

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Chronic obstructive pulmonary disease (COPD) is a respiratory disease with 10.3% global prevalence among people aged 30–79 years in 2019.12 It was the third most common cause of death in 2019 worldwide, and as one of the leading causes of global disease burden, it is a major public health concern.34 COPD is characterized by persistent respiratory symptoms and airflow obstruction caused by significant exposure to noxious gases or particles, including smoking.5 Smoking generates aberrant inflammation that injures the airways and alveoli, contributing to the occurrence of COPD, and is regarded as the most critical risk factor for COPD.6 However, the detailed molecular mechanisms in the pathogenesis of COPD have not been fully elucidated,7 leading to setbacks in development of better therapies for COPD patients.
Recent advances in genomics and proteomics have enabled screens for the components that play essential roles in COPD pathogenesis.8910 However, previous studies have limitations. Firstly, most studies have shown the functional roles of cytokines and immune cells in COPD, and few studies have explored the direct changes in epithelial cells.10 Considering that inhaled cigarette smoke has primary contact with the airway epithelial cells and that key histological findings in COPD, such as squamous cell metaplasia, and goblet cell hyperplasia, occur in epithelial cells, it is important to understand the pathological changes of epithelial cells in COPD. Secondly, smoking is one of the strongest factors causing COPD; however, most studies have focused on naïve COPD tissues and have yet to explore the differential response between COPD and non-COPD lungs to smoking exposure. To understand the link between smoking and COPD epithelial cells, we must understand the responses of the disease model to smoking exposure. Finally, high-throughput omics studies are vulnerable to false positive results since they produce numerous signals, especially in comparison to the number of samples. Therefore, feature selection algorithms need to be more resistant to noisy data in order to identify the key components in the pathogenesis of COPD.
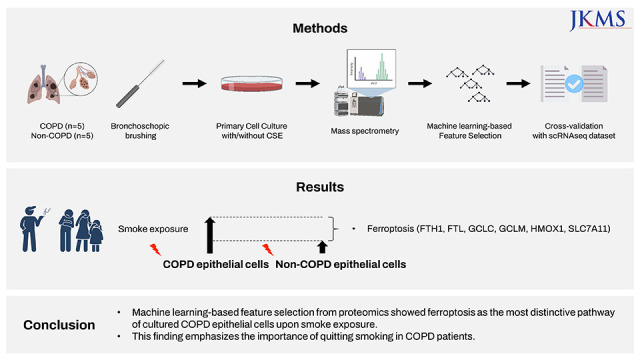
In this study, we performed mass spectrometry (MS) with machine learning (ML)-based pathway analysis to identify the most distinctive pathways in COPD epithelial cells exposed to cigarette smoke extract (CSE), compared to cells from patients without non-COPD that were also exposed. The pathways were also explored using publicly available COPD single-cell RNA sequencing data relating to cells from patients with COPD and healthy controls.
METHODS
Sample collection and preparation
COPD patients and non-COPD participants who underwent bronchoscopy evaluation for their clinical needs were enrolled with informed consent. During the examination, bronchial epithelial cells were collected by bronchoscopic brushing using the protected sheath on the distal small airways. The COPD group was defined as smokers with emphysema on lung computed tomography (CT) with post-bronchodilator (BD) forced expiratory volume in 1 second (FEV1) over post-BD forced vital capacity (FVC) < 70%. The non-COPD group was defined as non-smokers with no emphysema on CT, whose post-BD FEV1/FVC was > 80%. Specimens were obtained from the lobe with emphysema in patients with COPD, and from the right middle lobe in patients without COPD. The brushed cells were cultured in defined keratinocyte-SFM (GIBCO, Life Technologies, Grand Island, NY, USA) with 1% penicillin/streptomycin. Cells were grown at 37°C with 5% CO2, and the media were changed every other day. CSE was prepared from 20 commercial cigarettes (THIS; KT&G Corp., Daejeon, Korea) continuously bubbled in 60 mL of phosphate buffered saline using a bottle system connected to a vacuum machine (Gast Manufacturing Inc., Benton Harbor, MI, USA). To determine the optimal concentration of CSE, cultured bronchial epithelial cells were treated with 0, 0.5, 1, 2, or 4% CSE for 24 hours and then the cell viability was measured with an MTT assay. Based on these results, cells exposed to 2% CSE-containing culture medium were compared to those treated with control medium for 24 hours before harvest were used.
Mass spectrometry
Cultured primary cells, either treated or untreated with 2% CSE, were detached with TrypLE (Thermo Fisher Scientific, Waltham, MA, USA) and centrifuged at 300 g for 5 minutes. From each sample, cells were lysed using 100 µL of buffer (4% sodium dodecyl sulfate [SDS], 2 mM tris (2-carboxyethyl) phosphine in 100 mM Tris; pH 8.5) and incubated at 95°C for 20 minutes. Protein concentrations were measured using a reducing agent-compatible BCA assay (Thermo Fisher Scientific). After acetone precipitation of 100 µg of proteins, precipitated protein samples were digested using the filter-aided sample preparation procedure as previously described.11 Briefly, samples were dissolved with 2% SDS and 2 mM tris (2-carboxyethyl) phosphine in 100 mM Tris/HCl, pH 7.5, mixed with 0.3 mL of 8 M urea in 0.1 M Tris/HCl, pH 8.5, and loaded onto a 30 K Amicon filter (EMD Millipore, Billerica, MA, USA). The buffer was exchanged with urea solution by centrifugation. The reduced cysteines in the proteins were alkylated with iodoacetamide solution in the dark at room temperature for 30 minutes. An additional 40 mM of ammonium bicarbonate was added to exchange the urea solution. Finally, proteins were digested at 37°C overnight with trypsin at an enzyme-to-protein ratio of 1:100. After overnight incubation, the filtration unit was transferred to new collection tubes and centrifuged for 20 minutes. Peptides that were retained in the filtration units were eluted with 50 μL of 0.5 M NaCl to enhance the yield of digested protein. The resulting supernatants were acidified with 1% trifluoroacetic acid. Peptides were desalted and fractionated on homemade styrene divinylbenzene reversed-phase sulfonate-StageTips12 by basic reverse-phase using a stepwise gradient of acetonitrile (40%, 60%, and 80%) in 1% ammonium hydroxide.
Liquid chromatography-MS/MS analysis was performed using a Q Exactive HF-X Hybrid Quadrupole-Orbitrap mass spectrometer (Thermo Fisher Scientific) coupled to an Ultimate 3000 RSLC system (Dionex, Sunnyvale, CA, USA) via a nano-electrospray source as previously described with modifications.1213 Peptide samples were separated on a two-column system, consisting of a trap column and an analytic column (75 μm × 50 cm) with a 90 minutes gradient from 7% to 32% acetonitrile at 300 nL/min and analyzed via MS. The column temperature was maintained at 60°C using a column heater. Survey scans (300–1,650 m/z) were acquired with a resolution of 70,000 at m/z 200. The top-15 method was used to select precursor ions with an isolation window of 1.2 m/z. MS/MS spectra were acquired at an higher-energy collisional dissociation-normalized collision energy of 30 with a resolution of 17,500 at m/z 200. The maximum ion injection times for the full scan and MS/MS scans were 20 ms and 100 ms, respectively.
Mass spectra were processed using MaxQuant version 1.6.1.10.14 MS/MS spectra were searched against the Human Uniprot protein sequence database (December 2014, 88,657 entries) using the Andromeda search engine.15 Primary searches were performed using a 6-ppm precursor ion tolerance for total protein level analysis. MS/MS ion tolerance was set to 20 ppm. Cysteine carbamidomethylation was used as a fixed modification. N-acetylation of proteins and oxidation of methionine were set as the variable modifications. Enzyme specificity was set to full tryptic digestion. Peptides with a minimum length of six amino acids and up to two missed cleavages were considered. The required false discovery rate was set to 1% at the peptide, protein, and modification levels. We also enabled the “Match between Runs” option on the MaxQuant platform to maximize the number of quantification events across samples. For label-free quantification (LFQ), the intensity-based MaxLFQ algorithms16 were used as a part of the MaxQuant platform.
Differentially expressed peptide (DEP) analysis
MaxQuant data were processed and analyzed via the R package ‘DEP’ version 1.12.0, using default parameters. Briefly, LFQ intensities were normalized using variance-stabilizing transformation and missing data were imputed using random draws from a Gaussian distribution centered around a minimal value with a q-value of 0.05. Differential enrichment analysis was performed using linear and empirical Bayesian statistics. Significant proteins were denoted based on the following criteria: P value < 0.05 and log fold change > 1.5.
Feature selection by machine learning method
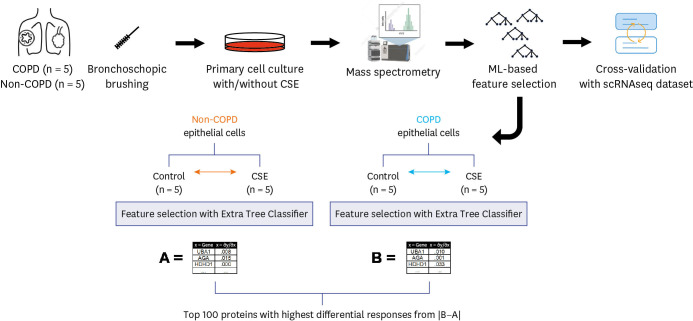
Next, we used an ML-based approach to select features from the raw LFQ intensity data to investigate the effects of various environmental factors on cells from COPD patients. To differentiate proteins that are enriched when exposed to CSE, we used Extra Trees Classifier, a variation of the random forest.
First, we trained Extra Trees Classifiers with raw LFQ data to distinguish the features of CSE-treated samples in the non-COPD group from the matched non-COPD samples. We interpreted the feature importance of the trained model, denoted as set A (Fig. 1), as the featured proteomic response to CSE. Next, the proteomic response of the COPD group was computed using Extra Trees Classifier trained on CSE-treated and non-CSE treated samples in the COPD group, denoted as set B (Fig. 1). Then, we subtracted the proteomic response of the non-COPD group from the COPD group and selected the top 100 proteins with the highest differential response. Pathway analysis was performed using Enrichr17 with the Gene Ontology gene set. All pathways were sorted on the adjusted P value. Z-score and combined score, log(p)·z, are adopted from Enrichr.
Validation with single cell RNA sequencing data
Data from the Gene Expression Omnibus accession number GSE136831 representing single-cell RNA sequencing data, including data obtained from 18 lungs from patients with COPD and from 28 control donor lungs, were re-analyzed.18 Only cells annotated in metadata were included, and cell types with at least 100 cells in both COPD and control groups were used for further analysis.
Differentially expressed genes (DEGs) between COPD and non-COPD samples were analyzed for each cell type using the Wilcoxon rank sum test. Gene set enrichment analysis was performed using the Gene Ontology Biological Process and Kyoto Encyclopedia of Gene and Genomes human databases under the following criteria: adjusted P value < 0.05, log fold-change > 1.5. An adjusted P value is a Bonferroni-corrected P value to avoid multiple comparison problem.
RESULTS
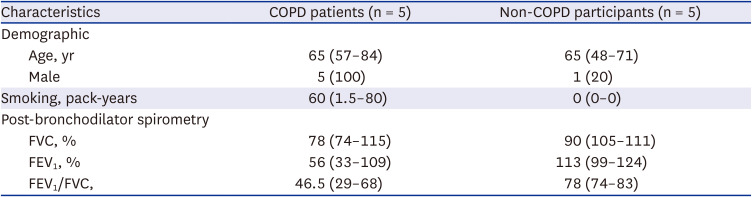
Five patients with COPD and five without COPD were included in the analysis. All COPD patients were men and had lower pulmonary function than the non-COPD participants (Table 1). Epithelial cells were successfully cultured from bronchoscopic brushing specimens obtained from all participants without complications. The MTT assay showed that 2% or less CSE did not affect cell viability but 4% induced cell death (Supplementary Fig. 1). Cultured cells were treated as either the control (medium only) or with 2% CSE for 24 hours and employed for proteomic analysis (Fig. 1).
Table 1
Basic characteristics of participants
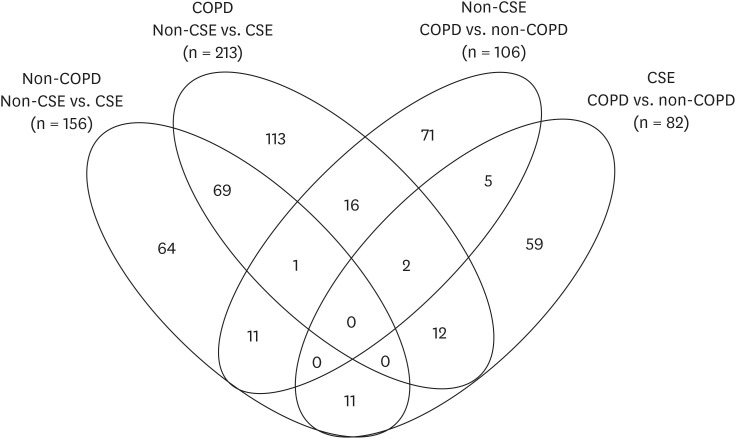
A total of 125,724 peptides, including 98,831 unique peptides, were characterized using MS, and subsequently, 7,953 proteins were identified. Initially, conventional analysis of DEPs was performed. A total of 156 DEPs were identified between CSE treated vs. non-CSE treated from non-COPD samples, and 213 DEPs from the corresponding samples derived from patients with COPD (Fig. 2, Supplementary Table 1). Only 70 DEPs were shared between these two DEPs above. Ferroptosis-related proteins, including ferritin heavy chain 1 (FTH1) and ferritin light chain (FTL), were identified in this set of overlapping DEPs, however, there were no significantly enriched pathways from these 70 DEPs. In addition, 106 DEPs were found between non-CSE treated COPD and non-COPD samples whereas 82 DEPs were found between CSE-treated COPD and non-COPD samples. There were no ferroptosis-related proteins identified from these DEPs (Fig. 2).
Fig. 2
Venn diagram and numbers of DEPs.
DEP = differentially expressed protein, CSE = cigarette smoking extract, COPD = chronic obstructive pulmonary disease.
To find the proteins that are most affected upon CSE exposure and to compare them between COPD and non-COPD directly, first we used an Extra Trees Classifier with the raw LFQ intensity data from COPD and non-COPD, separately. This ML-based algorithm calculated the importance of each protein in the process of classification between CSE-treated vs. non-CSE treated group. Set A was the feature importance of each protein from the non-COPD group, and set B was from COPD group (Fig. 1). Next, the top 100 proteins showing the highest differences between the COPD and non-COPD groups were selected (Supplementary Table 2), and a pathway analysis was performed. Interestingly, ferroptosis-related proteins, including FTH1, FTL, SLC7A11, and TP53, were shown to be the proteins that changed most in expression in response to CSE (adjusted P value = 4.172 × 10−6; Table 2); additionally, glutathione (GSH) metabolism was selected as a featured pathway in response to CSE (adjusted P value = 4.527 × 10−3).
Table 2
Featured pathways responsive to smoking exposure in the COPD group compared to the non-COPD group

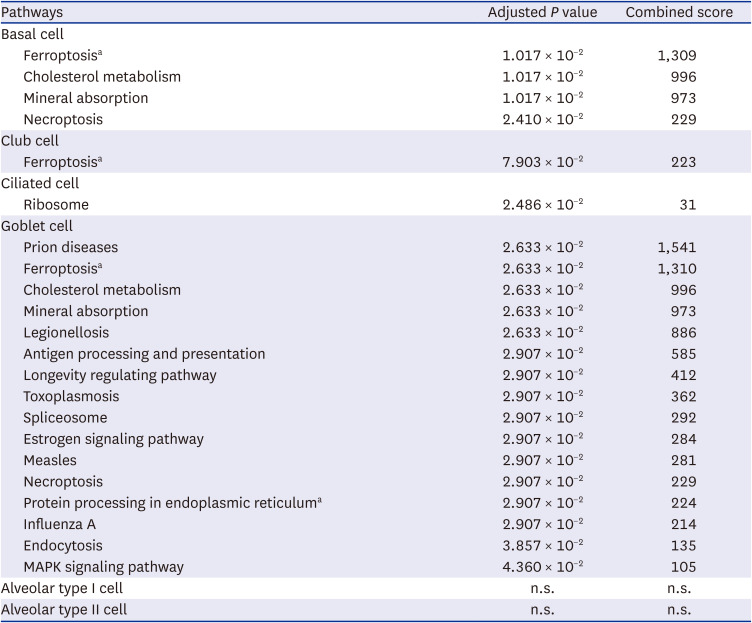
As an external validation of the featured pathways identified in this study, we reanalyzed publicly available single-cell RNA sequencing data from lung parenchyma cells from healthy controls and patients with COPD. Based on DEGs for each cell type, ferroptosis was selected as a featured pathway in basal cells, club cells, and goblet cells but not in ciliated cells (Table 3). We hypothesized that the ferroptosis-related signals from these epithelial cells were diluted in the bulk sequencing or proteomics data. Congruently, when we performed the same analysis of single-cell RNA sequencing data with broader cell categories, such as epithelial, myeloid, lymphoid, endothelial, and stromal cells, ferroptosis was not a distinct pathway in any of the other cell types (Supplementary Table 3).
Table 3
Featured pathways of chronic obstructive pulmonary disease single cell transcriptomics of epithelial cells
DISCUSSION
Airway epithelial cells play vital roles in the maintenance of respiratory homeostasis, including host defense, inflammation control, and tissue repair, all of which are important in the pathogenesis of COPD.19 COPD is associated with different types of regulatory cell death, including apoptosis20 and necroptosis21 in airway epithelial cells. Recent studies have reported that ferroptosis, a newly discovered form of regulated cell death, is also linked to COPD.2223 Ferroptosis has been identified in cancer cells and is widely observed in other diseases, including COPD.
In this study, we identified ferroptosis as a distinct signature of smoking exposure in cultured epithelial cells from COPD patients compared to those from non-COPD patients. The ferroptosis-related proteins, including FTH1, FTL, and SLC7A11, were also present in the results of conventional analysis; the DEP lists of CSE vs. non-CSE of COPD and those of non-COPD (Supplementary Table 1), which are consistent with the previous report.24 However, when we compared the effects of CSE treatment between the COPD group and the non-COPD group directly, the ferroptosis-related genes were not shown in the DEP list derived by conventional analysis (Supplementary Table 1). However, ML-based analysis provided the feature importance scores during the training process of Extra Trees Classifier, enabling us to directly compare the response to CSE. With our ML-based analysis, we revealed that the ferroptosis-related proteins responded more dramatically in COPD than in non-COPD.
The genes and proteins identified in this study have been reported to be significant signals of COPD in previous studies. A gene expression profile study using microarray analysis showed that FTL is one of 48 genes associated with COPD progression.25 A MS proteomic study of lung tissues also showed that FTL and FTH1 were two of the 25 proteins significantly associated with COPD.26 However, since only a few of the ferroptosis markers were present amongst dozens of markers detected in the previous studies, they were understandably dismissed.2526 This is mainly because lung tissue has many corresponding immune cells,1827 and ciliated cells, another dominant cell type in the airway,28 are not involved in ferroptosis, according to our analysis. In contrast to other studies, only epithelial cells were enriched in our study; therefore, ferroptosis could be selected as the most important featured pathway of COPD epithelial cells. Indeed, the single-cell RNA sequencing data of COPD and healthy controls showed that only basal cells, club cells, and goblet cells were enriched in ferroptosis, supporting our hypothesis.
In addition, the activity of post-translational modifications that regulate the degradation rate of proteins cannot be detected at the transcriptional level, and this might be another reason why ferroptosis was not distinguished in previous studies. For example, SLC7A11 was detected in this analysis and sustained the production of GSH,29 a key compound in the ferroptosis pathway. SLC7A11 is known to be regulated by ubiquitination in ferroptosis,29 however, previous studies did not establish SLC7A11 as a gene differentially expressed in COPD.
Ferroptosis has an iron-dependent mechanism and biological characteristics that are distinct from other regulated cell death processes, such as apoptosis and autophagy. Intracellular GSH depletion and iron accumulation led to an increased level of lipid peroxides through the Fenton reaction, resulting in a large increase in the number of radical oxygen species (ROS), which in turn promotes cell death.30 An in vitro and in vivo study showed a correlation between ferroptosis in airway epithelial cells and COPD pathogenesis. Cigarette smoke exposure initiates ferritin-selective autophagy (ferritinophagy), which leads to labile iron deposits, enhanced lipid peroxidation, and concomitant regulated cell death (ferroptosis) that are associated with the COPD phenotype, including emphysema. This CSE-induced ferroptosis-related emphysema was augmented in glutathione peroxidase 4 (Gpx4)+/− mice.24 GSH and Gpx4 depletion results in ferroptosis, suggesting that ferroptosis is linked to oxidative stress.31 GSH is depleted in acute smoking, leading to the prevalence of ROS and lung injury.32 Necrotic cell death via ferroptosis also provides danger-associated molecular patterns that induce and enhance the proinflammatory state.33 Hence, ferroptosis may contribute to COPD pathogenesis through cell death, increased oxidative stress, and proinflammatory responses.
Our study has limitations; 1) All COPD patients were male whereas there was only one male in the non-COPD participants in this study. This sex difference may cause sex-specific biomarkers to be mis-classified as COPD-specific biomarkers. The prevalence of COPD is higher in males and one of the challenging problems in mining biomarkers in COPD.34 Although ferroptosis is not thought to be a sex-specific biologic process, the confounding effects on ferroptosis from sex difference cannot be excluded in this study and further study is required. 2) The single cell data are based on tissue from patients with COPD and healthy controls but lacks information on smoking exposure prior to donation of the tissues. A better validation would be possible with larger set of single cell transcriptomics data with a detailed smoking history. However, we consider it is still valid for supporting the importance of ferroptosis in the airway epithelial cells, but not other cell types.
In conclusion, ML-based feature selection revealed that ferroptosis is the most distinctive difference between epithelial cells from patients with COPD vs. non-COPD patients that were exposed to CSE. Our study contributes to the understanding of the pathogenesis of COPD and the development of improved therapeutic strategies for COPD.
 XML Download
XML Download