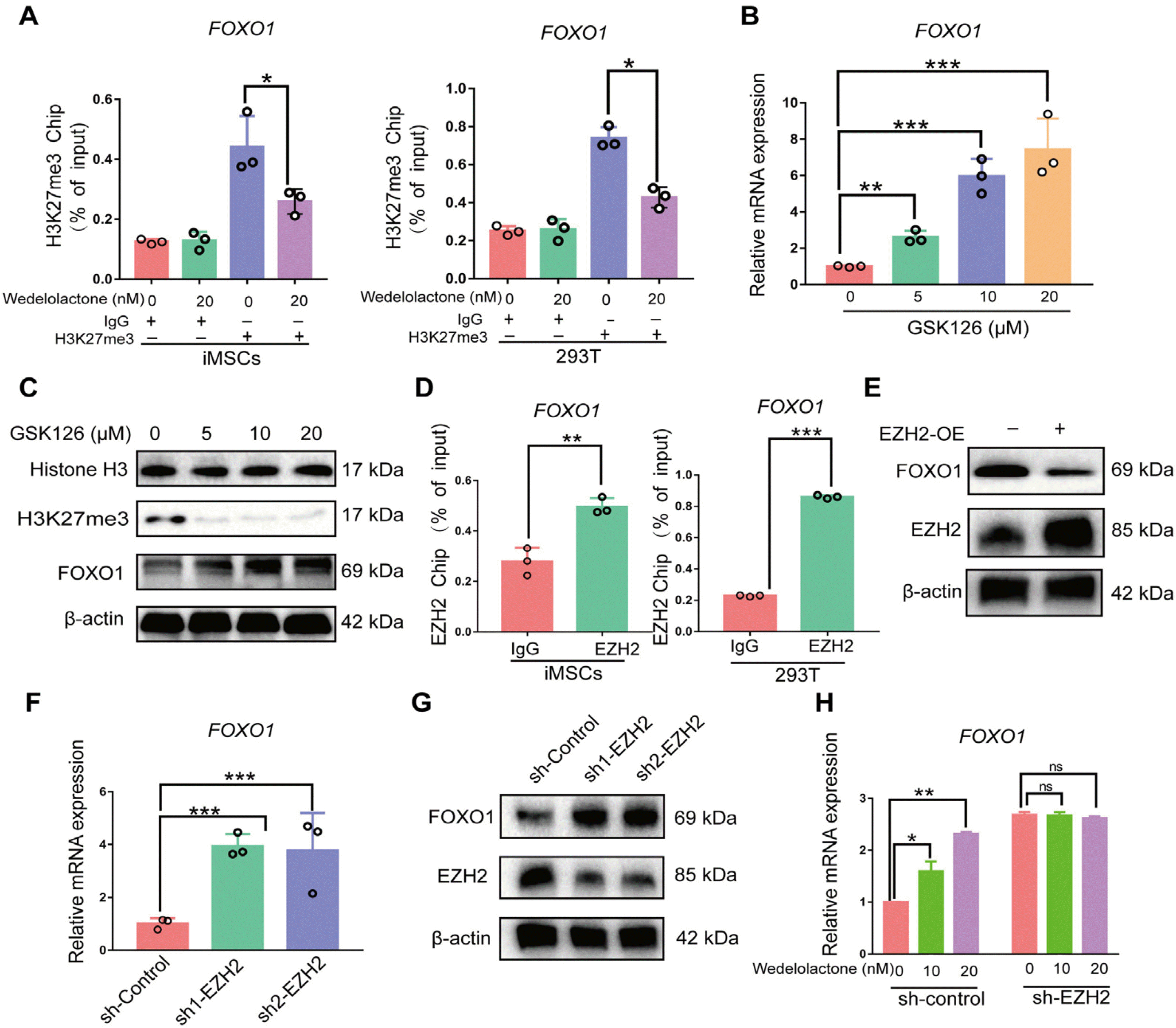
Fig. 5
Wedelolactone decreases EZH2-dependent trimethylation of H3K27 on the promoter region of FOXO1. (A) Chromatin immu-noprecipitation-quantitative polymerase chain reaction (ChIP-qPCR) analysis of H3K27me3 enrichment in FOXO1 promoter after wedelolactone intervention in both human iPSC-derived MSCs and 293T cells. (B) Reverse transcription (RT)-qPCR analysis of FOXO1 in human iPSC-derived MSCs after FOXO1 inhibitor (GSK126) intervention. (C) Western blotting analysis of FOXO1 in human iPSC-derived MSCs after FOXO1 inhibitor (GSK126) intervention. (D) ChIP-qPCR analysis of EZH2 occupancy in FOXO1 promoter in both human iPSC-derived MSCs and 293T cells. (E) Western blotting analysis of FOXO1 in human iPSC-derived MSCs after EZH2 overexpression. (F) RT-qPCR analysis of FOXO1 in human iPSC-derived MSCs after EZH2 knockdown. (G) Western blotting analysis of FOXO1 in human iPSC-derived MSCs after EZH2 knockdown. (H) RT-qPCR analysis of FOXO1 in human iPSC-derived MSCs induced with wedelolactone after EZH2 knockdown.
![]()
To verify whether FOXO1 is a repression target of EZH2 during chondrogenic differentiation, we performed a ChIP–qPCR assay this demonstrated that EZH2 bound to the promoter of FOXO1 in hiPSC-derived MSCs and 293T cells (Fig. 5D). Moreover, EZH2 overexpression in hiPSC-derived MSCs led to the downregulation of FOXO1 expression (Fig. 5E). In contrast, EZH2 silencing in hiPSC-derived MSCs caused a significant upregulation of FOXO1 expre-ssion at both the mRNA and protein levels during chondrogenic differentiation (Fig. 5F and 5G). Wedelolactone had no effect on FOXO1 gene expression levels in EZH2-knoc-kdown hiPSC-derived MSCs (Fig. 5H), suggesting that wedelolactone increases FOXO1 expression by targeting EZH2 in an H3K27me3-dependent manner.
miR-1271-5P interferes with FOXO1 expression during chondrogenic differentiation
miRNAs play important roles in regulating gene expression at the post-transcriptional level. To investigate the potential post-transcriptional regulatory mechanisms of FOXO1, we performed miRNA-seq of hiPSC-derived MSCs with or without wedelolactone added during chondrogenic differentiation (Fig. 6A). Eighteen miRNAs were upregulated and 111 were downregulated upon chondrogenic differentiation (Fig. 6B). Venn diagram analysis showed that two miRNAs (miR-12136 and miR3648-5P) were upregulated after chondrogenic differentiation and further upregulated after wedelolactone treatment (Fig. 6C, Supplementary Fig. S4A). Four miRNAs (miR-1271-5P, miR-22-3P, miR-1296-5P, and miR-361-5P) were downregulated after chondrogenic differentiation and further downregulated after wedelolactone treatment (Fig. 6D, Supplementary Fig. S4B). These six miRNAs may play important roles in the effects of wedelolactone on chondrogenic differentiation.
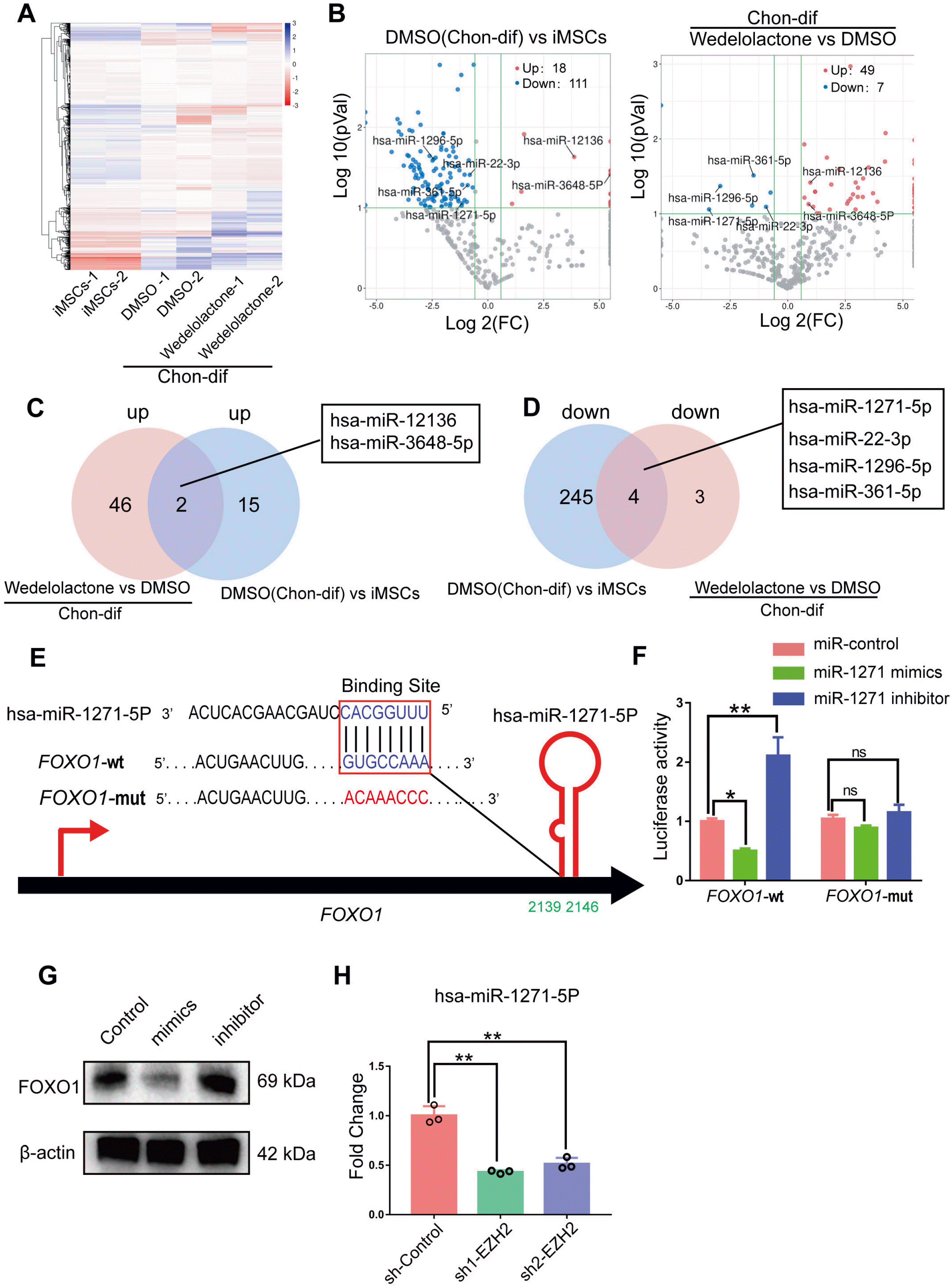
Fig. 6
miR-1271-5P interferes with FOXO1 expression during chondrogenic differentiation. (A) Heatmap of miRNA-seq analysis, showing the differently expressed miRNAs in the chondrogenic pellet differentiated from human iPSC-derived MSCs, and human iPSC-derived MSCs treated with DMSO or wedelolactone. (B) Volcano plots showing the differently expressed miRNAs. Red color indicates the significantly upregulated miRNAs, blue color indicates the significantly downregulated miRNAs, and gray color indicates the genes with no differential expression. (C) Venn diagram overlap showing the number of upregulated miRNAs and miRNAs in the DMSO group compared to the human iPSC-derived MSCs and wedelolactone groups. (D) Venn diagram overlap showing the numbers of downregulated miRNAs and miRNAs in the DMSO group compared to the human iPSC-derived MSCs and wedelolactone groups. (E) miR-1271-5P target sequence in the 3’-untranslated region (UTR) of FOXO1 predicted by the TargetScan database. (F) MiR-1271-5P target 3’-untranslated region (UTR) of FOXO1 was confirmed by the luciferase reporter assay. (G) Western blotting detection of protein expression levels of FOXO1 in human iPSC-derived MSCs transfected with the miR-1271-5P mimic or inhibitor under chondrogenic differentiation. (H) RT-qPCR analysis of miR-1271-5P in human iPSC-derived MSCs after EZH2 knockdown. Statistical differences were analyzed by one-way ANOVA followed by Dunnett’s test: *p<0.05, **p<0.01, ***p<0.001.
![]()
By using the TargetScan and miRDB databases to predict the target genes of these six miRNAs, we found that FOXO1 may bea target gene of miR-1271-5P (Fig. 6E). To test whether miR-1271-5P directly targets FOXO1, we constructed luciferase reporters that had either a FOXO1-3’-UTR-wt or a 3FOXO1-3’-UTR-mut sequence at the miR-1271-5P-binding site. We found that miR-1271-5P overexpression strongly inhibited the luciferase reporter activity of FOXO1 3’-UTR-wt but not that of FOXO1-3’-UTR-mut (Fig. 6F). Further experiments confirmed that miR-1271-5P overexpression markedly decreased FOXO1 protein expression (Fig. 6G). These results indicate that FOXO1 is a target gene of miR-1271-5P. To explore the mechanism by which EZH2 affects miR-1271-5P expression, we silenced EZH2 in hiPSC-derived MSCs during chondrogenic differentiation and found that miR-1271-5P expression was remarkably decreased after EZH2 knockdown (Fig. 6H).
Discussion
Studies have reported that wedelolactone has antihemorrhagic and antiproteolytic activity and that it facilitates osteoblastogenesis but inhibits adipogenesis in MSCs (6, 22). However, the effects of wedelolactone on the differentiation of MSCs into chondrocytes have not been investigated. In this study, we demonstrated that wedelolactone promotes chondrocyte differentiation in both hiPSC-derived MSCs and rat BMSCs. The expression of chondrocyte marker genes, COL2A1 and SOX9, was significantly upregulated upon wedelolactone treatment. Intriguingly, we found that wedelolactone facilitated chondrocyte differentiation by upregulating the FOXO1 signaling pathway via EZH2 suppression, indicating that wedelolactone may promote cartilage repair and increase the cartilage matrix in OA.
Wedelolactone is known to exert regulatory effects on inflammation by suppressing the IkappaB kinase/nuclear factor kappa B (NF-kB) signaling pathway (4). However, the mechanism by which wedelolactone affects stem cell differentiation is not well understood. Studies have demonstrated that wedelolactone enhances osteoblastogenesis by regulating the WNT and ERK pathways (9, 22). In this study, our proteomic data indicate that wedelolactone did not affect the WNT, ERK, or NF-κB signaling pathways of chondrocyte differentiation but decreased the expression of hypertrophic markers in chondrogenic-differentiated MSCs. This discrepancy may be due to the cell type- and context-dependent effects of wedelolactone on stem cell differentiation. The master transcription factor in chondrogenesis is SOX9, which controls the expression of key chondrocyte-specific genes (24). We showed that wedelolactone increased SOX9 expression during the chondrogenic differentiation of MSCs. Moreover, EZH2 catalyzes the deposition of methyl groups on H3K27 for gene silencing (25). A previous study showed that EZH2 overexpression suppressed SOX9 expression in rat endplate chondrocytes (26). This suggests that wedelolactone suppresses H3K27me3 on the promoter of SOX9 during the chondrogenic differentiation of MSCs.
Our data also suggest that wedelolactone potently upregulates FOXO1-dependent signaling pathways. FOXO proteins are transcription factors that play important roles in stem cell differentiation (27, 28). The function of FOXO3A in the chondrogenic differentiation of MSCs has been well studied (29, 30). A recent study and our RNA-seq data showed that FOXO1 expression increased du chondrogenic differentiation. In addition, Kurakazu et al. (31) showed that FOXO1 inhibition leads to cell cycle arrest and chondrogenic differentiation suppression via TGF-β1 signaling. Indeed, we found that inhibition of FOXO1 activity by AS1842856, a specific FOXO1 inhibitor, significantly attenuated the ability of wedelolactone to promote chondrogenesis of MSCs. A previous study reported that ectopic overexpression of EZH2 decreased FOXO1 expression in human oral squamous cell carcinoma cells (32). Based on these reports, we hypothesize that wedelolactone regulates FOXO1 expression during chondrogenic differentiation via EZH2 modulation. To test this, we investigated the mechanism by which wedelolactone treatment regulates FOXO1 expression in chondrogenesis. First, we confirmed that FOXO1 expression increased upon EZH2 inhibitor treatment during chondrogenic differentiation. Furthermore, we showed that EZH2 binds to the promoter of FOXO1. This is consistent with the observation that EZH2 inhibition reduced H3K27me3 on the promoter of FOXO1.
To date, the functions and underlying mechanisms of EZH2 in chondrogenesis have not been completely eluci-dated. During stem cell differentiation or embryonic development, EZH2 mediates the silencing of a diverse group of developmental genes and reshapes the epigenetic landscape in an H3K27me3-dependent manner (33, 34). However, Camilleri et al. (35) showed that EZH2 activity is dispensable for normal chondrocyte maturation. That is, they found that although EZH2 deficiency resulted in a global reduction in H3K27me3 in chondrocytes, conditional knockout mice showed normal cartilage development (35). Another group showed that EZH2 expression was significantly higher in the chondrocytes of patients with OA than in those of healthy individuals (36). Intra-articular injection of EZH2 inhibitor also delayed OA development in a mouse model (36). These findings imply that EZH2 plays physiological and pathological roles in normal or OA chondrocytes by modulating different downstream signaling pathways. Our findings do not rule out the possibility of EZH2 regulating other targets in chondrogenic differen-tiation; however, they highlight the importance of FOXO1 as an EZH2 target in chondrocytes.
Finally, we demonstrated that the noncanonical role of EZH2 as a transcriptional activator for miR-1271-5p expression coexists with its conventional catalytic role as a repressor of the gene FOXO1 in chondrogenic differenti-ation. Interestingly, emerging evidence suggests that there are noncanonical roles played by EZH2 in various cell types. For example, Li et al. (37) showed that EZH2 binds to the promoter of methyltransferase-like 3 to activate its expression in an H3K27me3 independent manner in glioblastoma. Moreover, EZH2 can directly methylate Jarid2 to regulate its transcriptional activity during cell differentiation (38). In our study, depletion of EZH2 by specific short hairpin RNAs decreased miR-1271-5p expre-ssion. Our results suggest that wedelolactone can block the dual roles of EZH2. Therefore, future studies should develop small-molecule inhibitors of EZH2 as they may be effective therapeutic agents for the treatment of OA.
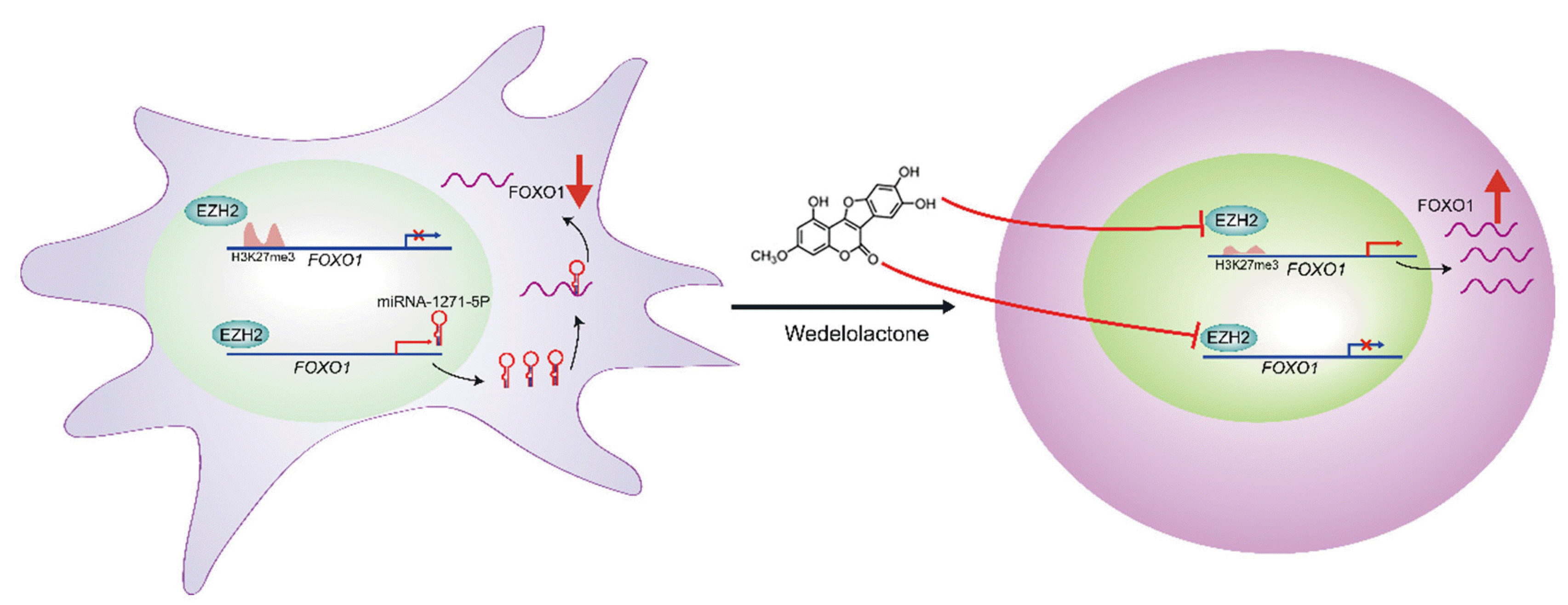
In summary, we demonstrated that wedelolactone promotes chondrogenic differentiation of MSCs by reducing EZH2-mediated H3K27me3 of the promoter region of FOXO1. Additionally, wedelolactone represses miR-1271-5p expression, which post-transcriptionally suppresses FOXO1 expression that is dependent on the binding of miR-1271-5p to the FOXO1 3’-UTR (Fig. 7).
 XML Download
XML Download