

 PDF
PDF Citation
Citation Print
Print



Candida albicans, a predominant human fungal pathogen, causes both mucosal and bloodstream infections (BSIs), and fluconazole is one of the most widely prescribed antifungal agents used to treat these infections [1, 2]. Acquired fluconazole resistance (FR) in C. albicans has been reported at its highest frequency in HIV-infected patients with oropharyngeal candidiasis as well as in patients with recurrent vaginal candidiasis [2, 3]. The long-term use of fluconazole for prophylaxis or treatment of mucosal C. albicans infections can lead to selective pressure, resulting in the emergence of acquired FR in C. albicans [2, 3]. The main mechanisms responsible for acquired FR in C. albicans from mucosal infections include mutations or overexpression of ERG11, which encodes an enzyme targeting the drug (lanosterol 14α-demethylase), and the overexpression of genes encoding efflux pumps (CDR1, CDR2, and MDR1) [2, 4, 5]. The overexpression of FR-associated genes occurs mainly due to gain-of-function (GoF) mutations in the transcription factor-encoding genes TAC1 (involved in CDR1 and CDR2 regulation), MRR1 (involved in MDR1 regulation), and UPC2 (involved in ERG11 regulation) [2, 5-7].
In contrast to mucosal isolates with FR rates of 12%–22%, the rates of FR among BSI isolates of C. albicans are low (0.06%–2.3%); this could be partly because of the relatively short-term use of antifungal agents for the treatment of candidemia [2, 8-10]. Among 2,712 C. albicans BSI isolates obtained from Korean multicenter surveillance studies during 2006–2021, only 14 (0.5%) were determined to be fluconazole non-susceptible (FNS; minimum inhibitory concentration [MIC] ≥4 μg/mL). To date, only few studies have characterized the molecular and clinical features of BSI isolates of C. albicans with acquired FR [8]. Therefore, we investigated the gene mutations causing amino acid substitutions (AASs) in ERG11, TAC1, MRR1, and UPC2; their genotypic relationships; and the clinical features of FNS BSI isolates of C. albicans that were submitted to Chonnam National University Hospital from Korean multicenter surveillance studies over 15 years (2006–2021).
We assessed 26 BSI isolates of C. albicans: 11 FR (MIC ≥8 μg/mL), three fluconazole-susceptible dose-dependent (F-SDD; MIC, 4 μg/mL), and 12 control fluconazole-susceptible (FS; MIC, 0.25–0.5 μg/mL) isolates. All isolates were identified using matrix-assisted laser desorption/ionization-time of flight mass spectrometry (Bruker Biotyper library v. 4.0; Bruker Daltonics GmbH, Bremen, Germany) or by sequencing the D1/D2 domains of the 26S rRNA gene [11]. In vitro antifungal susceptibility testing was performed with the Sensititre Yeast One system (Thermo Fisher Scientific Inc., Cleveland, OH, USA). Sequence analyses of ERG11, TAC1, MRR1, and UPC2 were conducted as described previously [6, 12, 13]. All isolates were genotyped using multi-locus sequence typing (MLST); each strain was assigned a diploid sequence type (DST) reflecting the combination of the genotypes of seven genes in the MLST database (https://pubmlst.org/organisms/candida-albicans), and a dendrogram was constructed [14]. Clinical information for all 14 patients with FNS C. albicans BSI isolates was collected retrospectively [15]. This study was approved by the Institutional Review Board of Chonnam National University Hospital, Gwangju, Korea (approval No. CNUH-2014-290) that also waived the requirement for informed consent.
Table 1 shows the results of antifungal susceptibility testing and ERG11, TAC1, MRR1, and UPC2 sequencing of all 14 FNS (11 FR [R1–R11] and 3 F-SDD [D1–D3]) and 12 FS (S1–S12) C. albicans BSI isolates. No isolate was found to be resistant to amphotericin B or the three echinocandins. The sequencing results of all 14 FNS C. albicans isolates were compared with those of 12 FS control isolates and previously reported data [2, 6, 7, 16-20]; five Erg11p, seven Tac1p, and one Mrr1p AASs were found in only the FNS isolates. Of the five Erg11p AASs that were found in only the FNS isolates, two (R264T and D428N) were potentially novel; three AAS (K143R, F145L, and G464S) that were found in eight FNS isolates were confirmed to cause FR through in vitro experiments [16]. Of the seven AASs in Tac1p that were not present in the FS isolates, three (T225A, A736T, and A736V) AASs that were found in six FNS isolates were previously described as GoF mutations [2]; another (R673L) AAS found in one FNS isolate was previously described in azole-resistant isolates [2, 18-20]. The remaining three (Y269H, L744I, and N972K) Tac1p AASs from four FNS isolates were potentially novel, although N972K occurred at a position already described in FR isolates [19]. Only one substitution in Mrr1p (N33S), which was found in one FNS isolate, was not described previously [2, 7]. Although it is unclear whether the newly identified AASs in this study result in FR, the Erg11p, Tac1p, Mrr1p, and Upc2p AASs that were either novel or described in azole-resistant isolates were found in 8 (57.1%), 11 (78.6%), 1 (7.1%), and 0 (0%) isolates, respectively. A previous study characterized azole resistance mechanisms in five invasive FNS C. albicans isolates that had been collected in 29 countries in 2014 and 2015. The authors found that MDR1 overexpression (three isolates) was more common than CDR2 expression (one isolate) or ERG11 mutation (one isolate) [10]. In contrast, our results suggested that ERG11 mutations and CDR overexpression are likely the dominant mechanisms of FR in C. albicans BSI isolates from Korean hospitals.
Of the 14 FNS isolates, 12 exhibited weak FR (MICs: 4–16 mg/L) without voriconazole resistance, whereas the remaining two isolates (R1 and R2) showed high MICs for fluconazole (>256 mg/L) and voriconazole (>8 mg/L). Isolate R1 harbored L744I in Tac1p (new AAS), which might be the major contributor to CDR-mediated azole resistance; isolate R2 harbored Erg11p K143R. Of the five Erg11p K143R isolates with variable MICs for fluconazole (8–>256 mg/L), isolate R2 showed the highest fluconazole MIC; however, it did not show FR-specific Tac1p, Mrr1p, or Upc2p AASs. Two FNS isolates (R8 and D2) did not show any of the FR-associated AASs that were evaluated in this study. CDR1/CDR2 and MDR1 overexpression can be explained by TAC1 and MRR1 GoF mutations, but ERG11 overexpression is not always associated with UPC2 GoF mutations, suggesting the existence of additional regulators [21]. Thus, these isolates might have other resistance mechanisms, such as the overexpression of ERG11, which could not be detected in this study.
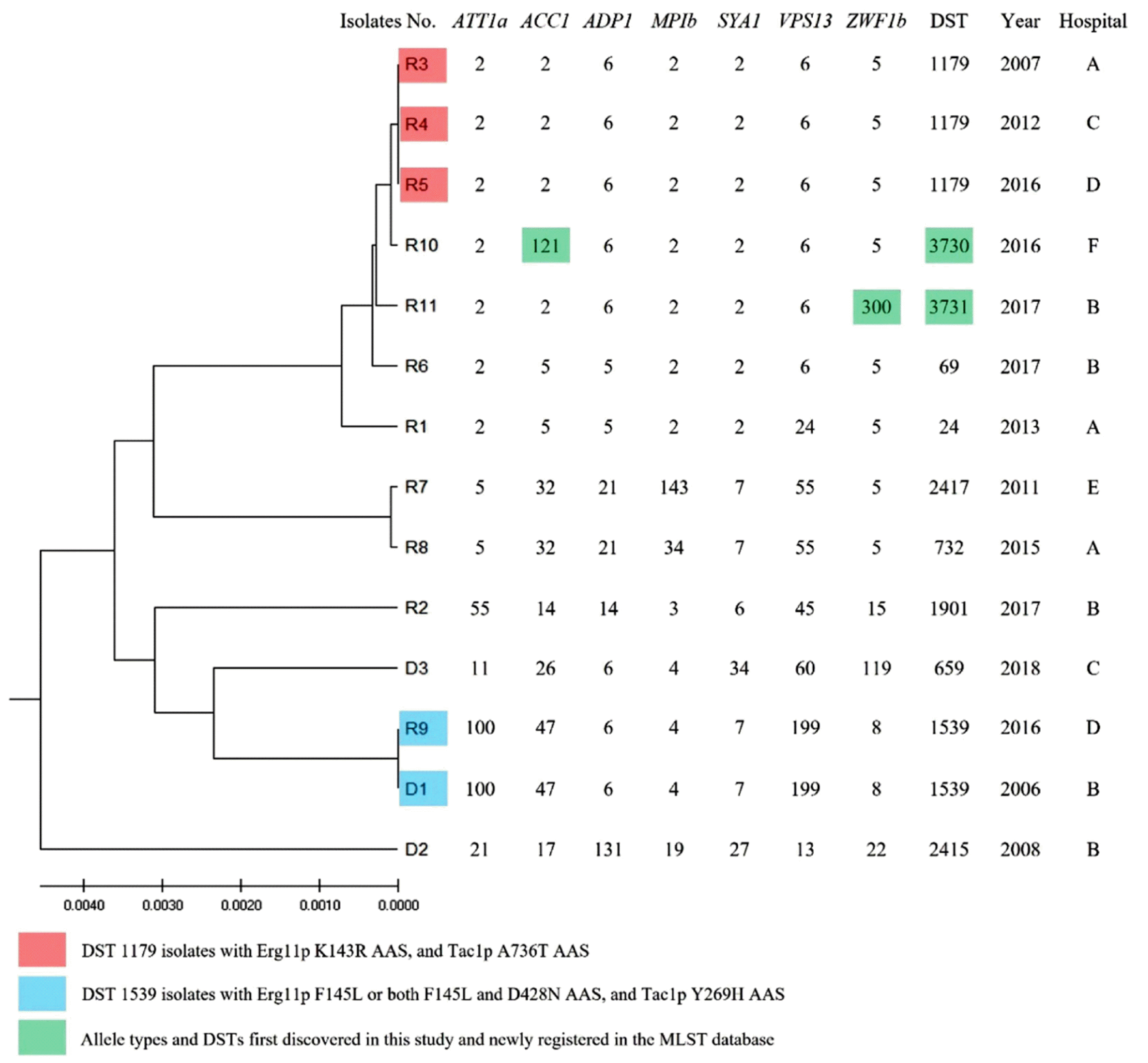
The MLST results for the 14 FNS isolates showed that nine isolates had different DSTs, whereas three and two isolates belonged to the DSTs 1179 and 1539, respectively (Fig. 1). Three DST 1179 isolates (R3–R5) were isolated at three different hospitals but had the same AASs in Erg11p (K143R) and Tac1p (A736T). Two isolates of DST 1539 (R9 and D1) were also isolated at different hospitals, and they had similar Erg11p AASs (F145L and F145L+D428N) and the same Tac1p (Y269H) AAS. TAC1 is located on the left arm of chromosome 5, where ERG11 is also located, and a combination of TAC1 and ERG11 point mutations has been suggested to contribute to an increased MIC for fluconazole among azole-resistant isolates [17]. Overall, 50.0% (7/14) of the FNS isolates showed combined Erg11p and Tac1p AASs in this study.
The clinical features of all 14 patients are summarized in Table 2. All 14 patients were adults with various underlying diseases, but no patient was infected with HIV. Previous amphotericin B (two patients) or fluconazole (one patient) exposure was identified in only three patients, indicating that almost all (92.9%) FNS C. albicans BSI isolates were from patients not previously exposed to azole. Among the 14 patients with FNS BSIs, eight had a fatal outcome within 30 days, three (R5, R7, and D3) died without receiving antifungal therapy, two (R4 and D1) died despite fluconazole therapy, and three (R1, R6, and R8) with a hematological malignancy died despite >3 days of echinocandin or amphotericin B therapy. The overall 30-day mortality rate of the patients was 57.1% (8/14), which was higher than the mean 30-day mortality rate (36.4%, 123/338) of patients with C. albicans BSIs reported at 11 Korean hospitals from 2017 to 2018, although the difference was not significant [15].
Given the marked genetic diversity among Korean C. albicans BSI isolates in our previous MLST study [14], it is interesting that in the present study, five FNS isolates shared two DSTs (1179 and 1539). Additionally, the dendrogram obtained through MLST testing revealed that three isolates of DST 1179 (isolates R3–R5) were more closely related to four FNS isolates (R1, R6, R10, and R11) (Fig. 1). However, there was no time- or location-based clustering of these isolates, which excludes the potential of cross-transmission in the hospitals. Alternatively, some patients could accidentally acquire clonal FNS isolates of C. albicans already present in the environment of healthcare settings in Korea, leading to the development of healthcare-associated BSIs; further studies are needed to confirm this possibility.
In summary, our results showed that most FNS C. albicans BSI isolates from Korean hospitals harbor mutations in ERG11 or TAC1 and that fungemia can develop without azole exposure. This is the first study to describe both the molecular and clinical features of FNS BSI isolates of C. albicans obtained from candidemia surveillance studies.
 XML Download
XML Download