

 PDF
PDF Citation
Citation Print
Print


Introduction
Multipotent mesenchymal stromal cells (MSCs) are obtained from various adult tissues, such as bone marrow (BM), adipose tissue, umbilical cord blood, Wharton’s jelly, dental pulp, and skin (1). MSCs have been investigated as an attractive option for regenerative medicine and immune-mediated disorders due to their self-renewal ability, multilineage differentiation, and anti-inflammatory and immunomodulatory functions, which involve both paracrine and cell-to-cell contact mechanisms (2). Several studies have shown that the potent immunomodulatory properties of MSCs are due to the production of immune-modulating factors, such as indoleamine 2,3-dioxygenase (IDO), heme oxygenase-1 (HO-1), transforming growth factor-beta (TGF-β), tumor necrosis factor-alpha (TNF-α)-stimulated gene protein-6 (TSG-6), cyclooxyge-nase-2 (COX-2), and prostaglandin E2 (PGE2), which target various components of the immune system (3). These properties support the use of MSCs for cellular therapeutics, and accordingly, various tissue regeneration treatments with MSCs have been evaluated in clinical trials. Nonetheless, their clinical applications have not been satisfactory (4) due to poor engraftment, in vitro senescence, functional quiescence after transplantation in vivo, and the heterogeneity of MSC properties, which can be attributed to an incomplete understanding of the mechanisms regulating MSCs. Elucidating the anatomical and functional features of MSC niches in their native tissues and the interactions of MSCs with the extracellular matrix (ECM) and surrounding cells is crucial to understanding the behavior of MSCs and to harnessing this knowledge for clinical applications.
The MSC niche has been defined as all of the elements immediately surrounding the stem cells when they are in their naive state (5). These elements include the non-stem cells in direct contact with the MSCs as well as the ECM and physical cues found in that locale (6, 7). As MSCs move away from this nurturing niche, they may encounter different environments, such as fewer cell-cell interactions and more ECM interactions, which regulate MSCs differentially. Features of the surrounding environment of MSCs, such as the microarchitecture, substrate rigidity, or oxygen level, affect and regulate MSC function and property. Maintaining the in vivo niche or enforcing this condition is considered to provide greater therapeutic potential and efficacy. Therefore, to better mimic in vivo conditions, nature-inspired platforms for cell culture have come to the fore (8). The extracellular microenvironment that controls the ability of MSCs includes its biological, chemical, and physical aspects. Although many studies have focused mainly on biological and chemical cues, the physical cues of the extracellular microenvironment are receiving increasing attention. The physical properties of the extracellular microenvironment include biomechanical forces and the intrinsic ECM.
Therefore, in this review, we summarize the application of nature-inspired platforms that mimic the biomecha-nical forces and the biophysical cues in the MSC niche. Additionally, the effects of the interaction between MSCs and their microenvironment on MSC immunomodulatory potential will be discussed.
Immunomodulatory Function of MSCs by Mechanical Force: Sensing and Reaction
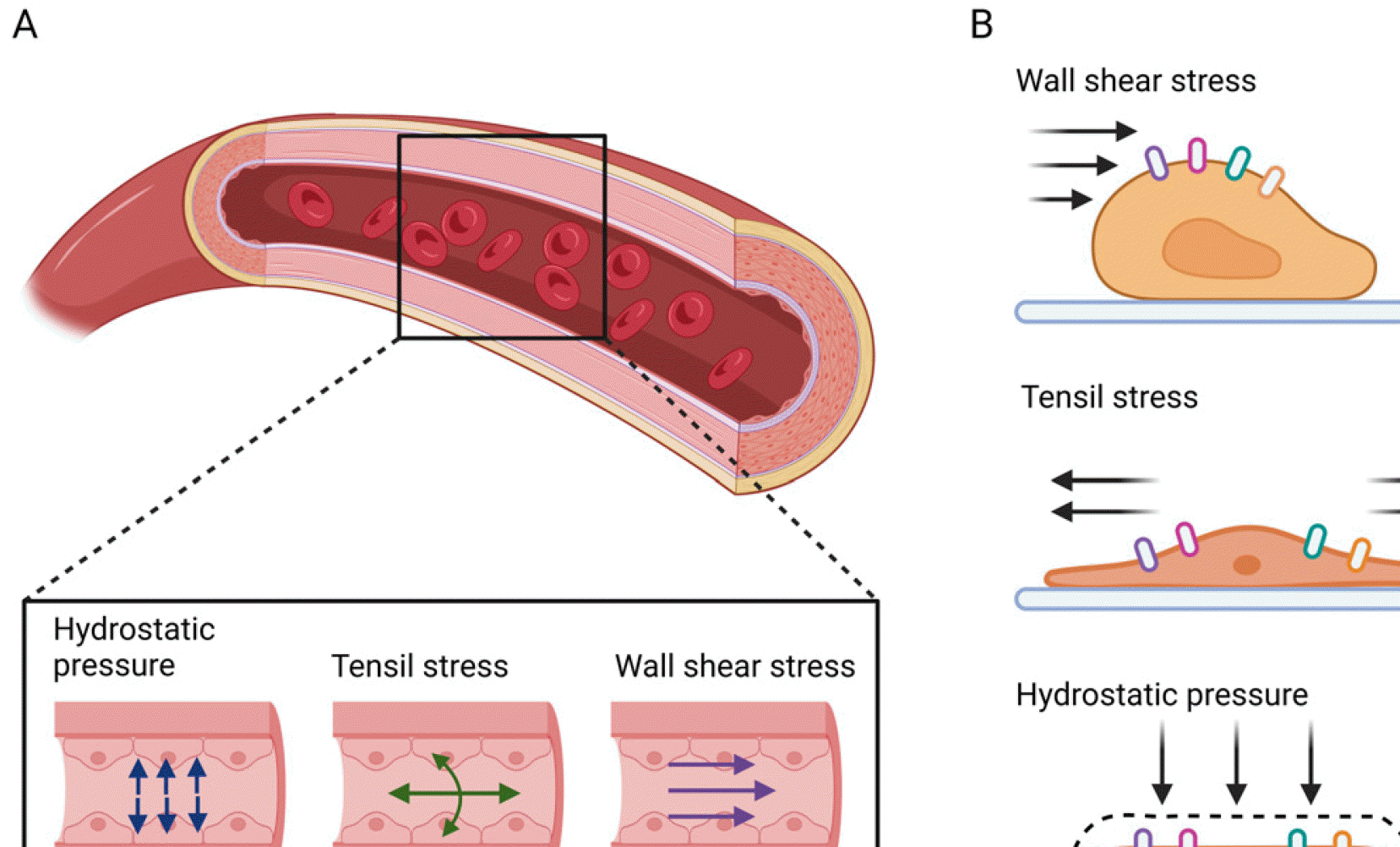
MSCs have served as an archetypal model for understanding the properties of mechanical forces on cellular potential largely because of their ubiquitous nature, ease of ex vivo culture, and multipotentiality as bone progenitors (9). For example, BM-derived MSCs experience many mechanical forces, such as shear stress, hydrostatic pressure, and tensile forces, in their MSC niche (10, 11). These biomechanical forces can be transduced into internal biochemical signals in MSCs through their mechanosensors or responsive microdomains (12, 13). In the following section, we describe the immunomodulatory functions of MSCs in terms of their response to biomechanical forces, including shear stress, tensile stress, and hydrostatic pressure (Fig. 1).
Shear stress, tensile forces, and hydrostatic pressure as biomechanical forces
As a force tending to cause deformation of an object by slippage along a plane or planes parallel to the imposed stress, shear stress is known to exist in the blood vessel, lymphatic vessel, or lacunar-canalicular network of bone due to the fluid movement (Fig. 1) (14). These fluid flow patterns are not uniform in the biological system because shear stress depends on the velocity gradient between these layers and the viscosity of the liquid (15). Diaz et al. (9) demonstrated that vascular lumen mimicking fluid frictional forces (15 dyne/cm2) elevated antioxidant and anti-inflammatory mediators from BM-derived MSCs, suggesting that biomechanical forces can improve the immunomodulatory function of MSCs by providing critical cues to MSCs residing at the vascular interface. Shear stress facilitates the opening of the stretch-activated calcium ion channels on the surface of MSCs, inducing an influx of intracellular calcium ions and activation of the focal adhesion kinase (FAK)-Akt signaling axis, inclu-ding upregulation of COX-2, HO-1, and PGE2 and downregulation of TNF-α secretion (16). This signaling elevates the immunomodulatory function of MSCs to suppress an inflammatory response (16). Santos et al. (17) demonstrated the increased secretion of interleukin (IL)-6 by MSC spheroids exposed to spinner vessels equipped with ball impellers at 80 rpm, whereas a spinner flask at 40∼50 rpm elevated the immunomodulatory properties of MSCs, which suppressed T cell proliferation (18). Further, artificial lymph node mimicking reactor enhanced anti-inflammatory cytokines (IL-1 and IL-12) and inhibited pro-inflammatory cytokines (TNF-α and IFN-γ) in rat MSCs (19). These findings indicate that an appropriate range of shear stress can trigger a cascade of inflammatory mediators critical in MSC immunomodulatory function.
As another biomechanical force, tensile stress is known to affect cell biology and gap junction intracellular communication (20). The blood vessel wall structure is built to withstand and propagate the forces applied by blood flow, pressure, and the surrounding tissues (Fig. 1A). Blood pressure measures the cyclic tensile forces acting radially and longitudinally on the vascular wall (21). Thus, vascular endothelial and smooth muscle cells are subject to tensile stress under pulsatile flow conditions (Fig. 1B). In addition, tensile forces are important physical cues that regulate MSCs within the stem cell niche (22). Intracellu-lar tensile forces resulting from cytoskeletal reorganization play a critical role in regulating morphogenesis during development and enhancing the immunomodulatory properties of MSCs. When MSCs were cultivated on a custom-made polydimethylsiloxane (PDMS) membrane of 150 μm in thickness that was deflected to obtain a circumferential stretch of about 20% at a frequency of 0.2 Hz, the IL-6 and TNF-α levels were decreased, thus suggesting that the secretome from MSCs had changed (23). This secretion profile increased the anti-inflammatory and immunosuppressive outcomes of MSCs (23). By contrast, 10% or 12% uniaxial tensile strain on BM-derived MSCs increased IL-6 or IL-8 secretion as inflammatory cytokines, initiating osteogenic differentiation (24), which suggested that biomimetic forces modulate the autocrine signals of MSCs, such as IL-6 or IL-8. In summary, tensile stress differentially affects the function of MSCs dependent on the magnitude of the force or type of material to modulate MSC function.
Hydrostatic pressure is an important cellular cue (Fig. 1A) that regulates cell behaviors, such as differentiation, migration, apoptosis, and proliferation in vivo and in vitro (25, 26). The physiological hydrostatic pressure differs substantially by cell and tissue type. In some cases, changes in the hydrostatic pressure can be correlated with a pathological condition. For example, high hydrostatic pressure can induce vesicoureteral reflux and upper urinary tract deterioration (27). On the microscale, hydrostatic pressure can affect the cytoskeleton of cells. At sufficient amplitudes, hydrostatic pressure has been shown to induce the disassembly of the microtubule-based mitotic apparatus, resulting in cell-cycle arrest (28). Furthermore, hydrostatic pressure activates the noncanonical Hippo-YAP (Yes-associated protein)/TAZ (Transcriptional coactivator with PDZ-binding motif) pathway, enhancing clathrin-dependent endocytosis by regulating the cytoskeleton of cells (29). Tra-nscriptional factors YAP and TAZ have been recognized as key mechanotransducers that sense mechanical stimuli and relay the signals to regulate the transcriptional programs for cell proliferation, differentiation, and transfor-mation.
Sugimoto et al. (30) demonstrated that hydrostatic pressure induced Piezo-type mechanosensitive ion channel component 1 (Piezo1) channel activation as a mechanosensor, thereby promoting osteogenic differentiation through extracellular-regulated kinase (ERK) and p38 signaling in MSCs. Moreover, MSCs increased the expression of COX-2 when MSCs were cultivated in a custom pressure bioreactor at 10∼300 kPa magnitude, 0.5∼2 Hz, and up to 4 h of stimulation (31). Recently, bone mimicking mechanical forces drove physiological level of TNF-α secretion from mouse MSCs which was helpful to maintain stem cell homeostasis (32). Overall, these outcomes indi-cated that biomechanical forces regulate the differentiation and immunomodulatory properties of MSCs by stimulating mechanosensors on their membrane (Fig. 1B). A literature overview of the control of MSC immunomodulation by biomechanical forces is summarized in Table 1.
Biomimetic ECM
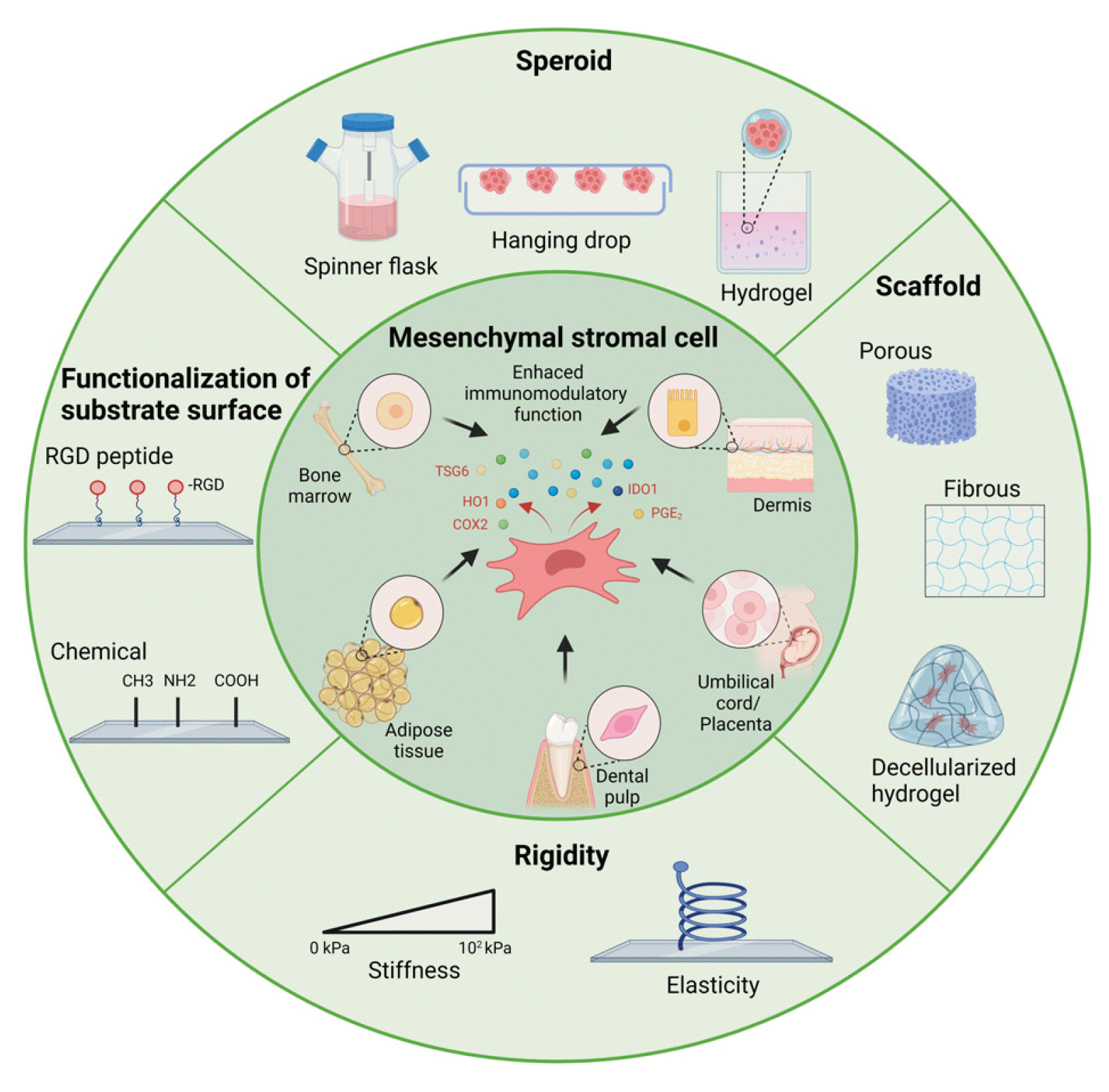
The MSC niche is a specialized microenvironment composed of MSCs, differentiated progenitor cells, non-stem supporting cells, and a non-cell part, the ECM (33). The ECM is a complex three-dimensional (3D) network of interlaced fibronectin, collagen, proteoglycans, multiple matrix protein macromolecules, growth factors, and bioactive factors that provide physical support for cells and modulate cell functional activity, proliferation, adhesion, and migration (34). Thus, various 3D microstructures have been designed to mimic the ECM of the native MSC niche. Multiple lines of evidence indicate that 3D aggrega-tion of MSCs enhances their immunomodulatory proper-ties. The formation of cell spheroids enhances cell-cell contact, and the mechanical properties of the extracellu-lar microenvironment, such as matrix stiffness and elasticity, also affect MSC biology. The following section discusses the immunomodulatory properties of MSCs as affected by the ECM, such as stiffness and substrate surface functionalization, and various 3D culture systems using a spheroid culture or polymer scaffold (Fig. 2).
Spheroid as a 3D culture
Cell attachment to the ECM and the bidirectional signal transduction between the ECM and cells is mediated by a subset of integrins that specifically recognize the prevalently surface-displayed and evolutionary conserved arginine-glycine-aspartate (RGD) motif, which is present in fibronectin, fibrinogen, and various other proteins of the ECM. Integrin-mediated adhesion leads to the formation of loose masses composed of several cells, which increases intercellular communication and cadherin expre-ssion in these cells. Gradual accumulation on the surface of MSCs leads to increased cell density and multicellular spheroid formation (8).
Spheroid formation technologies can be divided into static models, including hanging drop, hydrogel, and concave micro-well aggregation, and dynamic models, including suspension cultures and spinner flask bioreactors (8). In several reports, the 3D spheroid culture enhanced the secretion of several immunomodulatory factors regardless of the spheroid formation technique. Bartosh et al. (35) and others showed that 3D spheroids formed using the hanging drop technique enhanced the protein expression of tumor necrosis factor-(TNF)-stimulated gene-6 (TSG-6) and stanniocalcin-1 (STC-1), which have anti-inflammatory effects (35-37). Moreover, Camões et al. (38) showed that the systemic levels of the pro-inflammatory cytokines IL-6 and TNF-α in the exosomes of spheroids formed using a spinner flask culture system were reduced in the late stages of an in vivo wound healing model.
Scaffolds, substrate rigidity, and their functionalization as a biophysical cue
Scaffolds mimicking the properties of the MSC niches can provide a 3D environment for transplanted cells to support cell survival, adhesion, growth, and differentiation and enhance paracrine function, including immunomodulatory potential. A scaffold based on biodegradable biomaterials is preferred, except for bone and dental implants, because of the elimination of triggering a host immune response or toxic response. For this purpose, various techniques considering materials and shapes that can provide excellent biocompatibility and biomechanical properties have been considered (39). MSCs are typically delivered to a graft site using a decellularized ECM scaffold. Li et al. (40) showed that the MSCs cultured on a porcine decellularized liver scaffold system secreted more PGE2 and less interferon-amma (IFN-γ) than traditional monolayer culture. In terms of biomaterials, highly tunable and biodegradable natural polymers, such as fibrin, collagen, and hyaluronic acid, possess biochemical cues or trophic factors that promote the recruitment of stem cells, suppress inflammation, and enhance tissue repair (41). Silk fibroin, a natural polymer derived from Bombyx mori, is a biocompatible material that can be processed into various forms, such as fibers, gels, films, and porous 3D microstructures. According to Kim et al. (42, 43), a meshed scaffold comprising silk fibroin nanofibers stimulated the expression of immune modulators, such as indoleamine-pyrrole 2,3-dioxygenase (IDO-1), COX-2, and PGE2, and promoted the survival rate of the polymicrobial sepsis-induced mouse model. Synthetic polymers, such as polycaprolactone (PCL), poly (L-lactic-co-glycolic acid) (PLLA), and poly (lactic-co-glycolic acid) (PLGA), provide a 3D environment in the form of fibrous or porous sponges, plates, or membranes, and change the function of MSCs (44-46). Wan et al. (47) reported that human adipose-derived MSCs enhanced the expression of COX-2 and TSG-6 in aligned PLLA electrospun fibrous scaffolds compared to randomly fibrous scaffolds. In addition, Li et al. (48) demonstrated that a 3D porous scaffold using collagen, chitosan, and PLGA improved the stemness of MSCs compared to two-dimensional culture and upregulated the expression of IL1A, IL1B, IL-1 receptor antagonist (IL1RN), hepatocyte growth factor (HGF), and epidermal growth factor (EGF), which are immunomodulation-related genes.
As another physical cue, substrate stiffness affects not only the morphology, proliferation, and differentiation of MSCs but also their immunomodulatory function (48). However, there are conflicting reports associating stiffness with enhanced immunomodulatory function. Wong et al. (49) demonstrated that soft ECM (0.3∼2 kPa) maximized the ability of TNF-α-primed MSCs to produce paracrine factors compared to a stiffer ECM (100 kPa). By contrast, Darnell et al. (50) reported that increasing the hydrogel stiffness from 3 to 18 kPa stimulated the expression of inflammatory modulators, such as IDO-1 and COX-2, in D1 mouse MSCs. Despite the contradictory findings, such studies indicate that the optimal range of stiffness of biomaterials can be a promising tool for enhancing the immunomodulatory properties of MSCs (51). Further studies are needed to suggest a range of strengths or structures that enhance immunomodulation.
Along with spheroids, scaffolds, differences in matrix stiffness, and different types of biomaterials, the functionalization of the substrate surface, such as surface chemistry and biomolecular properties, has been reported to provide a synergistic effect in improving the immunomodulatory function of MSCs (3). Roger et al. (52) demonstrated increased secretion of PGE2 and interleukin-1 receptor antagonist (IL1RA, a protein encoded by the IL1RN gene) by MSCs cultured on grid-like thermoplastic polyurethane (TPU) plates compared to unstructured TPU surfaces. In addition, an RGD peptide, one of the key peptide sequences found in ECM proteins (as mentioned above), can enhance MSC adhesion when immobilized to a substrate, ultimately increasing osteogenic differentiation. It has been reported that RGD peptides can alter macrophage behavior and function by mediating macrophage adhesion and alleviating macrophage inflammation in response to bioma-terials. Li et al. (53) applied a poly (dopamine) (DOP) coating to TiO2 nanotubes (T/DOP) to functionalize with IL-4 (T/DOP/IL-4) and an RGD peptide and then covered T/DOP/IL-4 with a carboxymethyl chitosan hydrogel layer (T/DOP/IL-4/CG-RGD) to control IL-4 release and RGD peptide immobilization. This T/DOP/IL-4/CG-RGD surface on macrophages not only induced the conversion of macrophages to the anti-inflammatory M2 but also increased the expression of IL-10 compared to T/DOP/CG (53). The immunomodulatory functions of MSCs according to biomimetic ECM types are summarized in Table 2.
Conclusions
In this review, various cell culture systems were proposed to enhance the immunomodulatory effects of MSCs. Enhancing the immunomodulatory capacity of MSCs may be critical in MSC-based cell therapy. However, several limitations will need to be overcome in order to realize the full clinical potential of MSCs. Despite various efforts to enhance the immunomodulatory ability of MSCs, there is no standardized cell culture system that embodies the microenvironment of MSCs. In addition, enhanced immu-nomodulation in the current system can be made through epigenetic regulation, which does not last long. Thus, it could cause a need for a stylized MSC culture system for use as an international standard. In addition, the characteristics of MSCs, which require applying different culture systems for patient-specific immune therapy, MSCs will require more stabilization. Furthermore, the secretion of anti-inflammatory cytokines and other factors may paradoxically cause pathological immune responses depending on the situation involved in disease progression. For these reasons, in some cases, there is still a need for more standardized monitoring data of MSC transplantation in various diseases to relieve concerns.
Although further study is needed to tackle these hurdles, MSCs have considerable potential as alternative treatments for various diseases. Therefore, a thorough understanding of the mechanisms of MSCs in a standardized cell culture platform considering the MSC niche will improve the safety, efficacy, and results of MSC-based therapy.
 XML Download
XML Download