

 PDF
PDF Citation
Citation Print
Print


Introduction
External ear anomalies describe a wide range of birth defects such as microtia and anotia, as well as amputation and burns that can result from animal bites, surgery and sometimes cancer (1). Different clinical approaches are used to correct such anomalies and one of the most used is a 2-step method, consisting of harvesting autologous cartilage from ribs and carving it in the shape of the ear before autologous implantation (2). However, this technique possesses some disadvantages such as donor-site morbidity and poor elasticity of the cartilage. On the other hand, synthetic scaffolds and prosthesis are also used to create a proper shape for ear reconstruction. However, these latter were also shown to produce undesirable effects, where they induced inflammation and tissue irritation (3). This explain the everlasting demand for tissue engineering, investigating multiple combinations of cells and scaffolds. Significant advances have been successfully accomplished in regenerative medicine, with various innovative approaches and scaffolds that have been introduced to replace current treatments (4). Many studies have investigated fabricated constructs, where cells were seeded in scaffolds or encapsulated into polymers of cross-linked monomers to be injected later on into ear molds (5, 6). Successful engineering of a clinically auricular implant necessitates a biomaterial, providing tridimensional (3D) system to control and support cellular growth and chondrogenesis. 3D systems are biologically more relevant and thus using such systems, add physiological relevance to cells and optimize their response to the surrounding environment (7). Among them, alginate has been proved to successfully maintain chondrogenic profile and sustain neocartilage regeneration (8). Recent researches highlighted that alginate-hydrogels serve as a stable, cytocompatible, high integrity scaffold in chondral regeneration (9) and auricular 3D bioprinting (10). In addition to 3D environment, oxygen level also plays a key role in chondrogenesis. Indeed, hypoxia, through the nuclear accumulation of HIF-1 alpha levels, promotes mesenchymal stem cells (MSC) chondrogenesis pathway by activating SRY-Box Transcription Factor 9 (SOX9) activity on target genes in mesenchymal stem cells (11-13). Therefore, chondrogenesis is usually conducted under hypoxic environment to enhance the process.
Regarding the cells used for cartilage reconstruction, expanded chondrocytes are employed, usually from autologous sources. However, the low cell density of biopsies, the possible donor-site creation and the propensity of chondrocytes to dedifferentiate upon expansion, led to the widely use of progenitor cells that are characterized by a good proliferation rate and differentiation potential, including under specific conditions, in vitro tridimensional culture, growth factor and peculiar environment (14). Bone marrow mesenchymal stem cells (BMMSC) are well documented and are considered as gold standard for chondrogenic differentiation and high cartilage repair ability especially under hypoxia (12, 15).
Interestingly, several studies have reported the high proliferative rate of tissue progenitor stem cells identified in auricular perichondrium of different species (16, 17). This cell population was characterized by multiple differentiation potential as well as the expression of cell surface markers and colony forming units (18). Perichondrium-derived porcine progenitors were shown to induce cartilage-like matrix (19), both in vitro and in vivo. Furthe-rmore, monkey-derived perichondrial cells transplanted in subcutaneous autologous craniofacial injuries developed mature elastic cartilage after 3 months (20). Another recent study has also suggested the importance of the use of auricular perichondrial cells (AuP) in mice where they produced mature elastic cartilage under chondrogenic conditions (21).
Dental pulp stem cells (DPSC) have also been investigated for cartilage regeneration and repair. When encapsulated in nanopatterned polyethylene glycol and gelatin methacrylate hyaluronic acid (PEG-GelMA-HA) 3D spheroids, these cells upregulated the chondrogenic gene markers and enhanced cartilage tissue engineering (22). Embe-dded in a nanoscale thermosensitive scaffolds, DPSC presented a stable biocompatible construct in vitro and in vivo where they showed a proliferative capacity similar to MSC and retained mechanical strength (23). In other reports, DPSC were evaluated in animal models and their chondrogenic potential for hyaline cartilage (24) and fibrous cartilage repair (25).
Recent literature has also shown that nasal chondrocytes have similar structural composition as articular chondro-cytes, with a good chondrogenic ability since they possess greater cell density and higher amount of glycosamino-glycans (GAGs) (26). They also reported that nasal chondrocytes and neural crest-derived cells successfully repair articular cartilage with a greater chondrogenic capacity than MSC (27). Moreover, do Amaral et al. (28), characterized chondrogenic cell population from human nasal septal cartilage tissue and showed their potential to differentiate into osteogenic and chondrogenic lineages where they observed a high expression of the master gene SOX 9 and also presented cartilage-like extracellular matrix (ECM) in pellet culture system. Other reports have also compared the chondrogenic potential of nasal septal chondrocytes to perichondrial cells and established a novel process to ensure effective and safe clinical use of these cells (29).
In this study, we wanted to determine the best cell candidate to use in order to engineer auricular cartilage for cartilaginous graft preparation. AuP and nasal perichondrial cells (NsP) were chosen as well as DPSC to compare their chondrogenic potential to the gold standard BMMSC. Due to the different embryological origins of these cells from ear cartilage which is mainly derived from neural crests, it was therefore, necessary to investigate and compare progenitors from head area.
Materials and Methods
Isolation and cultivation of human cell sources
Different human cell types were used. BMMSC derived from bone marrow aspirates of patients undergoing hip arthroplasty and having signed the consent form in agreement with the local ethics committee (Comité de Prote-ction des Personnes, CPP Nord-Ouest III). Biopsies were separated on a Ficoll Paque density gradient (Sigma-Aldrich, Saint-Quentin-Fallavier, France) and the ring of mononuclear cells was collected and then seeded in amplification medium: Minimum Essential Medium α with glutamine and sodium pyruvate (MEMα, Dutcher, Berno-lsheim, France) with 10% Foetal Bovine Serum (FBS, Dutcher), 0.5 μg/ml of Fibroblast Growth Factor (FGF, Sigma-Aldrich), and penicillin-streptomycin (Lonza, Levallois Perret, France). The medium was changed three times a week, apart from the first medium change after five days of cell adhesion. The cells were incubated at 37℃ in a humid atmosphere containing 5% CO2. They were cultured and amplified until the 4th passage before use. At this stage, tests are carried out to check the absence of hematopoietic markers (CD34-, CD45-) by RT-PCR.
Human NsP or AuP cells came from surgical wastes within the otorhinolaryngology or maxillofacial surgery surgical units of the Centre Hospitalier Universitaire (CHU) of Caen, from patients (or legals) who signed consent forms. After dissection of the nasal perichondrium, pieces were cut and used for outgrowth cultures. NsP were expanded in MEM α culture medium. After 6 passages, they were freezed at −150℃ for further use.
AuP were obtained from auricular cartilage biopsies of 12-year old children undergoing otoplasty. The perichondrium was separated from cartilage by dissection and sliced into small pieces. Cells were collected by sequential enzymatic digestion using pronase for 45∼60 min at 37℃ (Sigma-Aldrich) and collagenase II overnight at 37℃ (Thermo Fisher Scientific, Illkirch-Graffenstaden, France). The next day, cells were collected by centrifugation and seeded in 75 cm2 surface flasks for expansion until passage 3 and then freezed for further use.
Dental pulp stem cells (DPSC) were obtained from freshly-extracted teeth of young donors (16∼20 years). Teeth were transferred into hypotonic phosphate buffered saline solution (PBS, Lonza), washed and then disinfected by a solution of antibiotics. Then, the pulp tissue was isolated from the chamber and minced into small pieces and sequential enzymatic treatment was carried out by collagenase I (3 mg/ml) and dispase (4 mg/ml) for 30∼45 min, at 37℃ (Thermo Fisher Scientific). After enzymatic digestion, cell suspension was centrifuged at 2,000 rpm for 10 min, and the cell pellet was suspended in culture media (MEM α+10% FBS+0.1% penicillin/streptomycin+FGF 0.5 μg/ml) and expanded until passage 3, and then freezed.
Trilineage differentiation
Cells were tested for their ability to differentiate in vitro towards osteogenesis, adipogenesis and chondrogenesis. Cells were then plated in 6-well plates at a density of 12.5×104 cells/cm2. After 24 hours, they were incubated with appropriate differentiation media. Osteogenic medium: MEM α supplemented with, 0.1% antibiotics, 10% FBS, 100 nM dexamethasone, 50 μg/ml ascorbic acid-2 phosphate, β-glyceraldehyde 10 μM. Adipogenic medium: MEM α with 0.1% antibiotics, 10% FBS, 100 nM dexamethasone, 0.5 mM Isobutylmethylxanthanine. Chondrogenic medium: Dulbecco’s Modified Eagle Medium high glucose with glutamine and sodium pyruvate (DMEM, Dutcher), 0.1% antibiotics, 100 nM dexamethasone, 50μg/ml ascorbic acid-2 phosphate, 40 μg/ml proline, 10 ng/ml of Transforming growth factor beta-3 (TGF beta 3) and Insulin Transferrin Selenium media supplement, 1X (ITS+1). (In this part, all materials are from Sigma-Aldrich, unless mentioned).
Cells were incubated for 21 days with media change twice per week and then fixed with 4% paraformaldehyde (PFA) before cytological staining with Alizarin red, Oil red and Alcian Blue to reveal osteogenesis, adipogenesis and chondrogenesis, respectively.
Tridimensional culture and chondrogenic conditions
Cells were used after the fourth passage, collected by centrifugation and mixed with 1% alginate solution (5× 106 cells/ml). The suspension was extruded into calcium chloride solution (0.1 M) to produce gel beads that were rinsed thereafter with NaCl (0.15 M). The beads were then separated into desired conditions and cultured in 6-well plates dishes, in both normoxic (21% O2) and hypoxic (3% O2) conditions with or without chondrogenic medium for 2 weeks.
Cytological stainings
After 2 weeks, alginate beads were fixed in 4% PFA for 30 min and rinsed overnight with 30% sucrose solution. The next day they were embedded in Optimal Cutting Temperature (OCT) mounting medium and cryosectioned on Leica CM 3050S (Leica Microsystems, Nanterre, France) at −21℃. Sections of 10 μm were collected on super frostⓇ plus microscopic slides (Thermo Fisher Scientific) and air-dried before histology and immunostaining. Safranin O and Alcian blue staining were applied to stain glycosaminoglycans and proteoglycans, respectively.
Immunohistochemistry
Slides were rinsed in PBS and incubated overnight at 4℃ with anti-elastin (1:100, # ab21607 abcam, Cam-bridge, UK), anti-collagen II (1:100, # ab34712 abcam) and anti-aggrecan (1:200, # ab3773 abcam) primary antibodies. A negative control was performed by replacing primary antibody solutions with PBS. The next day, slides were rinsed five times in PBS to remove traces of primary antibody, and a secondary antibody (Alexa Fluorophore 594 conjugated anti-rabbit) diluted (1:800) was used to incubate slides for 1.5 hour. Rinsing with PBS was done to remove excessive secondary antibody. Slides were then rinsed three times with PBS before mounting for microscopic observation by EVOS cell imaging system (Thermo Fisher Scientific).
RNA extraction and real time RT-PCR
After incubations, alginate microspheres were dissolved in a solution of sodium citrate (0.1 M) and total RNA was extracted on cell pellets, by Qiagen RNeasyⓇ mini kit (Qiagen, Courtaboeuf, France), according to the manufacturer’s protocol. Total RNA (1 μg) was treated with DNAse before reverse transcription into cDNA using Moloney Murine Leukemia virus M-MLV enzyme (Invi-trogen by Thermo Fisher Scientific) and oligodT primers. Specific transcripts were then amplified by real time PCR on StepOne Plus apparatus (Applied Biosystems by Ther-mo Fisher Scientific) using Power SYBR Green mix (Thermo Fisher Scientific) and the following primer sequences listed in Table 1.
Table 1
List of primers used
![]()
The relative expression was determined with the 2-∆∆CT method, using geometric mean of 3 housekeeping genes (GAPDH, RPL13 and β2-MG) for normalization.
Results
Trilineage differentiation ability of cell progenitors
In this study, we wanted to compare the potential of different progenitors to produce cartilaginous and elastic matrix in comparison to MSC isolated from human bone marrow as a gold standard for chondrogenesis. These cells are simple to harvest and known for their high accessibility and low risk of tumorigenicity. Moreover, progenitors were derived from head and neck area, i.e. nasal and auricular perichondrium and dental pulp. Nasal perichondrial cells are expandable, stable cell progenitors that can be expanded to more than 30 passages without losing their phenotype. Auricular perichondrial cells express high levels of elastin and are able to produce cartilage ECM proteins. Nevertheless, Dental pulp stem cells are characterized by their unlimited -self renewal as well as their multipotent differentiation ability.
These cells were tested first for their ability to trigger chondrogenesis, osteogenesis and adipogenesis in vitro. Cytological staining showed their potential to evolve towards the trilineage, indicating an undifferentiated status (Fig. 1, Supplementary Fig. S1). Differences in staining intensity were shown depending on cell types. Although not quantitative, they suggested however some differentiation tendencies towards cartilage and bone tissue.
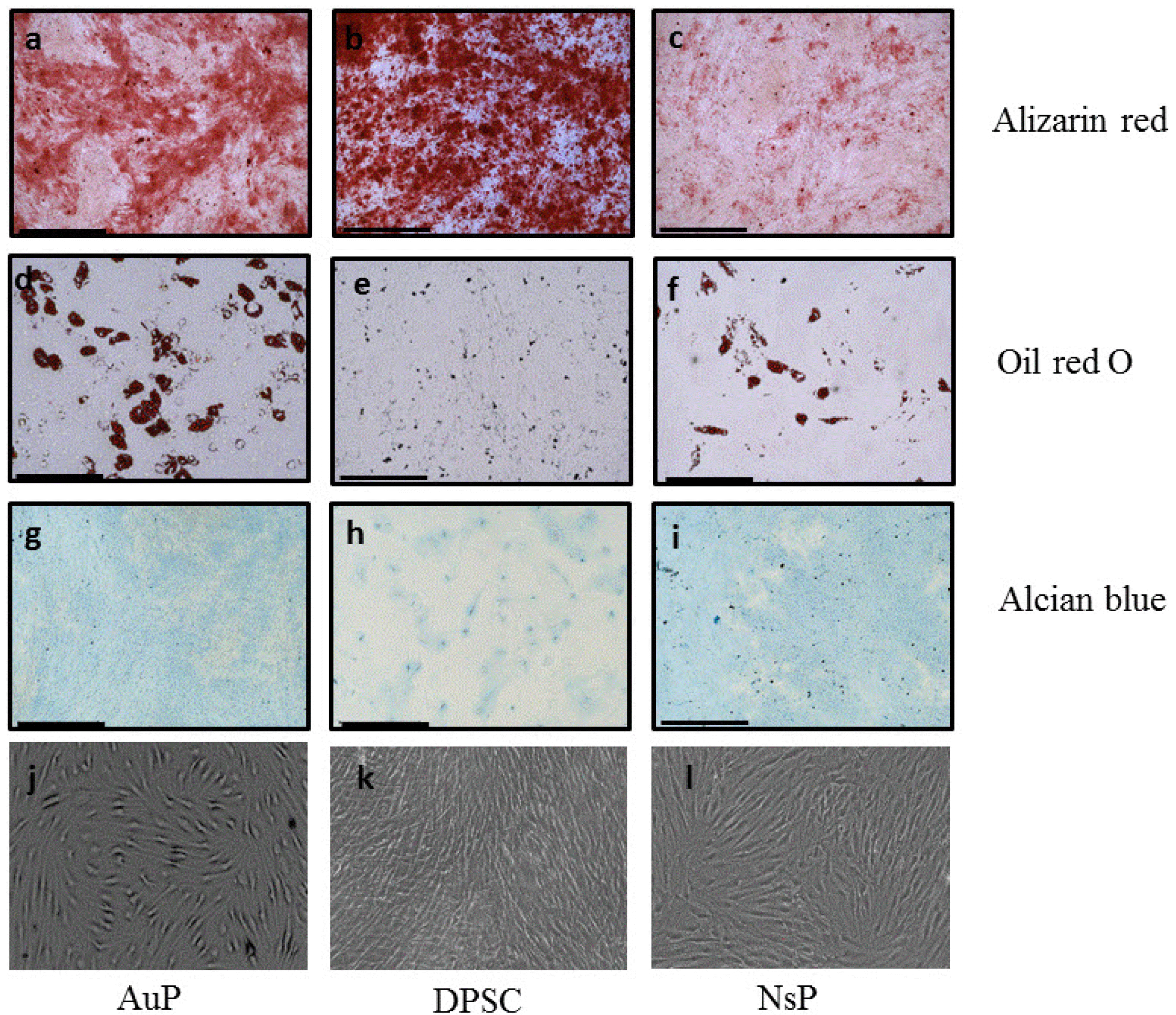
Fig. 1
Multilineage differentiation potential of human progenitors. The trilineage differentiation of AuP, DPSC and NsP were carried out by Alizarin red (a∼c), oil red O (d∼f), and alcian blue (g∼i), respectively. (j∼l): Bright field images of the cells. These results were compared to a control (Supplementary Fig. S1). Scale 500 μm.
![]()
Chondrogenic differentiation in 3D alginate model
Next, a comparative chondrogenic study of different cells was performed in a well-mastered 3D hydrogel model (alginate beads) in comparison with human BMMSC. Cellularized beads were cultured under chondrogenic media in normoxic (21% O2) or hypoxic environment (3% O2) for 2 weeks. Beads were then analyzed by cytological staining for ECM production and cell morphology (Fig. 2). Alcian blue is known to stain proteoglycans while safranin O stains cartilage and mucins. After 2 weeks of culture, all cell types were able to produce cartilaginous ECM as shown in histological staining.
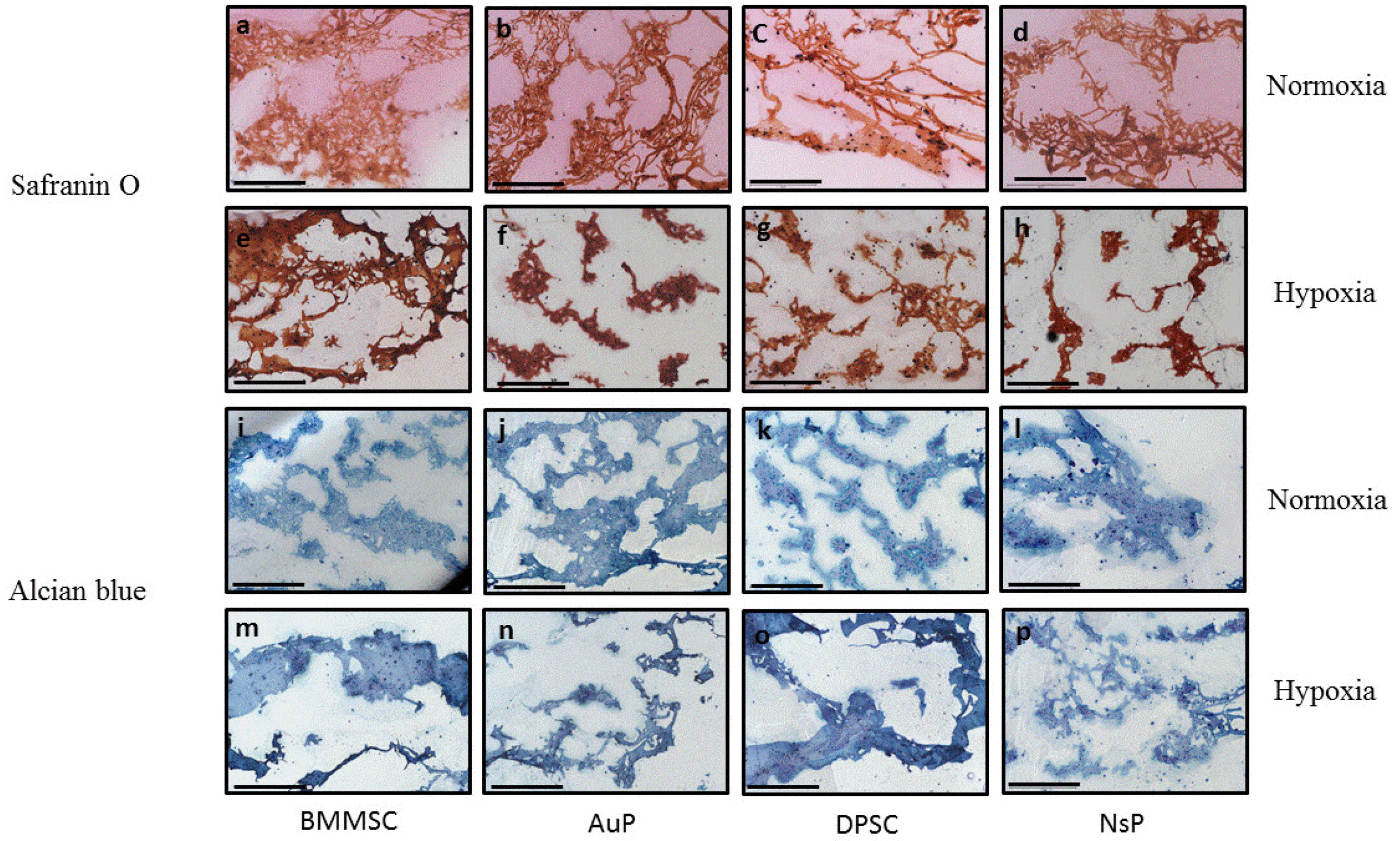
Fig. 2
Production of GAGs and glycoproteins after 2 weeks of culture. Safranin O and alcian blue staining were respectively performed for illustrating the positive production of proteoglycans (red) and glycosaminogly-cans (blue) in bone marrow-derived mesenchymal stem cells (BMMSC) a, e, i, m, auricular perichondrocytes (AuP) b, f, j, n dental stem cells (DPSC) c, g, k, o, and nasal perichondrocytes (NsP) d, h, l, p, encapsulated in 3D alginate microsphere in normoxic and hypoxic conditions, respectively, after 2 weeks of culture in chondrogenic medium (scale bar 500 μm).
![]()
Hypoxia differentially affects chondrogenesis of progenitors
Gene expression analysis was also conducted after the same treatment, targeting cartilaginous matrix markers. The expression of cartilage markers genes (COL2A1, ACAN, SOX9), elastin (ELN) and unspecific collagen type I (COL1) as well as hypertrophic marker collagen type X (COL X) was analyzed by real-time RT-PCR after normalizing data against three housekeeping genes (RPL13, GAPDH and beta2-MG) (Fig. 3). After 2 weeks of chondrogenic differentiation under normal atmosphere, even all cell types were able to trigger the expression of cartilage markers and ELN, AuP cells appear more efficient. They also showed more interesting results, regarding undesirable genes, since expression of COL1 and COLX were not increased by chondrogenic medium, compared to other progenitors.
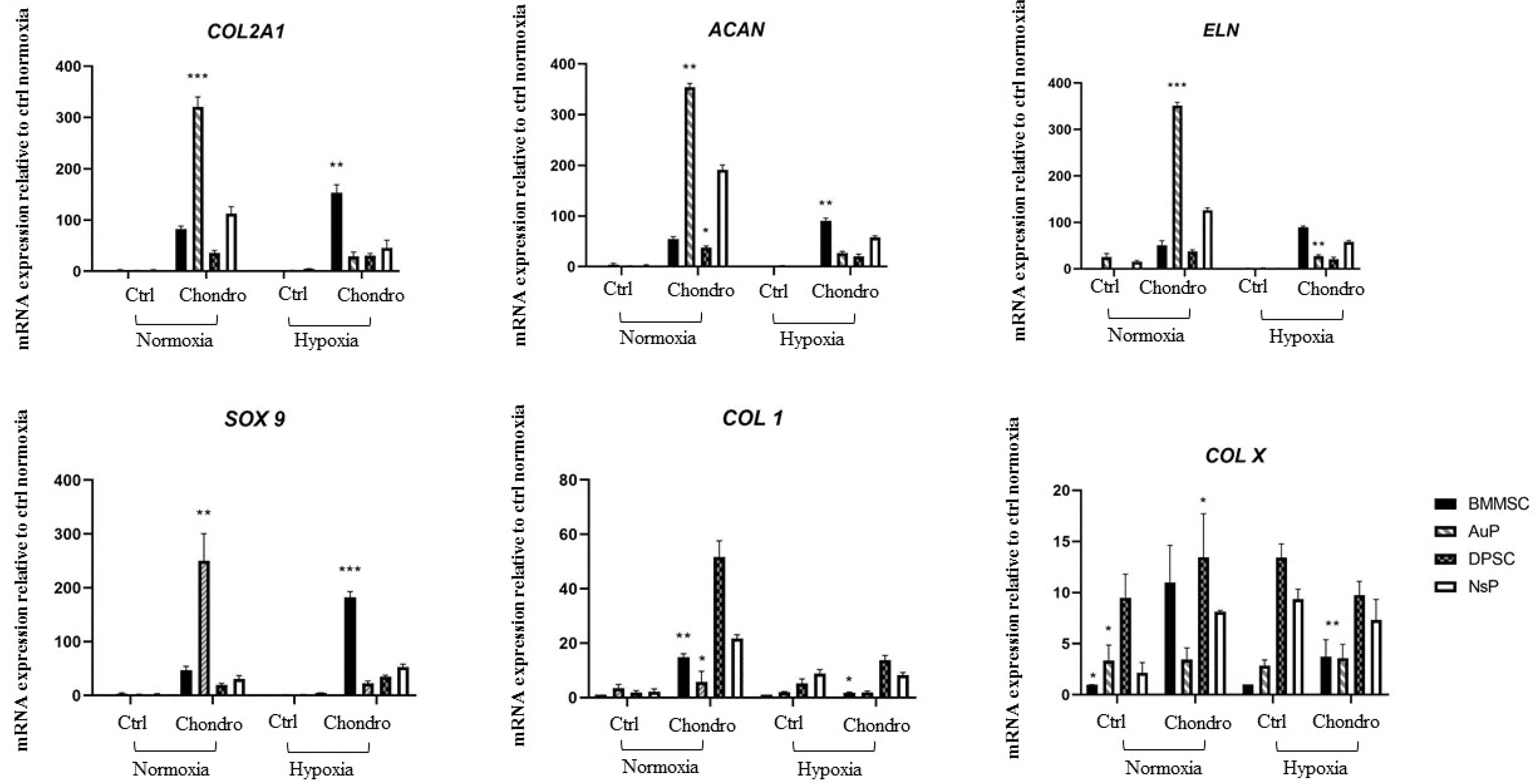
Fig. 3
Expression of cartilage genes in four different cell sources in Normoxic and hypoxic environments. RT-PCR analysis were performed to examine cartilaginous markers in 4 different cell lines in normoxic and hypoxic conditions. The expression of these markers was compared to the average of the 3 housekeeping genes. *p<0.05, **p<0.01, ***p<0.005.
![]()
Under hypoxia and as expected, enhanced cartilage ma-rkers gene expression (COL2A1, ACAN, SOX9, ELN) is observed for BMMSC cells with low expression of COL1 and COLX. Interestingly, even chondrogenic medium enhanced cartilaginous markers in the other progenitors, it appears that hypoxia doesn’t improve this effect, regarding the results under normoxic condition (Fig. 3). In the same time, COL1 and COLX are also differentially regulated, with very high expression in nasal and dental progenitor, no matter under hypoxia or not. This suggests that hypoxia has different mechanisms of action in controlling chondrogenesis depending on the cell progenitor source used.
Taken together, these results indicate that among the cells tested, AuP cells, highly express cartilaginous genes and that hypoxia is not needed to promote their chondro-genesis.
Chondrogenesis of auricular progenitors is less affected by hypoxia than BMMSCs
The expression of type II collagen, aggrecan and elastin were subsequently observed on sections of cellularized alginate beads by immunohistochemistry. Regarding the high expression of COL1 and COLX in NsP and DSP cells, we limited the test to AuP and compared them to BMMSC. Negative controls were performed to ensure that secondary antibody binding is specific (Fig. 4A). As expected, both cell types were able to produce cartilage specific proteins (type II collagen, agrecan and elastin, respectively Fig. 4B∼4D). Although the test is not quantitative, we notice that hypoxia doesn’t impact the expression of the targeted proteins in AuP, contrary to BMMSC. This is particularly glaring when regarding type II collagen staining (Fig. 4D).

Fig. 4
Immunohistochemistry of ELN, COL II and ACAN in presence and absence of oxygen tension. (A) Negative control was illustrated where primary antibody was replaced by PBS (a∼c). (B∼D) Type 2 collagen, aggrecan and elastin expression was detected in auricular perichondrocytes and bone marrow-derived mesenchymal stem cells in presence of high (a∼c, g∼i) and low oxygen tension (d∼f, j∼l), respectively (scale bar 200 μm).
![]()
Discussion
The aim of this study was to evaluate the ability of different cellular sources to form and better express cartilage genes and to investigate whether hypoxia can affect their chondrogenesis. The cells progenitors were chosen for their ability to differentiate into cartilage and their origin area, in comparison with human BMMSC as gold standard for chondrogenesis. These latter were used regarding several previous studies and as expected, when encapsulated in 3D alginate microspheres they presented the highest expression of cartilaginous genes and a dense proteoglycans aggregates in 3% O2 hypoxic environment, which is correlated with previous studies that demonstrated that human BMMSC grown in 3D with extended hypoxia increased the expression of cartilaginous genes (30). It also corroborates our previous studies showing that hypoxia induced the differentiation of BMMSC towards cartilage-like cells, with an upregulated expression of COL2A1, ACAN and SOX transcription factors with no hypertrophic markers in both in vitro and in vivo conditions (11). Other groups have also investigated that immortalized murine MSC incubated in 1% oxygen increased the levels of chondrogenesis markers SOX 9, ACAN, COL2A1 (31). More-over, hypoxic culture of BMMSC enhanced in vitro and in vivo chondrogenesis capacity, proliferation and tissue formation (11, 32). The choice of alginate hydrogel to recapitulate 3D environment, was based on its porosity, flexibility and elasticity. It is biocompatible and easily injectable in minimal invasive manner, which leaves it very interesting in tissue engineering applications. In addition, this biomaterial supports chondrogenesis and stem cell differentiation into cartilage cells (11, 33-38).
Human BMMSC are the gold standard cells and a well mastered source for chondrogenesis. However, their embryological origin is different from auricular cartilage. To fully engineer auricular tissue, progenitors from head and neck area should be investigated for their chondrogenic ability to form robust cartilage. Hence, we used cartilage-derived progenitors from ear and nose perichondrium, since they can generate large number of cells and represent a promising source for tissue engineering based cell therapy (39).
Human nasoseptal derived perichondrium were also tested for their ability to form cartilage. Staining results showed a better proteoglycans and collagen amount under standard oxygen tension compared to other cell sources. Also, these cells presented a good expression profile of elastic cartilage. Although NsP cells expressed ELN, SOX 9, ACAN and COL2A1, that expression was significantly lower than that of AuP cells. A similar pattern of results was obtained with auricular and nasoseptal chondrocytes as cell sources for regeneration of cartilage. Chondrocytes originating from both nasal septum and auricles showed potent proliferation in either monolayer or 3D conditions, suggesting that auricular chondrocytes produced large pellets with more cartilage-like matrix and had a higher expression of anabolic growth factors than nasoseptal chondrocytes (40). However, both cell types didn’t expressed COLX gene, in agreement with our results.
AuP were also compared to progenitors from dental pulp. Recent reports have described the potential of dental stem cells for stem cell therapies in articular cartilage repair (41). These cells contained microvilli-like structures on their surfaces, permitting a good attachment to a specific scaffold. Once grown in chondrogenic medium, dental stem cells presented a rounded morphology that was correlated with ACAN and COL2A1 expression (42). Another study reported that formation of collagen fibers by dental stem cells, confirming their ability to differentiate into chondrocyte within matrices (43). In our experiment, the chondrogenic differentiation was weaker compared to BMMSC and AuPs, corroborating earlier study showing that 3% hypoxia suppressed differentiation of dental stem cells. In fact, the precise O2 tension is important to trigger cell responses. Comparison of different O2 levels (3 up to 20%) reported different behavior and gene expression, in line with previous reports, that suggested that hypoxia boosted bone healing process and promoted VEGF expression in dental stem cells, as well as bone formation, collagen ECM arrangement and numerous mineralization modules (44). Our results showed that after 14 days of culture, both in normoxic and hypoxic conditions, dental stem cell showed the highest expression of COL1, suggesting more an osteoblastic profile rather than chondrogenic ability. These is consistent with finding that showed that hypoxic condition promoted mineralization and differentiation of dental pulp cells of the odontoblastic layer (45). Hypoxia was also shown to be an effective treatment to expand numbers of progenitor cells and to enhance angiogenic potential and affect the differentiation ability of odontoblasts (46).
Auricular progenitors were also described as multipotent with the ability to differentiate towards multiple lineages. Progenitors from auricular perichondrium were already described to be good candidates for ear cartilage produ-ction (21, 39). However, to date, no study has evaluated the effect of hypoxia on these cells.
In this study, AuP cells presented good ability for chondrogenesis where they highly produced elastic cartilage genes (ELN, COL2A1, ACAN, SOX9) with low expression of COL1 and COLX. It is also noteworthy that the best effect is obtained in normoxic conditions even though hypoxia strongly modulated the chondrogenesis of several other progenitors, including human BMMSC. The fact that auricular progenitors can be driven towards chondrogenesis without hypoxic condition is of importance in the scope of tissue engineering or cell therapies.
Aside to the remarkable increase of elastic cartilage marker ELN, the relatively low expression of COLX in auricular progenitors indicates the preservation of the phenotype in these cell types under in vitro chondrogenic conditions.
Similarly, the chondrocyte dedifferentiation marker, COL1 was also mostly reduced in AuP, both in normoxic and hypoxic conditions. It is widely known that human perichondrium tissue initially contains COL1. When these cells were encapsulated in alginate 3D beads, COL1 expression was significantly decreased whith a low expre-ssion, confirmed by our results in normoxia as well as in hypoxia. This indicated that 3D encapsulation of AuP suppressed the expression of COL1 and increased elastic cartilage phenotype.
On the contrary, it has been shown that cells derived from the cartilage tissue appear more apt to differentiate towards chondrogenic and osteogenic lineages, whereas perichondrium-derived progenitor cells differentiate more into adipose tissue (18). This wasn’t the case in our work since AuP turned out to be the best cell source to produce elastic and non-hypertrophic cartilage when cultured in alginate beads under normoxic conditions. Also, MSC from both bone marrow and adipose tissue, despite a high proliferation capacity, appear not to be the optimal cell source for cartilage due to a limited chondrogenic differentiation and development of hypertrophic cartilage (19).
Immunohistochemistry results were consistent with gene expression data. Indeed, hypoxia suppressed cartilaginous activity of auricular progenitor cells on one hand, and stimulated the significant increase of BMMSC on the other hand. Overall, these finding are in accordance of results by Kobayashi et al. (16) who identified human progenitor auricular perichondrium cells as highly proliferative and able to differentiate into multiple lineages and produce an elastic mature cartilage rich with elastic fibers and proteoglycans, when subjected to basic fibroblast growth factor (bFGF) and Insulin –like growth factor 1 (IGF1).
In conclusion, AuP are the best cell candidates for elastic cartilage engineering as previously described. The novelty of this study is that these cells highly express elastic cartilage markers in normoxic conditions. Yet, hypoxia, revealed ineffective and useless condition for these cells, unlike other progenitors. These results should be taken into account for tissue engineering of elastic cartilage and extended by performing studies in animal models for further investigation of in vivo of cartilage repair.
 XML Download
XML Download