

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
According to the World Health Organization, obesity is defined as abnormal or excessive fat accumulation that presents a health risk. In recent decades, with changes in diet and reduction in physical activity, obesity has become a global epidemic issue [1,2]. In 2016, an estimated 13% of the global adult population (11% of men and 15% of women) was obese [2]. Obesity is one of the susceptibility factors for type 2 diabetes mellitus (T2DM), and a study showed that patients with obesity are more than seven times more likely to develop T2DM than people with a healthy weight range [3]. Both obesity and T2DM are risk factors for adverse complications such as cardiovascular disease, and the cardiovascular risk increases significantly when both coexist. Therefore, the treatment of obesity and T2DM is a key issue in controlling cardiovascular morbidity and mortality.
Bariatric surgery is considered the most effective and sustainable treatment for obesity and obesity-related complications [4], significantly reducing T2DM and greatly improving the quality of life of patients with obesity. Studies have shown that bariatric surgery could better control blood glucose and improve remission rates for T2DM than medication alone, and the remission is durable [5-7]. The mutual regulatory network between obesity and T2DM, and the regulatory mechanism of bariatric surgery alleviating T2DM have always been the focus of scientists’ attention and the difficulty of research. Patients with obesity and T2DM who undergo bariatric surgery experience rapid improvement in glycemic control in a short period of time [8]. Significant enhancement in T2DM after 21 days of bariatric surgery has been reported before noticeable weight loss in patients with obesity [9]. Therefore, this present study focused on the mechanisms regulating the improvement of T2DM in the early phase (1 month) of bariatric surgery.
MicroRNAs (miRNAs) are a type of single-stranded, endogenous, non-coding RNAs, approximately 18 to 22 nucleotides in length [10]. They act on messenger RNAs by binding to the complementary sequence of the 3’ untranslated region, thereby affecting genes expression. MiRNAs can regulate 30% to 80% of genes expressed in the human genome [11], and each miRNA can target hundreds of genes while multiple miRNAs may only impact on one gene [12]. Thus, miRNAs form a complex genes regulatory network that controls almost every biological process (BP). Recent studies have reported that miRNAs perform a key role in obesity and its associated complications [13-15]. Moreover, miRNAs can regulate the uptake and utilization of glucose, the production and secretion of insulin, the maturation and differentiation of islet cells, and participate in BPs such as insulin resistance, glucose homeostasis and lipid metabolism, which may explain the pathogenesis of T2DM [16-19].
Given the close relationship between obesity and T2DM, the objects of this study were patients with obesity, and the changes of miRNAs in patients without T2DM or with T2DM were firstly compared and analyzed, which could explore the regulatory network of miRNAs in the occurrence and development of T2DM. Furthermore, for the significantly relieving effect of bariatric surgery on T2DM, this study continued to compare and analyze the changes of miRNAs in T2DM patients with obesity before and after bariatric surgery, which would reveal the regulatory network of miRNAs in the process of remission of T2DM. By comprehensively sorting out the above two comparative analyses, it is expected to find the most potential miRNAs regulating T2DM, thereby alleviating T2DM in patients with obesity.
METHODS
Recruitment
All patients from The First Affiliated Hospital of Jinan University (China) with a body mass index (BMI) ≥27.5 kg/m2, aged between 18 and 65 years, of either sex, who met the indications for bariatric surgery were included in this study. Fifteen patients with obesity but without T2DM and 15 patients with both T2DM and obesity were screened for this study, and all 30 patients underwent bariatric surgery, either sleeve gastrectomy (SG) or Roux-en-Y gastric bypass (RYGB). In addition, all patients were followed up for 1 month postoperatively. Of the 15 patients with T2DM, nine had newly discovered diabetes, and another six had varying degrees of diabetes with a mean duration of 5.06 years. These six diabetic patients were on no more than one glucose control medication and did not start any new glucose-lowering medications within 1 month of bariatric surgery. The study was ethically approved by the Institutional Review Board of The First Affiliated Hospital of Jinan University (No.: KY-2021-102). All participants provided informed written consent prior to the study, and the study assessments were conducted in accordance with the 1964 Helsinki declaration.
Serum samples preparation and extraction
All serum samples were obtained by standard venipuncture from patients who had fasted for at least 12 hours. The supernatant was acquired by low temperature centrifugation at 4,000 ×g for 10 minutes and stored at –80°C until use. The TRIzol Reagent (cat. NO. 15596026, Invitrogen, Waltham, MA, USA) was used to extract total RNA from serum samples under the guidance of the instructions. After total RNA quality determination and RNA integrity assessment, quantification was finally performed via Qubit 3.0 with QubitTM RNA Broad Range Assay kit (Q10210, Life Technologies, Carlsbad, CA, USA). KCDigitalTM small RNA Library Prep Kit for Illumina (cat. NO. DR08602, Wuhan Seqhealth Co. Ltd., Wuhan, China) was applied to construct the miRNA libraries. Lastly, the gel-purified cDNA libraries were also isolated, purified and quantified by Qubit 3.0.
Analysis of miRNAs sequencing data
Equimolar amounts of libraries were pooled and sequenced on Hiseq X-10 sequencer (Illumina) with PE150 model. MiRNA sequencing experiment and high throughput sequencing were performed by Seqhealth Technology Co. Ltd. Raw sequencing data were further filtered low quality reads, removed adaptor sequences and eliminated duplications. The clean sequences of each sample were mapped to the reference genome of Homo sapiens GRCh38 using Bowtie (version 1.1.2). Subsequently, sequences alignment was performed using mirdeep2 (version 2.0.0.8) and known miRNAs were detected. The reads were mapped to mature and precursor miRNAs obtained from the miRNAs database miRBase (https://www.mirbase.org/).
The edgeR package (version 3.12.1) was used to normalize the read counts of known miRNAs and to calculate the logarithmic value of counts per million (log CPM) for normalization analysis. The differentially expressed miRNAs (DEMs) between the groups were determined with |log2 fold change (FC)| >1 and P value <0.05. Volcano plots and heat maps of DEMs were done by R software version 4.1.2 (R Foundation for Statistical Computing, Vienna, Austria). The online Venn diagram production tool Venny (https://bioinfogp.cnb.csic.es/tools/venny/) was used to analyze the intersection of different variations of miRNAs across groups.
Analysis of miRNA target genes
Two databases, miRanda (version 3.3a; https://bioweb.pasteur.fr/packages/pack@miRanda@3.3a) and RNAhybrid (version 2.1.1; https://bibiserv.cebitec.uni-bielefeld.de/download/tools/rnahybrid.html), were used to predict the target genes and their intersection was taken as the list of target genes. P value <0.05 was defined as significant in the Gene Ontology (GO) terms and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis, both of which were plotted by a free online platform for data analysis and visualization (https://www.bioinformatics.com.cn). The Cytoscape (version 3.8.2; https://cytoscape.org/release_notes_3_8_2.html) tool cytoHubba obtained key target genes by analyzing the results exported from the STRING (version 11.5; https://cytoscape.org/release_notes_3_8_2.html) database.
Data calculation and formula
Homeostasis model assessment of insulin resistance (HOMA-IR), an authoritative index for assessing insulin resistance, is calculated as fasting blood glucose×fasting insulin/22.5. Weight change (weight loss=preoperative weight−postoperative weight), BMI change (∆BMI=preoperative BMI−postoperative BMI), and percentage total weight loss—%TWL=[(pre-operative weight−postoperative weight)/preoperative weight]×100%— are common indicators to assess the efficacy of weight loss before and after bariatric surgery.
Statistical analysis
Statistical analysis was performed utilizing IBM SPSS Statistics version 24 (IBM Co., Armonk, NY, USA) and R software version 4.1.2. In the comparison of patients with and without T2DM, or patients who underwent SG or RYGB preoperatively and postoperatively, we applied two independent samples t-test (two-tailed) for continuous variables and the Fisher’s exact test for categorical variables. Data before and after bariatric surgery were tested with two paired samples t-test. The relationships between continuous variables were analyzed by Pearson correlation. P value less than 0.05 was considered to be statistically significant.
RESULTS
Clinical characteristics
To explore the DEMs caused by whether patients with obesity have T2DM or not, a total of 30 patients with obesity were included in this study, 15 of whom did not have T2DM and 15 of whom had T2DM. As shown in Table 1, except for glycated serum protein (GSP, P=0.008), glycosylated hemoglobin (HbA1c, P=0.000), fasting glucose (GLU, P=0.001), and HOMA-IR (P=0.015), there were no significant differences between the two groups in other indicators such as anthropometric indicators, lipid metabolism-related indicators, and weight loss effectiveness evaluation indicators. GSP and GLU could reflect the short term glycemic control status. To further investigate the DEMs of early remission of T2DM by bariatric surgery, patients were followed up for 1 month after bariatric surgery in this study. We were surprised to find remarkable improvements in weight, BMI, triglycerides (TG), total cholesterol, high density lipoprotein, GSP, HbA1c, GLU, insulin, and HOMA-IR in T2DM patients at only 1 month postoperatively. These two interesting sets of changes set the stage for our next step of DEMs analysis.
In addition to this, 17 of these 30 patients underwent SG, while the other 13 underwent RYGB, and we further compared the clinical characteristics of these patients before and after the SG or RYGB procedures (Supplementary Table 1). Before the surgery, there were significant differences in weight (P=0.039), BMI (P=0.024), and waist to hip ratio (P=0.022) between SG and RYGB. And 1 month after surgery, SG and RYGB displayed significant differences in BMI (P=0.037), weight loss (P=0.007), and change in BMI (∆BMI, P=0.025). Patients who underwent RYGB had higher weight and BMI relative to those who underwent SG, as a result of a joint choice between surgeons and patients. In contrast, the two surgical procedures did not show significant differences in lipid metabolism-related or glucose metabolism-related indicators, either preoperatively or postoperatively.
DEMs in different groups
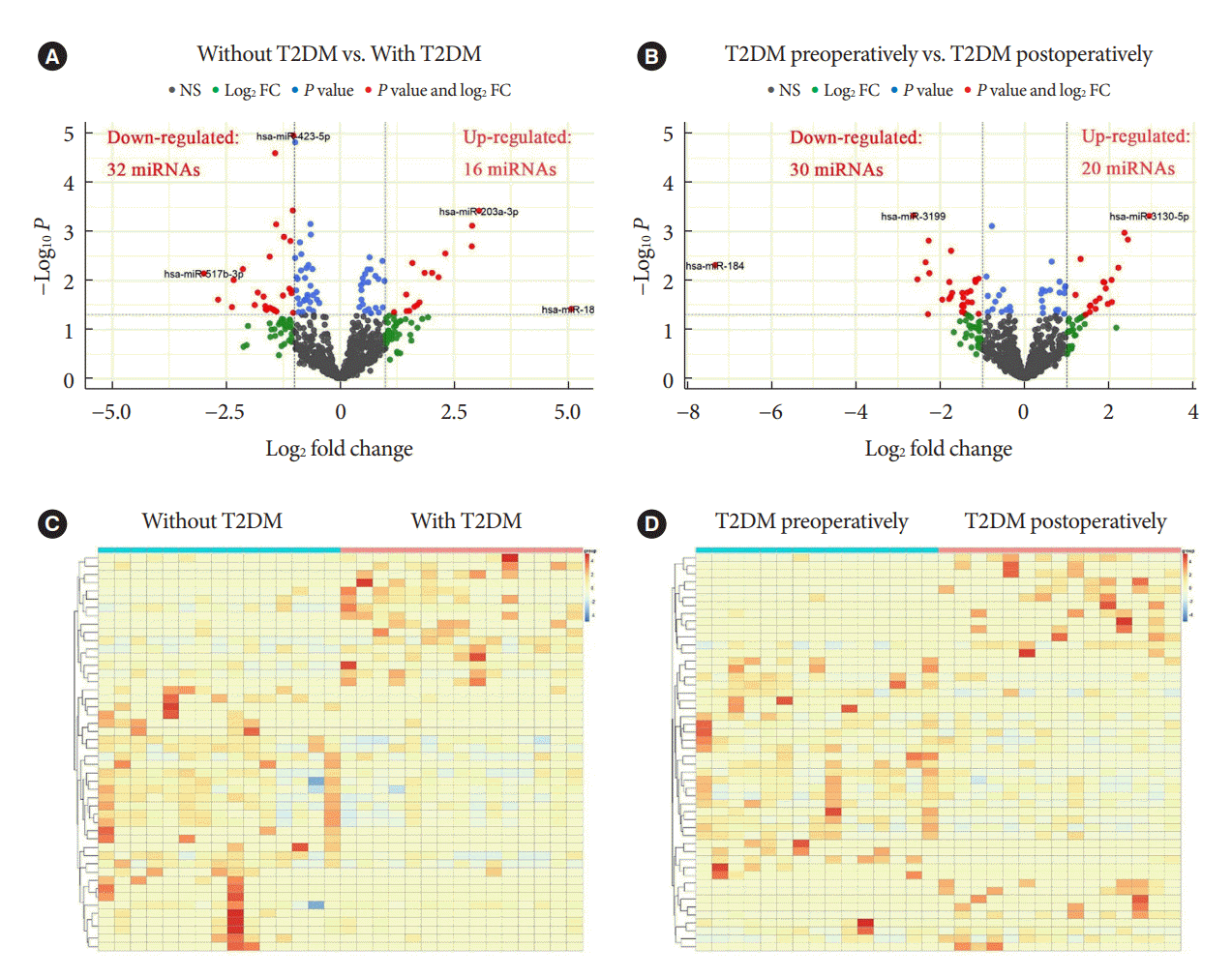
Known miRNAs were identified by mapping with databases such as Bowtie, mirdeep2, miRBase, etc. Intercomparison of different groups was performed after edgeR package normalization process. We define log2 FC >1 and P<0.05 as DEMs with up-regulated expression, while log2 FC <1 and P<0.05 as DEMs with down-regulated expression. When compared to patients with obesity but without T2DM, patients with both T2DM and obesity demonstrated 16 up-regulated DEMs and 32 down-regulated DEMs (red dots in Fig. 1A). In addition, T2DM patients with obesity displayed 20 up-regulated DEMs and 30 down-regulated DEMs postoperatively relative to preoperatively (red dots in Fig. 1B). The volcano maps showed several DEMs with the most significant or largest fold change. Furthermore, cluster comparison by heatmaps found that DEMs of different groups were significantly different (Fig. 1C and D). These data indicated that miRNAs expression profiles were different in patients with obesity, either with and without T2DM, or before and after bariatric surgery.
The intersection of DEMs in different groups
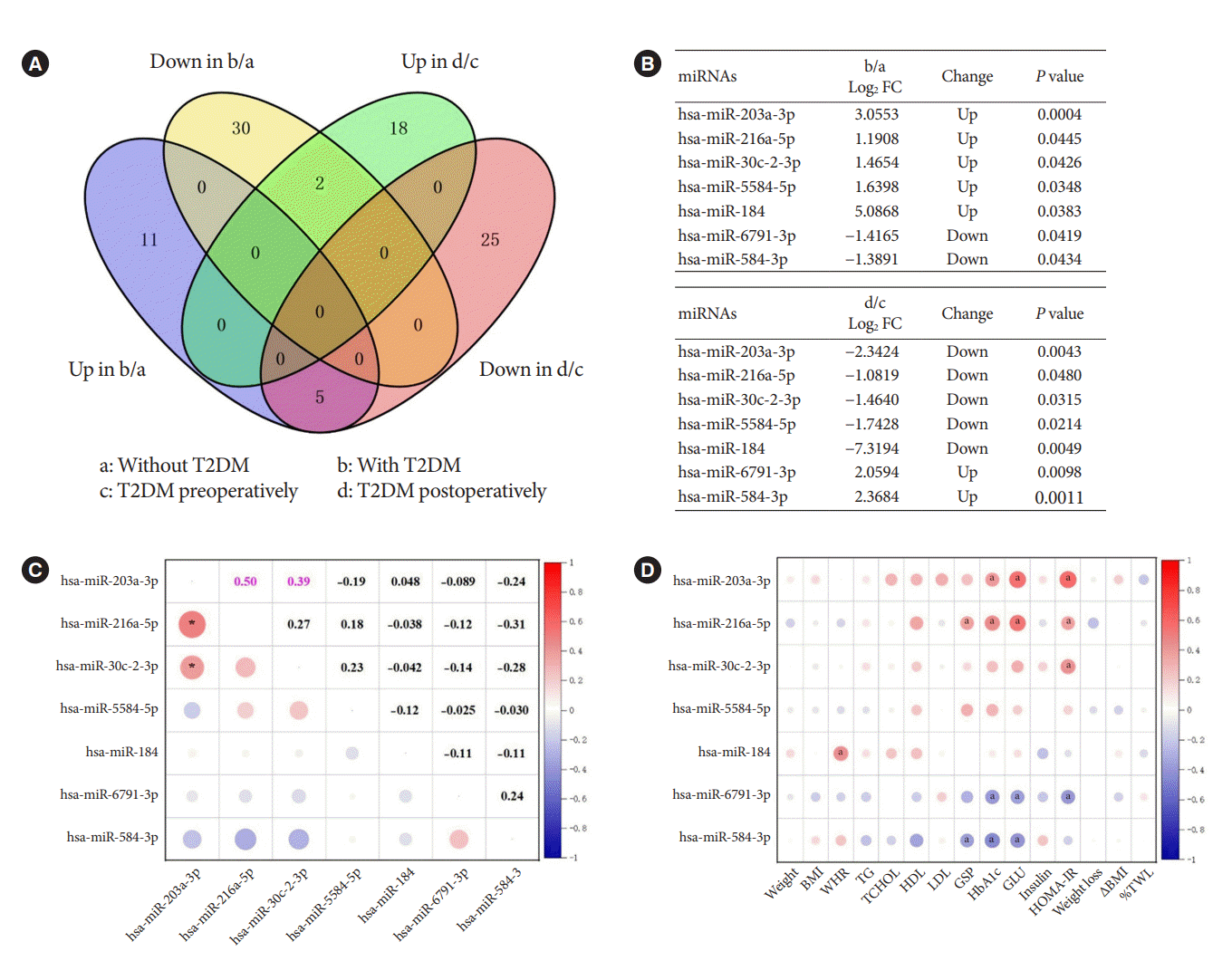
In the previous analysis, we identified DEMs in obesity with and without T2DM, and DEMs in obesity with T2DM before and after bariatric surgery. To investigate the pathogenesis of T2DM in patients with obesity and the possible mechanism of remission after bariatric surgery, we used Venn diagrams to intersect the aforementioned DEMs and find their co-varying DEMs (Fig. 2A). We finally found seven co-variant DEMs (Fig. 2B). While compared to obesity without T2DM, hsa-miR-203a-3p, hsa-miR-216a-5p, hsa-miR-30c-2-3p, hsa-miR-5584-5p, and hsa-miR-184 were up-regulated in obesity with T2DM. However, they were indeed down-regulated in obesity with T2DM 1 month after surgery. This was an interesting finding, demonstrating that the increased expression of these miRNAs might be associated with the onset, and progression of T2DM. In contrast, the opposite results were observed for hsa-miR6791-3p and hsa-miR-584-3p, suggesting that a decrease in their expression correlated with progression of T2DM and an increase in their expression correlated with postoperative remission of T2DM.
As the two most prevailing procedures for bariatric surgery, SG and RYGB might also differ in the expression of miRNAs before and after surgery, denoting different mechanisms. Consequently, we used the surgical procedures as subgroups to profile the changes of these seven miRNAs which are extremely associated with T2DM. As shown in Supplementary Table 2, hsa-miR-216a-5p (P=0.013) was down-regulated after SG surgery, which was consistent with the trend of change in Fig. 2B. However, hsa-miR-203a-3p (P=0.000) and hsa-miR-30c-2-3p (P=0.004) were rising after RYGB surgery, contradicting the trend of Fig. 2B. These differences might be caused by the low or even zero expression of these seven miRNAs in some cases when analyzed by surgical methods as subgroup.
In further, Pearson’s correlation analysis of the seven miRNAs from 30 patients preoperatively revealed a significant positive correlation between hsa-miR-203a-3p and hsa-miR-216a-5p (r=0.50, P<0.05), and hsa-miR-203a-3p and hsa-miR-30c-2-3p (r=0.39, P<0.05) (Fig. 2C). Their trends were also the same in both groups, indicating their mutual synergistic relationship within them. To initially validate the relationship between these miRNAs and T2DM, we further subjected these preoperative miRNAs and clinical characteristics of 30 preoperative patients to Pearson correlation analysis. The expression of preoperative miRNAs was all represented by log CPM. Most of the five miRNAs up-regulated in T2DM patients with obesity were positively correlated with glucose metabolism indicators such as GSP, HbA1c, GLU and HOMA-IR, while the two down-regulated miRNAs were negatively correlated with these indicators (Fig. 2D). These results further characterized the changes of these seven miRNAs in T2DM.
It is worth noting that not all patients with T2DM could be relieved by bariatric surgery. To discriminate the DEMs between the T2DM remission group and the T2DM non-remission group after bariatric surgery, we defined T2DM remission as fasting glucose ≥7.00 mmol/L and T2DM non-remission otherwise. As shown in Supplementary Fig. 1A and B, there were a total of 69 DEMs in the T2DM remission group, while there were a total of 17 DEMs in the T2DM non-remission group. There was little overlap in the miRNAs profiles of the two groups, except for one miRNA. We further analyzed the change characteristics of the above seven miRNAs that are extremely associated with T2DM in both groups. As displayed in Supplementary Fig. 1C and D, we discovered that all these seven miRNAs were significantly changed in the T2DM remission group, which was almost consistent with the results in Fig. 2B. In contrast, in the T2DM non-remission group, these seven miRNAs were not significantly altered before and after surgery. These results further illustrated the crucial regulatory role of these seven miRNAs in T2DM.
GO and KEGG enrichment analysis
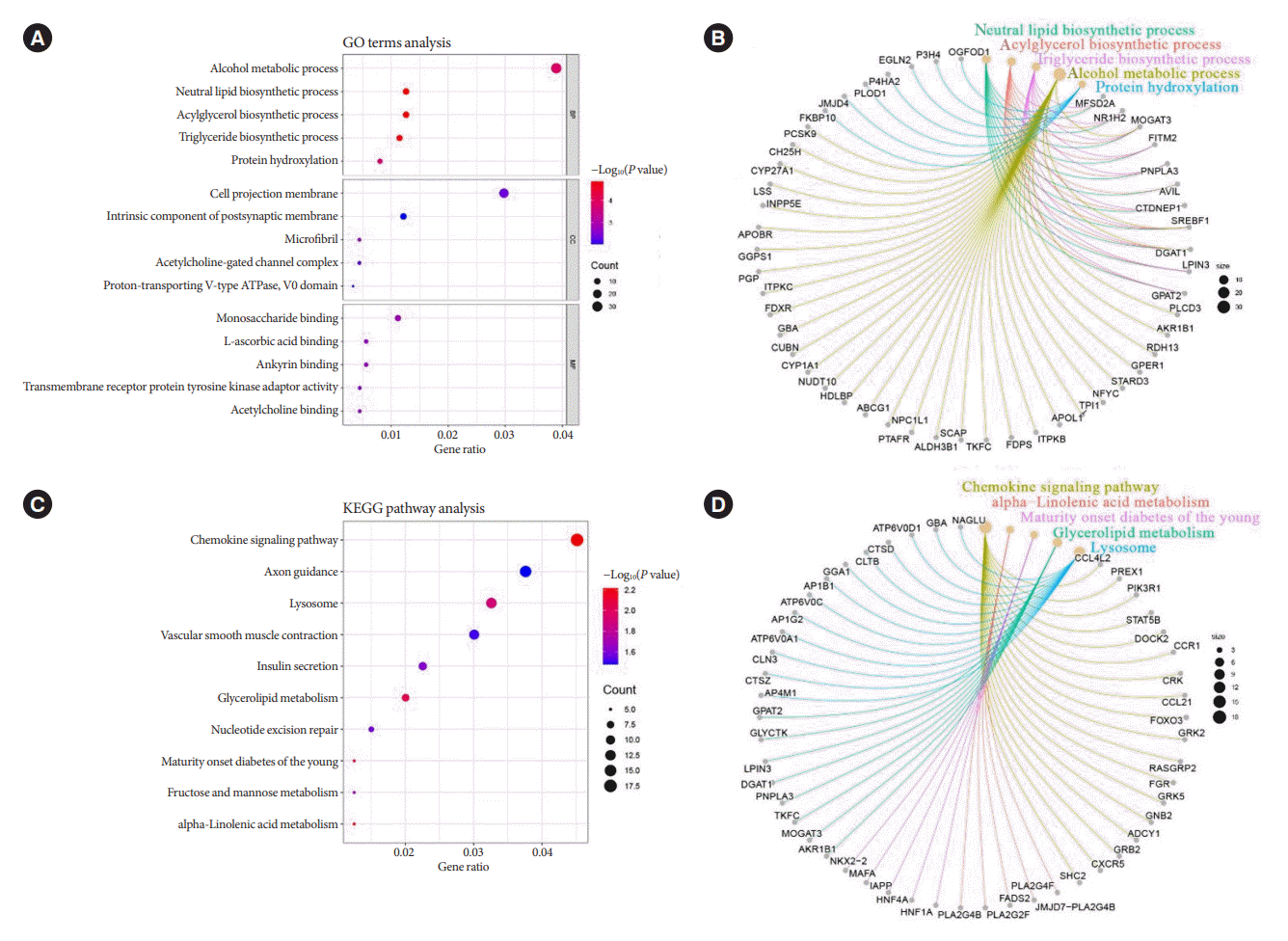
The predicted target genes of the seven miRNAs were identified by taking the intersection of miRanda and RNAhybrid databases for subsequent analysis. Fig. 3A showed the five most significant terms of GO terms in the three major annotation categories of BP, cellular component, and molecular function, respectively. Among all the taxonomic annotations, the five most significant ones were neutral lipid biosynthetic process, acylglycerol biosynthetic process, TG biosynthetic process, alcohol metabolic process, and protein hydroxylation, respectively. The correlation between the taxonomic annotations and the target genes was shown in Fig. 3B. Correspondingly, Fig. 3C and D presented the KEGG pathway analysis and the correlation of the five most significant pathways with the target genes. The five most significant pathways were chemokine signaling pathway, alpha-linolenic acid metabolism, Maturity onset diabetes of the young, glycerolipid metabolism, and lysosome, respectively. Both GO terms analysis and KEGG pathway analysis revealed important links between the target genes of the seven miRNAs and glycolipid metabolism.
Hub gene analysis
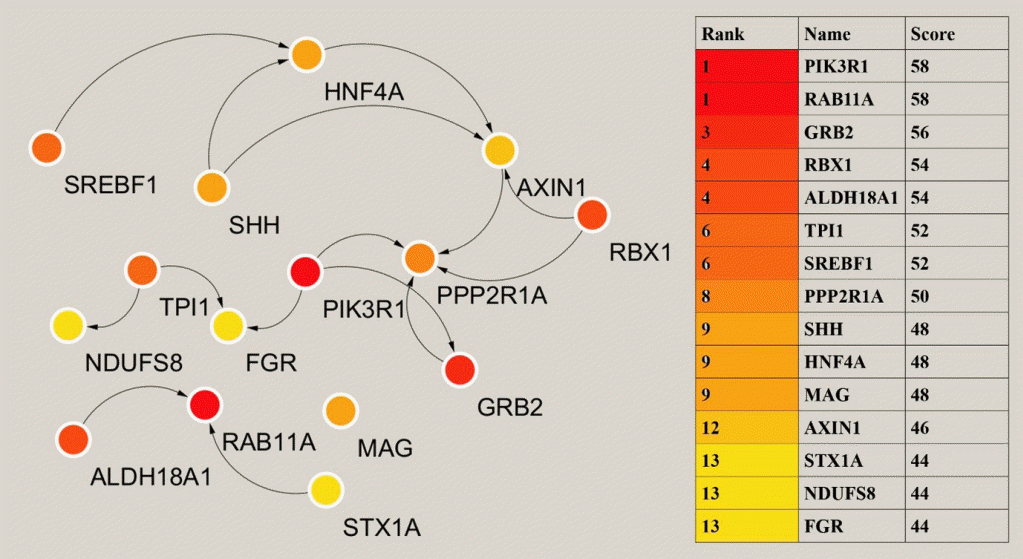
The analysis of key or central genes is beneficial to find the most representative genes in regulating T2DM. To obtain them, we imported the list of target genes into STRING database, and then performed hub genes analysis by Cytoscape’s plugin cytoHubba. CytoHubba provided 11 models for analysis, and we finally chose the degree method to rank the top 15 hub genes. The relationship and specific names between the top 15 hub genes were shown in Fig. 4.
DISCUSSION
MiRNAs are related to multiple aspects of glycolipid metabolism involved in T2DM and may play a role in the pathogenesis of obesity and its associated complications. Zampetaki et al. [20] revealed the characteristics of plasma miRNAs in diabetic patients, while 13 candidate miRNAs were identified. A growing number of studies have demonstrated that miRNAs and insulin resistance [17,21,22], glucose metabolism [15,16,23], lipid metabolism [24,25], and β-cell function [26,27] were closely related, and these altered miRNAs may be potential targets for the prediction and treatment of T2DM. Bariatric surgery is considered the most effective intervention for the treatment of obesity and T2DM, with significant weight loss and remission of T2DM in the short term [28], and this effect is durable. It has been shown that some miRNAs changed significantly after bariatric surgery and that these miRNAs played an important and unique role in improving metabolic indicators rather than acting through weight reduction or body fat factors [29-32], explaining to some extent the mechanisms involved in T2DM remission after bariatric surgery. Atkin et al. [9] analyzed 29 patients with severe obesity and diabetes who underwent RYGB surgery, and they found significant changes in seven miRNAs 21 days after bariatric surgery, which occurred prior to significant weight loss. We strongly approve of the clinical relevance of the study by Atkin et al. [9]. Unfortunately, they did not perform further functional categorization of miRNAs, analysis of the relationship with clinical characteristics, or prediction of relevant target genes. These elements were supplemented in our work.
In the present study, we performed a comparative analysis of serum miRNAs in patients with and without T2DM in obesity to find the DEMs of both. Then, we found significant improvements in metabolic indicators in obesity with T2DM at 1 month after bariatric surgery. Based on this, we further analyzed the characteristics of DEMs before and after surgery. We pooled the above DEMs for analysis to identify co-variant miRNAs and map the miRNAs profiles of T2DM patients with obesity, which might reveal the mechanisms of T2DM progression in obesity and remission in bariatric surgery. We finally found seven co-altered miRNAs that might be key miRNAs in the regulation of T2DM. Relative to obesity without T2DM, obesity with T2DM had five up-regulated miRNAs and two down-regulated miRNAs that changed in opposite ways after bariatric surgery for obesity with T2DM.
Although most studies have found that these seven miRNAs are associated with cancer, some previous studies have reported the possible mechanisms of action of these miRNAs. Zhang et al. [33] found elevated plasma miR-203a-3p levels in a rat hyperlipidemia model and verified that atorvastatin functions as a regulator of lipids by inhibiting the expression of miR203a-3p. MiR-216a-5p might play a role in diabetic nephropathy [34]. In mice fed a high-fat diet, miR-30c-2-3p was found to be significantly up-regulated in brown fat, inducing adipogenesis and inflammatory stress, which in turn affects energy metabolism [35]. These studies demonstrated the important role of the above mentioned miRNAs in glycolipid metabolism. Further Pearson correlation analysis of these miRNAs with clinical characteristics revealed that most of the five previously mentioned miRNAs were positively correlated with glucose metabolic indexes and insulin resistance, suggesting that their increase induces the development of T2DM. The other two miRNAs were negatively correlated with these metabolic indicators, suggesting that their decrease would likewise lead to the development of T2DM. Moreover, the target genes enrichment analysis of these seven miRNAs demonstrated the terms or pathways associated with T2DM in both the five most significant GO terms and the five most significant KEGG pathways. In addition to this, the expression profiles of miRNAs differ in SG and RYGB surgeries, which might indicate different mechanisms [36]. When we analyzed the changes of seven miRNAs before and after surgery using the surgical procedures as subgroups, we found that their change characteristics differed from the above results. Future studies should delve into the expression profiles of miRNAs in the very short term with different surgical strategies.
MiRNAs exert their effects by regulating gene expression, and we therefore, predicted the target genes of miRNAs. The hub genes were further analyzed by Cytoscape to identify the genes at the center and reveal the key regulatory genes of T2DM in patients with obesity. Phosphoinositide-3-kinase regulatory subunit 1 (PIK3R1) plays an important role in the metabolic action of insulin and is required for increased glucose uptake and glycogen synthesis in insulin-stimulated insulin-sensitive tissues, and mutations in this gene are associated with insulin resistance [37,38]. PIK3R1 was one of the highest scoring genes in hub genes analysis. FGR proto-oncogene, Src family tyrosine kinase (FGR) promotes the phosphorylation of PIK3R1 thereby facilitating its activation [39]. Triosephate isomerase 1 (TPI1) encodes an enzyme consisting of two identical proteins that catalyze the isomerization of glyceraldehyde 3-phosphate and dihydroxyacetone phosphate during glycolysis and gluconeogenesis [40]. Mutations in this gene have been associated with a deficiency of propanose phosphate isomerase, and related pathways include glucose metabolism. Sterol regulatory element binding transcription factor 1 (SREBF1) regulates metabolism and the mammalian target of rapamycin (mTOR) signaling pathway, and is a key transcription factor regulating gene expression of cholesterol biosynthesis and lipid homeostasis [41]. It plays a role in the nutritional regulation of fatty acids and TG in lipogenic organs [42]. Syntaxin 1A (STX1A) encodes a member of the synaptic protein superfamily that plays an important role in the cytokinesis and endocytosis of hormones and neurotransmitters [43], and has also been shown to have a similar role in the cytokinesis of hormones such as insulin or glucagon-like peptide 1 [44]. NADH: ubiquinone oxidoreductase core subunit S8 (NDUFS8) encodes a subunit of mitochondrial nicotinamide adenine dinucleotide (NADH) that catalyzes the electron transfer of NADH through the respiratory chain and is closely involved in respiratory electron transport, adenosine triphosphate (ATP) synthesis and uncoupling protein thermogenesis, potentially a key gene for improved energy metabolism after bariatric surgery [45,46]. These hub genes regulate multiple aspects related to T2DM and are highly promising therapeutic targets.
Overall, we depicted miRNAs profiles for obesity with T2DM relative to obesity without T2DM, as well as for T2DM patients postoperatively relative to T2DM patients preoperatively. Further analysis intersected seven miRNAs that showed opposite features of change in the two miRNAs profiles, suggesting their important role in T2DM. The seven screened miRNAs showed different expression profiles in SG and RYGB. Target genes analysis of miRNAs revealed enrichment patterns for most of these target genes associated with T2DM, both in GO terms and in the KEGG pathway. We also identified genes that might be key regulators of T2DM. These findings certainly provided targets that could be intervened in the management of T2DM.
 XML Download
XML Download