

 PDF
PDF Citation
Citation Print
Print


Introduction
Oxygen is a key regulator of cellular functions in stem cells and lowered oxygen tension of approximately 3% is beneficial for stem cell proliferation and maintenance in neural stem cells (NSCs) (1, 2). Sensitivity to oxygen seems to vary between NSCs from different fetal brain regions with NSCs isolated from the midbrain showing a selective vulnerability to oxygen (1, 3). Such observations are not surprising as oxygen tension in vivo is almost always substantially lower than atmospheric oxygen tension (21% O2): The physiological oxygen levels within the mammalian brain range from 1% to 5% O2 (herein called physioxia (4) and thus differ remarkably from the atmospheric oxygen condition routinely used in in vitro culture.
The major oxygen sensing molecule regulating the molecular response to hypoxia is the hypoxia inducible factor (Hif) (5). Hif-1α is by far the best characterized Hif family member and is considered a key regulator of oxygen homeostasis (6). Expression of Hif-1α is crucial for normal development of the brain as shown in neural-specific Hif-1α conditional knock-out model (7, 8). Moreover, in our previous study we demonstrated a selective vulnerability of predopaminergic midbrain NSCs (mNSCs) against the knock-out of Hif-1α leading to impaired proliferation, survival and dopaminergic differentiation of mNSCs, while cortical NSCs were not affected (3).
The Notch pathway is one of the key factors for stem cell maintenance and cellular fate decisions. Extensive research of the Notch pathway in vitro and in mouse mutants revealed that Notch maintains the NSC state and regulates their number and activity (9, 10). Recent studies identified specifically Hes1 oscillations as key mechanism enabling NSCs to switch their fate (11, 12). Overexpre-ssion of Hes1, which presumably enables Hes1 oscillations, and also of Notch was found to inhibit fate decision in neurons and oligodendrocytes while the disruption of the Notch pathway promotes neuronal differentiation and decreases the expression of neural stem cell markers (13, 14). A rapid reduction in cell death of murine NSC upon Notch stimulation by its ligand Dll4 has also been shown (9).
A few studies implicate that Hif-1α physically interacts with Notch intracellular domain (NICD) during hypoxia, leading to its stabilisation and enhanced transcription of its targets with subsequent promotion of stem cell maintenance of cortical NSCs and medulloblastoma-derived neuroprogenitors (15, 16). Apart from the recruitment of Hif-1α to the transcriptional complex of Notch, Notch was found vice versa to regulate Hif-1α transcription and to affect Hif-1α protein stability and promote its target gene expression by recruiting Hif-1α to the hypoxia response element (HRE) (15-17). There is evidence that both are needed for stem cell maintenance in murine embryonic stem cells (17). However, the involvement of Notch signalling in murine midbrain stem cells under physiological hypoxic conditions (physioxia) is not well characterized so far. Recent data from transcriptome analysis specifically identified the Notch pathway to be regulated in long-term physioxia cultured mNSCs (18). Transcriptome analysis and our observations that oxygen and Hif-1α knock-out are able to affect mNSC survival and differen-tiation (3) prompted us to investigate the involvement of Notch signalling in physioxia-dependent stem cell performance in a well-established mNSC in vitro cell culture model to further elucidate the molecular mechanisms mediating the selective effects of oxygen in mNSCs during embryonic development (19).
Materials and Methods
Isolation and culture of fetal murine midbrain neural stem cells (mNSCs)
mNSCs were isolated from wild-type or neural-specific Hif-1α conditional knock-out mice as described previously (20). Ventral mesencephalic samples were harvested from E13.5 mouse embryos (C57Bl/6J, Charles-River; Hif-1αflox/flox mice were a kind gift from Shushei Tomita, MD, PhD). Generation and analysis of Nes-Cre Hif-1α conditional knockout embryos has been described previously (7). The samples were disaggregated, incubated in 0.1% trypsin and 10 mg/ml DNase (both from Sigma-Aldrich) and triturated to a quasi-single cell suspension. Cells were re-suspended in NSC medium (65% (v/v) high glucose DMEM, 32% (v/v) F-12, 2% (v/v) B-27 and 1% (v/v) Penicillin/Streptomycin, all Gibco) supplemented with growth factors (20 ng/μl EGF and FGF2, both Sigma-Aldrich) and equilibrated for at least 24 h under the appropriate oxygen condition. Cells were plated onto poly-L-ornithine and fibronectin and incubated at 37℃ in 5% CO2 and 95% air (21% O2, normoxic condition) or under lower oxygen conditions in 5% CO2, 92% N2 and 3% O2 (hypoxic condi-tions) monitored with an O2-sensitive electrode system (Haereus, Germany).
Treatment paradigms
Treatment with the Notch agonist Delta like 4 (Dll4, 1 μM dissolved in 0.1% bovine serum albumin (BSA) in PBS, R&D Systems), the Notch antagonist N-[N-(3,5-Difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester (DAPT, final concentration: 10 μM, dissolved in dimet-hyl sulfoxide (DMSO) at a final concentration of 0.1%, Calbiochem) or with the respective solvents as control was performed for 3 days. For experiments requiring membrane-bound Dll4, mNSCs were co-cultured with the murine stromal cell line Sc9-19, which is known to express high levels of Dll4 (21), for 3 days in mNSCs media under the oxygen conditions stated above. In case of co-culture mNSCs stably expressing yellow fluorescence protein (YFP) were used in order to distinguish the two cell populations.
Immunocytochemistry
Cells were fixed in Accustain (Sigma-Aldrich) for 30 s. Immunocytochemistry was performed using standard pro-tocols. Nuclei were stained with Hoechst 33342 (Invitrogen) and the following primary antibodies: chicken anti-GFAP (1:1,000, Abcam), mouse anti-Nestin (1:500, Chemicon), rabbit anti-Tuj1 (1:500, Covance), rabbit anti-Ki67 (1:500, Novocastra), mouse anti-Map2 (1:500, BD Pharmin-gen) and rabbit anti-NICD (1:500, Abcam). Cells were analysed by conventional epifluorescence microscopy (Leica DM IRE2) and Leica FW4000 software. Image analysis and cell quantification were performed with ImageJ (Wayne Rasband) or Photoshop CS3 (Adobe). A total of 500 to 1,500 Hoechst+ cells were counted per coverslip and at least five coverslips were counted per time point and condition.
Quantitative real time PCR
Isolation of RNA for qRT-PCR was performed essentially as described previously (20). The relative gene expression levels between samples were obtained using the formula of the comparative threshold cycle (ΔCt method) 2(Ct [housekeeping gene] - Ct [target gene]). In case of DAPT, Dll4 treated samples, relative mRNA levels were first normalised to the relative mRNA levels of Hmbs (hydroxyme-thylbilane synthase) and then to their respective control condition. We considered a ≥2-fold change of mRNA levels a relevant change of gene expression. Following primers were used: Hes1 5’-AGG CAG ACA TTC TGG AAA TGA-3’ and 5’-GTC ACC TCG TTC ATG CAC TC-3’, Hes3 5’-CAG GAA ATC CAC GGA ACC-3’ and 5’-CCG TCC CCT CCC ATC TTC-3’, Hes5 5’-GCT CGC TAA TCG CCT CCA-3’ and 5’-GGT CCC GAC GCA TCT TCT-3’, Hey1 5’-CCT GGC CGA AGT TGC CCG TT-3’ and 5’-GCT TCC CGC TGG GAT GCG TAG-3’, Hmbs 5’ TCG GGG AAA CCT CAA CAC C 3’ and 5’ CCT GGC CCA CAG CAT ACA T-3’, Id1 5’-CTG AAC GGC GAG ATC AGT G-3’ and 5’-TTT TTC CTC TTG CCT CCT GA-3’, Vegf 5’-GCT ACT GCC GTC CGA TTG-3’ and 5’ CTC CAG GGC TTC ATC GTT AC-3’.
Microarray analysis
For the microarray gene chip assay, cells were cultured as described above for 48 hours or 13 days post dissection. Isolation of RNA was carried out as described above and RNA quality was approved by Agilent 2100 Bioanalyzer. RNA integrity number was between 9.8 and 10. Hybridi-zation to whole mouse genome microarray gene expression chips (Gene ST 1.0 Arrays, Affymetrix) was performed according to manufacturer’s instructions. Microarray chips were then immediately scanned using an Agilent microarray confocal laser scanner. Microarray data analysis was performed with the Agilent GeneSpring GX11 software. Statistical significance was computed using the unpaired Student’s t-test with the corrected p-value cut off 0.05 and Bonferroni as the multiple testing correction for the displayed genes.
Cell cycle analysis
The cellular distribution in the various cell cycle phases has been determined by flow cytometry using the Cycle-TESTTM PLUS DNA Reagent Kit (BD Biosciences) according to manufacturer’s instructions. Briefly, cells were washed 3 times in 1 ml washing solution at room tem-perature. To disassociate potential cell clusters, cells were incubated in 250 μl Solution A (trypsin buffer) for 10 minutes at room temperature. Trypsin activity was stopped by adding 200 μl Solution B (trypsin inhibitor and RNase buffer) for 10 minutes at room temperature. Cells were stained with propidium iodide (PI) for 10 minutes in the dark at 4°C. About 20,000 cells were analysed per condition and their particular cell cycle phase was distinguished by the total fluorescence intensity (FL2-Area parameter) as following: (G0-G1) cells with a standard fluorescence intensity, (S) cells with an intermediate fluorescence intensity, (G2-M) cells with a double fluorescence intensity. Data acquisition was performed with CellQuest software and FACSCalibur flow cytometer (both BD Biosciences) using a 488 nm argon-ion laser and FL2 585/42 nm filter for PI detection and data analysis was carried out with the software Modfit LT3.0 (Verity Soft-ware House).
Analysis of cell death
To analyse cell viability a mixed solution of Hoechst 33342 (Invitrogen), fluorescein diacetate (FDA, Sigma, 100 μg/ml, Sigma) and PI (120 μg/ml, Invitrogen) was added to the medium and incubated for up to 10 min at 37℃. Upon treatment cells were analysed by conventional epifluorescence microscopy (Leica DM IRE2) and images were acquired using the Leica FW4000 software. Image analysis and cell quantification were performed with ImageJ (Wayne Rasband) or Photoshop CS3 (Adobe). At least five images were analysed per condition and time point.
Results
Physioxia promotes proliferation and inhibits spontaneous differentiation of mNSCs while normoxia enhances the cell population in G0-G1 phase at the expense of S phase
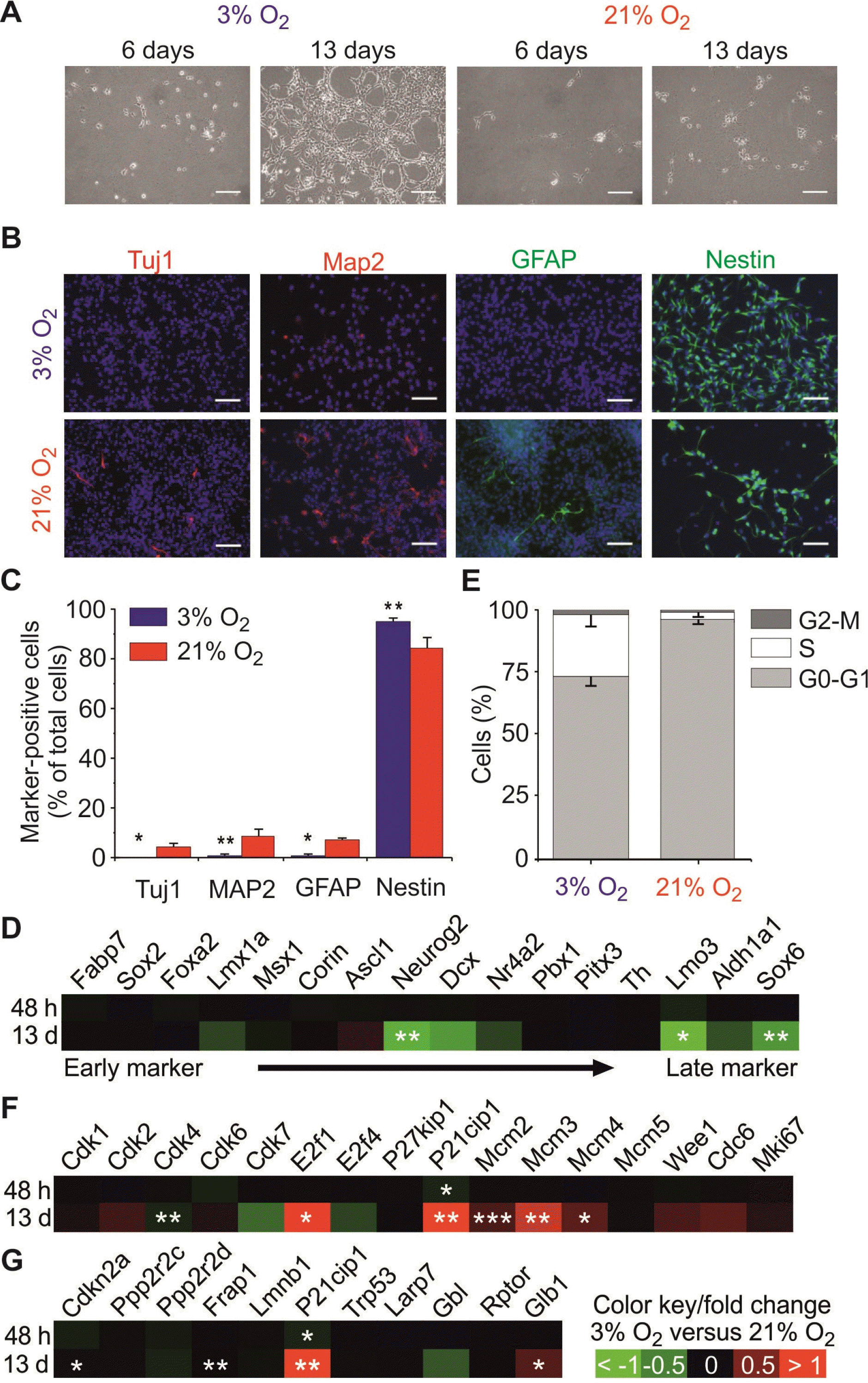
Although physiological hypoxia (physioxia, 3% O2) has been reported to improve the long-term proliferation and survival of mNSCs, little is known about spontaneous differentiation in these conditions. Here, we could observe that physioxia promotes also short-term proliferation of mNSCs when compared to those cultured under normoxia (Fig. 1A and 1B). To investigate whether lowered oxygen prevents spontaneous differentiation, the expression of common markers of the NSC state (Nestin) and their differentiated progenies (Tuj1, Map2, Gfap) complemented by additional markers of mNSC development (22) were used to determine the cell composition in the different culture conditions (Fig. 1B∼D). While cultures under physioxia resulted in a homogenous population of Nestin+ mNSCs without relevant signs of spontaneous differentiation (no expression of Tuj1, Gfap, little expression of Map2), normoxic conditions caused a significant increase in spontaneous neuro-glial differentiation of mNSCs (Fig. 1B and 1C). Fig. 1D displayed the comparative results between 3% vs. 21% oxygen tension of the marker panel for mNSC development from early markers on the left side (Fabp7, Sox2, Foxa2) to late post-mitotic markers on the right side (Th, Lmo3, Aldh1a1, Sox6) (22). We found a delayed increase after 13 days of Neurog2 as a marker for early post-mitotic neuroblasts (22) as well as the late post-mitotic markers Lmo3 and Sox6.
In the case of NSCs it has been reported that elongation of the G1 phase is observed with the start of neurogenesis (23). In order to study if spontaneous differentiation of mNSCs under normoxia is accompanied by changes in the cell cycle, the cell cycle pattern was studied by means of flow cytometry (Fig. 1E). We observed a significantly higher proportion of mNSCs in G0-G1 phase at the expense of cells in S phase in normoxic conditions if compared to those grown under physioxia. The fraction of cells residing in the G2-M phase remained at a similar level. Consistently, microarray analysis of genes attributed to cell cycle regulation (Fig. 1F) revealed that physioxia significantly alters some cell cycle regulators such as Cdk4, E2f1 as well as Mcm2, 3 and 4 in mNSC. Since P21Cip1, a cell cycle arrest marker, was upregulated we investigated other senescence/quiescence markers (Fig. 1G) (24). Minor changes could be observed regarding a slight upregulation Glb1 (Galactosidase), Frap1 (mTor) and Cdkn2a (P16) which interacts with Cdk4.
Together, these results show that physioxia stimulates proliferation of mNSCs, while sustaining their stemness by inhibiting spontaneous differentiation into the neuronal and glial fate and maintaining the cell population in the S-phase.
Chronic physioxia regulates Notch signalling in mNSCs
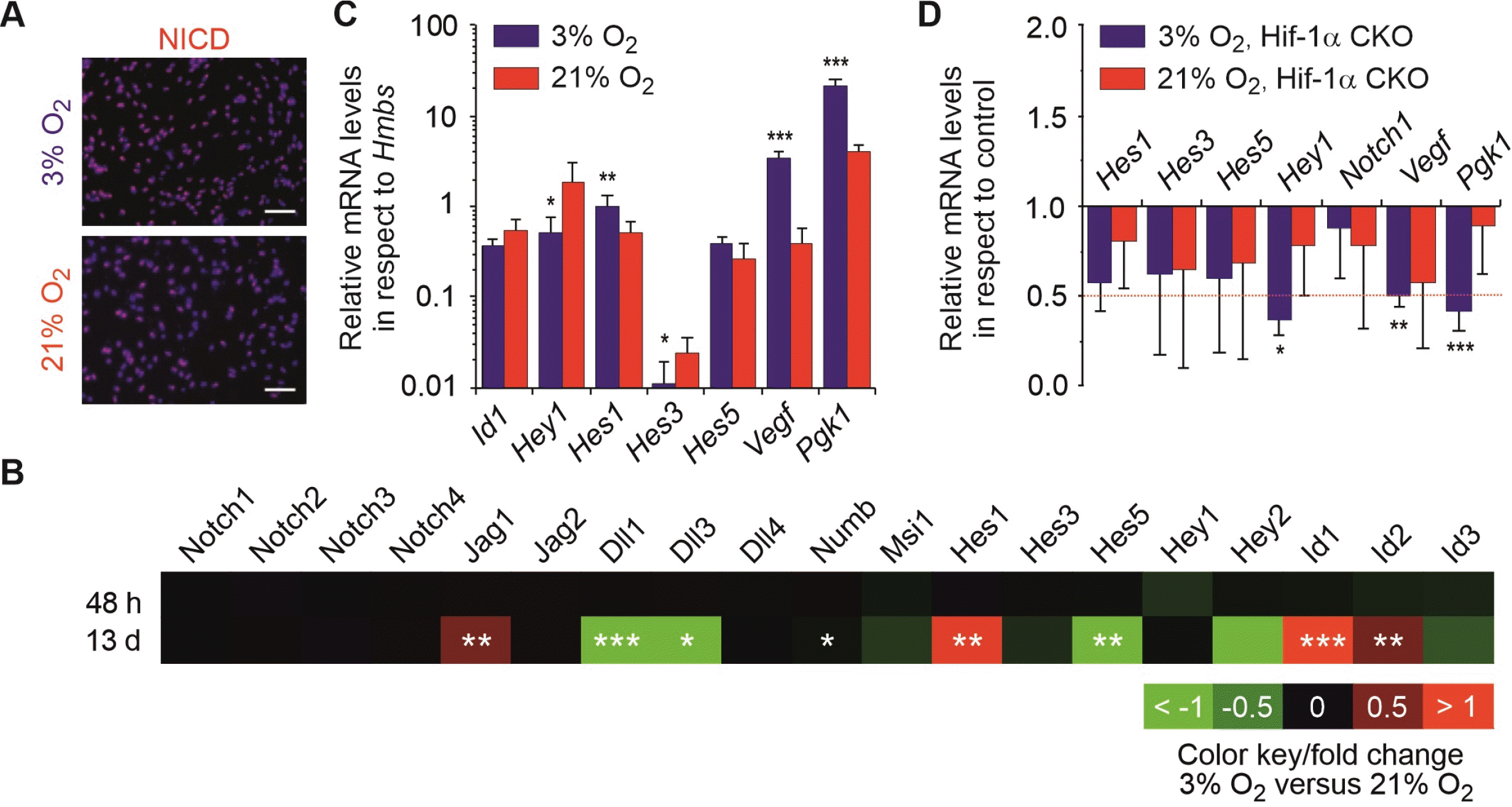
We next studied the Notch pathway by analysing the expression of members of the Notch signalling pathway using microarray analysis and immunocytochemistry. The latter revealed that NICD is present in both oxygen conditions indicating that the Notch pathway is active in mNSCs (Fig. 2A). To further assess the activity of Notch signalling, we measured Notch-related gene expression by microarray data analyses and qRT-PCR. As shown in Fig. 2B, microarray analysis of the Notch signalling pathway revealed that short-term culture of mNSCs in physioxia (48 h) led only to subtle changes, while a chronic exposure to physioxia (13 d) caused significant changes in the gene expression levels. The Notch target gene Hes1, as well as its interaction factors Id1, Id2 and also Jag1 are markedly upregulated. Notch ligands Dll1 and 3 were found to be downregulated alongside the Notch target gene Hes5. These data could be further validated by qRT-PCR showing increased levels of Notch target gene Hes1 in mNSCs upon physioxia, while the level of Hey1 was additionally decreased (Fig. 2C). Only a negligible expression of Hes3 with a significant decrease in physioxia could be found. The relative expression of other validated Notch target genes such as Hes5 and Id1 did not differ across the two oxygen conditions. Together our data indicate a shift of Notch signalling from Hes5 towards Hes1 in mNSC cultivated under chronic physioxia.
To analyse a potential crosstalk between the Hif-1α and Notch pathways, we investigated the transcription of validated target genes of both pathways in mNSCs with Hif-1α knock-out. Notably, all significant changes in Hif-1α CKO condition compared to controls were only observed in physioxia, which is in good agreement with the very low expression of Hif-1α under normoxic conditions leading to a functional knock-down also in wild-type cells (3). In Hif-1α CKO cells, the Hif-1α target gene levels of Vegf and Pgk1 were found to be downregulated under physioxia while from the investigated Notch targets, only Hey1 was down-regulated in Hif-1α CKO compared to controls (Fig. 2D).
Our results show that Notch signalling is active in mNSCs because Notch as well as its target genes were expressed in both oxygen conditions; however, physioxia caused significant changes in Notch ligand and target gene expre-ssion. We did not find any evidence for a regulation of Notch target genes by Hif-1α signalling, except for Hey1.
Notch enhances stem cell maintenance under normoxic conditions but has no influence on cell cycle distribution
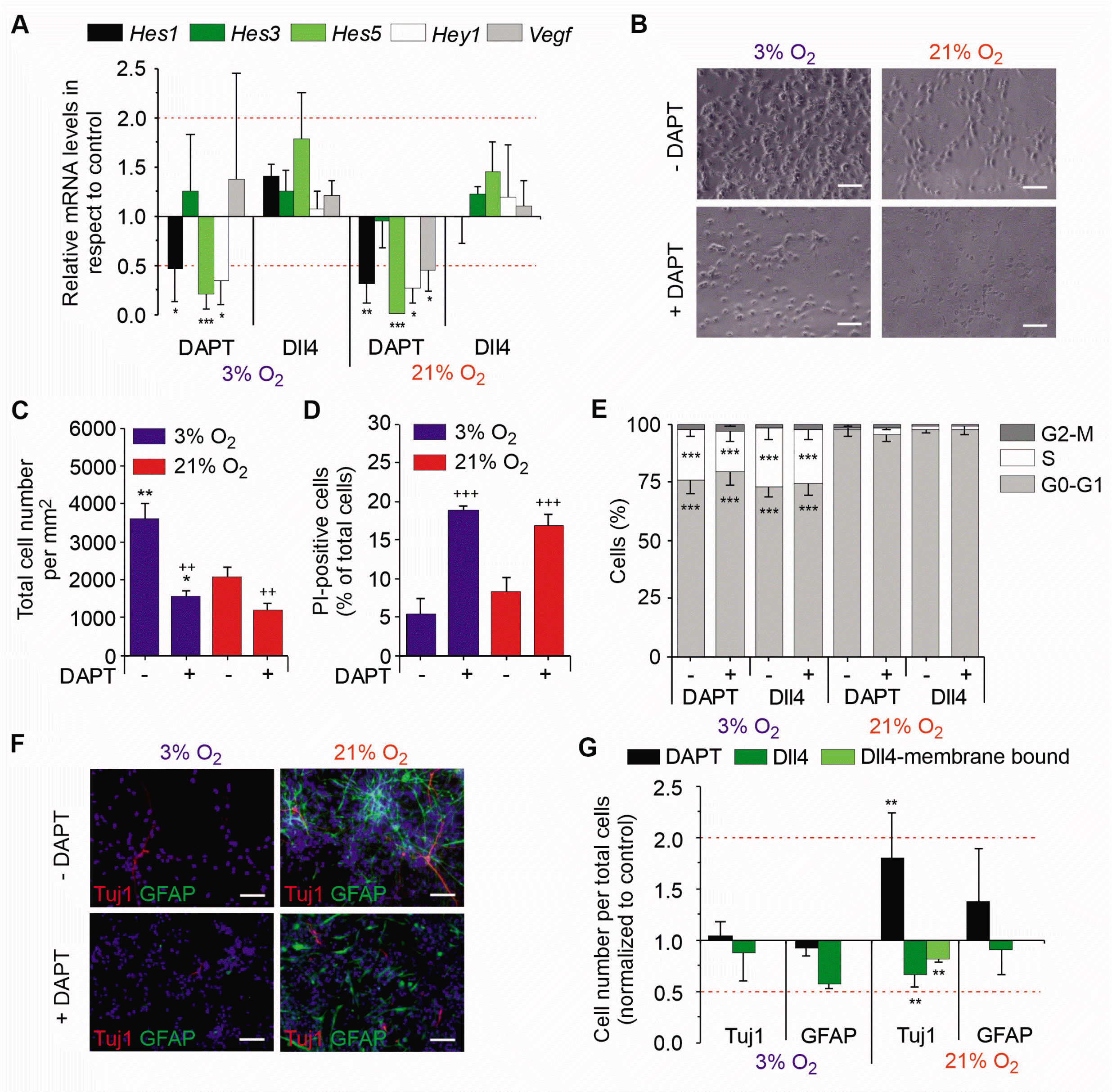
To substantiate the biological role of Notch in mNSC maintenance, mNSCs were cultured either in the presence of the γ-secretase inhibitor DAPT, considered to be a specific Notch inhibitor (25) or by addition of soluble Dll4, a Notch ligand promoting its activation (14). The effects of both strategies were validated by Notch target gene analyses (Fig. 3A): Addition of DAPT to mNSC cultures resulted in a downregulation of most target genes in both oxygen conditions, while stimulation with Dll4 did not change Notch target gene mRNA levels suggesting that the already active Notch pathways is not further stimulated by Dll4.
To study whether the Notch pathway, which is affected in mNSC under chronic physioxia, influences cell proliferation and cell death, mNSCs treated with DAPT were initially analysed for their cell count (Fig. 3B and 3C). Notch inhibition by DAPT treatment reduced cell counts of mNSCs by approximately 50% in both oxygen condi-tions. Consistently, DAPT treatment resulted in a significant increase of cell death compared to control with no significant differences between the two oxygen tensions (Fig. 3D). We then tested the effects of Notch signalling on cell cycle regulation by treating freshly isolated mNSCs with DAPT or Dll4 followed by flow cytometry analysis (Fig. 3E). While normoxia was found to enhance the cell population of G0-G1 phase at the expense of S as displayed above in Fig. 1A, inhibition or stimulation of Notch activity under either oxygen condition did not result in any change of the cell cycle phase distribution. Together, Notch signalling beneficially influences mNSC survival independent of oxygen effects, but does not mediate oxygen effects on cell proliferation or cell cycle kinetics.
To test the effects of Notch activity on stem cell maintenance of mNSCs, cells treated with DAPT (Fig. 3F) or Dll4 were analysed for their spontaneous differentiation using the markers Tuj1 and GFAP. In addition to soluble Dll4, we used an established murine cell system (Sc9-19) expressing high levels of Dll4 (21) to confirm the data on Dll4 treatment using membrane-bound Dll4. Notch inhibition by DAPT led to no changes of Tuj1+ cells under physioxia, while under normoxia a 2-fold increase of neurons was observed compared to control conditions. The activation of Notch by soluble and membrane-bound Dll4 did not alter Tuj1+ cell counts in physioxia, while we observed a decrease of neurons in normoxia. Inhibition or stimulation of Notch signalling in mNSCs had no effects on gliogenesis as significant changes in GFAP+ cell numbers were not observed (Fig. 3G). Together, our results show that Notch activity mediates stem cell maintenance exclusively under normoxic conditions, but has no influence on NSC proliferation and cell cycle kinetics.
Discussion
Physiological oxygen tension, also called physiological hypoxia or physioxia, varies among tissues and developmental stages, and is beneficial for proliferation and maintenance of several stem cell types. Here we show that the culture of murine fetal mNSCs under physiological oxygen levels of 3% sustains their stemness by promoting proliferation and suppressing spontaneous differentiation. Physioxia was found to regulate Notch signalling, which also contributes to mNSC proliferation. Since previous reports studying Notch function in neural stem or progenitor cell types were conducted in various cell types but mNSC, our data show for the first time that Notch signalling is not involved in physioxia mediated stem cell maintenance in vitro but acts as an alternative pathway for stem cell maintenance in mNSC cultured under chronic normoxia.
To address the impact of Notch in oxygen dependent stem cell maintenance, members of the Notch signalling pathway were analysed by microarray. While a short-term culture of mNSCs in physioxia lead to minor changes in expression levels, long-term culture of 13 d resulted in an upregulation of Hes1. Since Notch1, a classical activator of Hes1, as well as Notch2, 3 and 4 are not affected, the elevated level of Hes1 is likely not to be dependent on the classical notch signalling (26). Additionally, the level of the Notch ligand Jag1 was enhanced, while levels of classical Notch activators Dll1 and 3 were downregulated. Parti-cularly Jag1 is – at least in the cortical area – known to be involved in maintaining stem cells (27). Downregula-tion of Dll1 and 3 might further promote stemness as it has been reported that Dll1 inhibition caused by Hes1 oscillations sustains neural stem cell maintenance (28). Contributing to this, an overexpression of Hes1 was also found to inhibit fate decision in neurons and oligodendrocytes (13). Vice versa, decreased Hes1 expression together with an inhibition of Hes1 oscillations causes increased transcription of Hes3, Hes5 and Dll1 which inversely mimics our data (12). Besides Hes1, the Notch signalling inhibitors Id1 and 2, which are both able to regulate stem cell maintenance through binding bHLH proteins like Hes1 to 5, were increased under physioxia (29). Ids are assumed to uncouple Hes1 expression from classical Notch signalling which might explain unaffected levels of Notch ligands 1∼4 (30). Together with dampened levels of Hes5, Id3 and Hey2 these data suggest a cell-specific network of transcription factors and inhibitors working in synergy with hypoxic stimulation which interestingly reflects very early phase of hippocampal development in vivo (31). Hey2, for example, has been found to bind Hif-1β in the cardiovascular system, thereby preventing the DNA binding of the Hif-α/β heterodimer and thus transcription (32).
The addition of DAPT, an inhibitor of NICD release as part of the classical Notch signalling, efficiently inhibited Notch target gene expression of Hes1, 5 and Hey1 in mNSC under both oxygen conditions, while treatment with Dll4 resulted in no relevant upregulation, indicating that the Notch signalling pathway cannot be further activated in mNSCs, at least not by Dll4 stimulation. Very low Hes3 mRNA levels were found in mNSCs under both oxygen conditions – most likely due to the medium containing Egf which is known to downregulate Hes3 – without any evidence for its regulation by Notch (33). This is in accordance with our microarray results and it has also been reported that NICD does not bind to the Hes3 promoter, indicating that Hes3 is not regulated by Notch (34). In agreement, expression of Hes3 mRNA levels in human ES cell derived NSCs could also not be detected (25). Although oxygen-mediated increase of mRNA level of Hes1 does not depend on classical Notch signalling, the reduced mRNA level of Hes1 induced by DAPT together with detectable NICD proved that classical Notch signalling is still active under both oxygen conditions.
To study whether Notch influences spontaneous differentiation, mNSCs were cultured again in the presence of Dll4, which is known to result in Notch stimulation through interacting with hypoxia factors (35) and DAPT, a well-known inhibitor for Notch signalling. In physioxia, Notch inhibition had no effects on spontaneous differentiation, whereas in normoxia an increase of differentiated neurons and astrocytes could be observed. Vice versa, stimulation of Notch resulted in the reduction of neuronal differentiation in normoxia. Notch signalling, hence, does not promote stem cell maintenance in physioxia, but has an inhibitory role in neurogenesis in nor-moxia. Consistent to these data, it has been reported that Notch reduction enhances neuronal differentiation in normoxia, while its activation could abolish this effect and increase the amount of Nestin+ cells (36, 37). Borghese et al. (25) observed a five-fold increased neuronal differentiation upon DAPT treatment in in vitro culture of long-term human NSCs derived from ES cells as well as in vivo after transplantation into hippocampal slice cultures. In addition, the impact of Notch on NSC maintenance was found to vary depending on their developmental stage, their origin and their environment (38). It has been shown that a combination of diminished levels of Hes1, Hes3 and Hes5 is also able to change fate decisions in cortical NSCs cultured under normoxia (12).
A direct interaction of Hif-1α with NICD has been previously reported for other neuroprecursor cell types (15, 16). Indeed, we could find an upregulation of Notch target gene Hes1 under physioxia, although likely independent off classical Notch signalling. Moreover, Notch was shown to have effects on stem cell maintenance albeit only under normoxia where Notch might replace Hif-1α function. This view is supported by the facts that substitution of Vegf affects mNSC proliferation only in the Hif-1α knock-out condition in which Notch presumably takes over Hif-1α actions and that expression of the Hif-1α target gene Vegf is inhibited by DAPT only in normoxia. Kaufmann and colleagues reported that proteolytic γ-secretase function is needed for proper Hif activation via the amyloid precursor protein (APP)/APP intracellular domain (AICD) cleavage cascade (39). Since in fetal cells/tissue the γ-secretase inhibitor DAPT (as used in the present study) mimics the Notch knock-out phenotype, DAPT effects are assumed to be mediated mainly by the Notch pathway in fetal cells, but direct effects of DAPT on Hif-1α activity are not fully excluded.
In our hands, only one of the Notch target genes (Hey1) showed a downregulation upon Notch as well as Hif-1α inhibition suggesting a possible converging mechanism. This finding is in agreement with research about Droso-phila blood cells and cancer cells showing interactions of Hif-1α and Hif-1α orthologs with Notch signalling via a hypoxia response element within the Hey1 gene (40). Although the overall effect of Hif-1α cKO on Notch signalling was marginal, Hey1 is an important physiological factor in cortical and hippocampal brain development and therefore Hif-1α, at least in part, could also influence midbrain development (31).
In addition to the stem cell fate decision, cell cycle analysis revealed that normoxia decreases the percentage of mNSCs in S-phase compared to physioxia. The increase of the cell population in G0-G1 phase caused by normoxia might thus promotes spontaneous differentiation of mNSCs. As Notch activation or inhibition did not result in changes of cell cycle distribution, and stimulation of mNSCs with the Notch ligand Dll4 could not reverse the cell cycle arrest inflicted by normoxia, we suggest that changes in mNSC cell cycle distribution are Notch independent. In contrast, Borghese et al. (25) demonstrated that DAPT treatment for 3 d increases the cell population in G0-G1 phase from around 58% to 65% in normoxic embryonic stem cell-derived NSCs. One reason for this discrepancy might be the fact that in normoxia with active Notch almost all mNSCs (approximately 95%) were already in G0-G1 phase and a further increase was not observed.
It has been reported, that lengthening of the G1 phase acts as switch from proliferation to neurogenesis and thus differentiation and that the lengthening of the G0-G1 phase alone can lead to differentiation of neuroepithelial cells (23). Apart from its correlation to differentiation, the cell cycle length of murine neurons of the cerebral neocortex has been reported to double during embryonic day E11 and E16 and this lengthening was found to be caused solely by variations of the G0-G1 phase (41). These findings together with our results are an indication that the cell cycle length increases during embryonic development along with differentiation. These findings seem to contradict other observation where inhibition of cell cycle progression by G1 arrest under hypoxia was found (42). Although these studies were conducted under severe hypoxia (≤0.5% O2), a condition likely to inflict oxygen deprivation and different to the physiological level of hypoxia (3%) used here, we found also signs of cell cycle arrest as the level of the CDK inhibitor P21cip1 was upregulated, while Cdk4 was downregulated. Our analysis of quiescence/senescence markers further revealed minor changes regarding endogenous β-galactosidase, mTor pathway and Cdkn2a (P16), which directly interacts with Cdk4, indicating that at least a small proportion of mNSC developing a quiescence or senescence like status. However, the promotion of cell cycle progression was confirmed by regulation of transcription factors E2f1 and 4, which activate and repress G1-S phase transition, respectively (43). Indeed, levels of E2f1 were increased along decreased expression of E2f4. Together the changes in key regulators of the cell cycle inflicted by hypoxia could support mNSCs maintenance by promoting cell cycle progression as well as preventing exhaustive proliferation through expressing senescence-like markers and thus keeping a healthy balance of mNSCs (44).
In conclusion, we show that oxygen tension critically regulates stem cell performance of midbrain NSCs in vitro as physioxia regulates cell proliferation, stemness and the cell cycle. Notch target gene Hes1 was specifically upregulated in physioxia and Notch signalling was found to sustain mNSC maintenance under normoxia, indicating that Notch could act as Hif-1α substitute in normoxic condi-tions. Our new findings highlight the importance of appropriate culture conditions for mNSCs maintenance: the level of hypoxia used in the present studies does not only resemble their in vivo environment more closely but also improves their potential performance for regenerative therapies.
 XML Download
XML Download