

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Migraine is one of the most prevalent neurologic disorders in the world [1]. The Global Burden of Disease Study 2016 confirmed migraine as the second leading cause of disabling neurological disorder [2]. Migraine attacks are typically reported by patients to be severe, unilateral, and throbbing, and to be accompanied by abnormal sensitivity to light, noise, and smell [3]. Despite research over the last two decades, the pathophysiology of migraine is still incompletely understood. In this study, we aimed to gather more insights into the mechanisms underlying migraine treatment in a nitroglycerin (NTG)-induced rat model of migraine-like pain [4].
Pituitary adenylate-cyclase-activating polypeptide (PACAP) has emerged as an ubiquitous peptide in primary headache pathophysiology, such as migraine and cluster headache [5]. More than 10 years ago, intravenous administration of PACAP38 in individuals with migraine was discovered to provoke migraine-like attacks [6]. Moreover, PACAP has also been investigated in migraine animal models. In wild-type mice, photophobia and dilation of arteries, such as the middle cerebral and superficial temporal arteries, can be elicited by PACAP38, but not in PACAP-deficient mice [7]. In line with these findings, PACAP may play a mediating role in the initiation and/or promotion of migraine attacks.
Perampanel is a highly selective noncompetitive ionotropic glutamate AMPA receptor antagonist that is presumed to be the first-in-class antiepileptic drug [8]. It has been demonstrated that when the neurons are depolarized, the excitatory impulse can activate the AMPA receptors in the postsynaptic membrane and detach the Mg2+ in the NMDA receptor channel, thereby inducing Ca2+ influx [9]. Based on the existence of common pathophysiological features between epilepsy and migraine [10], it has been recently postulated that antagonists of glutamate receptors may be useful in the treatment of migraine [11]. In fact, the AMPA receptor antagonist perampanel attenuates spontaneous pain-associated behavior and reduces neuropathic and inflammatory pain in rats [12]. In a study by Tringali et al. [13] 100 μM perampanel significantly reduced secretion of calcitonin gene-related peptide (CGRP), the most important peptide neurotransmitter in migraine pathophysiology [14]. Despite this conclusive evidence to support the potential usefulness of perampanel in migraine antagonism, the underlying mechanism driving the effects of perampanel on migraine remains undiscovered. There are still few studies on the pathways that may be involved in this treatment process.
Therefore, the goal of this study was to explore the signaling pathways specifically involved in migraine pathophysiology and to provide new insights for therapeutic targeting of pathways related to migraine development.
Go to : 
MATERIALS AND METHODS
1. Animals
32 Male Sprague-Dawley rats (230–250 g, Pengyue Experimental Animal Breeding Co. Ltd.) were used for this in vivo study. All rats were kept in a 12-hour light-dark environment cycle in a specific pathogen-free animal breeding center, with a humidity of 40% to 70% and a temperature of 20 to 25°C. Food and water were freely available. In order to minimize errors and ensure the reliability of the experimental data, all rats were acclimated in the test environment 1 week before beginning the behavioral experiments. All research behaviors conducted on experimental animals strictly complied with the Institutional Animal Care and Use Committee of Binzhou Medical University Hospital, and the IACUC approval number is 20210802.
2. Experimental groups
The 32 animals were divided into four groups with eight animals in each group: The control group, in which the rats received saline; the NTG (10 mg/kg) group, in which the rats were intraperitoneally injected with NTG at a dose of 10 mg/kg; the NTG + 50 μg/kg perampanel pretreatment group [15], in which the rats were intraperitoneally injected with perampanel at a dose of 50 μg/kg 30 minutes before NTG (10 mg/kg) injection; and the NTG + 100 μg/kg perampanel pretreatment group, in which the rats were intraperitoneally injected with perampanel at a dose of 100 μg/kg 30 minutes before NTG (10 mg/kg) injection.
3. Migraine induction and drug administration
NTG (H11020289, Beijing Yimin Pharmaceutical Co., Ltd.) was diluted in 0.9% saline at 10 mg/kg before the experiment. The rats were treated intraperitoneally with perampanel (#380917-97-5, Shanghai McLean Biochemical Technology) at doses of 50 μg/kg or100 μg/kg 30 minutes before NTG injection. The rats were injected with the drug every other day over a 9-day period (5 total injections). The schedule of drug treatments is reported in Fig. 1. The rats were euthanized following the fifth injection of NTG. The trigeminal ganglion (TG) was dissected for western blot analysis and reverse transcription quantitative polymerase chain reaction (RT-qPCR) analysis, and the serum was used for enzyme-linked immunosorbent assay (ELISA) analysis.
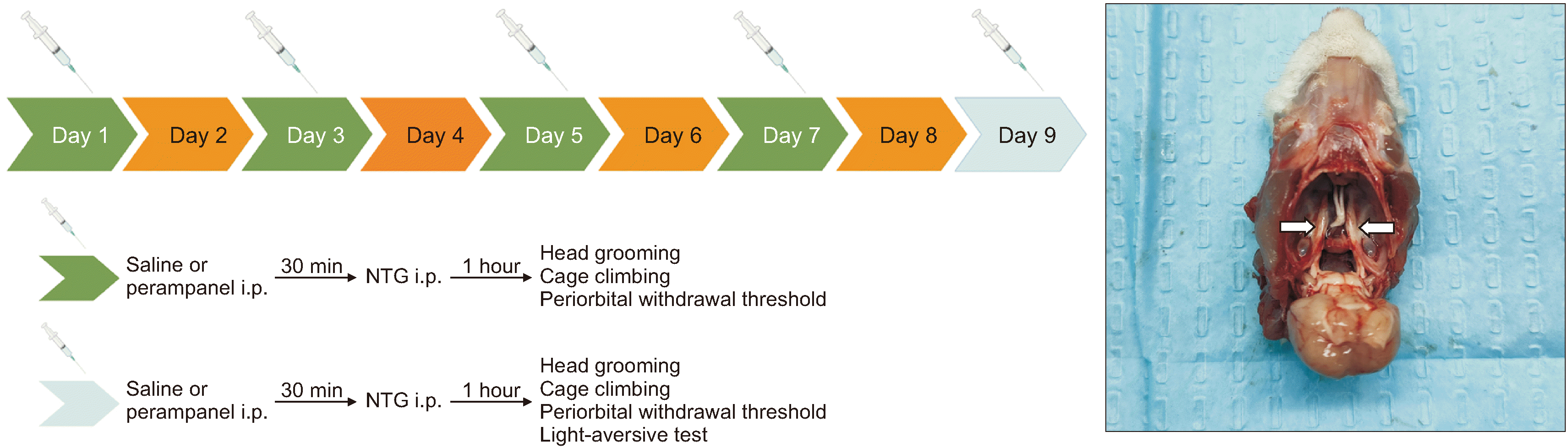
4. Nociceptive behavior test
Head grooming has been defined as a nociceptive behavior, so we observed and recorded these behaviors 30 minutes after the injection of NTG every other day. Head grooming behavior refers to cleaning the head/face with the frontpaws or hind paws. One of the indexes of spontaneous facial pain is to increase the time spent on head grooming [16].We used a video camera to record that. The time it spent grooming its head was calculated.
5. Light-aversive behavior test
One of the characteristics of migraine is photophobia [17]. Therefore, we carried out a light-aversive behavior test 1.5 hours following the injection of drugs on day 9. We used a polypropylene compartment (80 cm length × 40 cm width × 35 cm height) and separated it into two equally sized compartments [18]. There was a gate (12 × 12 cm) opening in the middle. The rats were tested individually in the compartments for 10 minutes where they could move freely. We used a video camera to record the time spent on the dark side, the latency to re-enter the light compartment and the transitions. The light-aversive behavior test was started from the dark side. Latency to re-enter the light compartment means rats were placed individually in the center of the light compartment facing away from the door. The latency for the rats to move to the light side again was counted.
6. Periorbital withdrawal threshold test
A migraine attack is a headache that may be felt anywhere around the head or neck but is typically localized in the frontotemporal and ocular areas. Therefore, we carried out a periorbital withdrawal threshold test to measure mechanical hypersensitivity. The test was conducted every other day 1 hour after drug injection. Researchers restrained each rat with their left hand while the right hand held von Frey filaments and applied them vertically to the midline of the forehead at eye level (periorbital area; innervated by the trigeminal nerve V1 branch) to measure the mechanical hypersensitivity thresholds. The filaments should be held there for 3 to 6 seconds. Asymmetric facial friction, withdrawal, escape, or attack are regarded as a positive reaction. The thresholds were determined using the ‘up-down’ method. In the "up and down" method, the test starts with an estimated monofilament close to the 50% threshold, which causes a positive reaction in 50% of animals. If this monofilament fails to create a response, select a stronger monofilament to continue the test until a positive reaction is obtained. When a positive reaction occurs, the next lower-strength filament is selected for testing. The test continues until at least four readings are obtained after the change of direction and the 50% threshold is calculated. Generally, in the NTG group target force of each von Frey filament floated between 1.2 and 8 g, the control group floated between 2 and 10 g, and the perampanel + NTG group floated between 2 and 26 g, and we didn’t use the cut-off value in this study.
7. Body weight and food consumption measurement
Previous studies have shown that there are tentative links between migraine and eating disorders. In a small clinical study, more than 4 out of 5 female migraine patients reported being on a diet, with over half of patients reporting binge eating and 26% of patients reporting self-induced vomiting [19]. Body weight and food consumption were measured every other day between 10 and 12 am throughout the duration of the study.
8. Western blot
Rats were anesthetized by intraperitoneal injection of chloral hydrate (400 mg/kg) and euthanized, and the brain of each rat was quickly dissected. The individual primary antibodies used in this experiment were rabbit anti-PACAP (1:1,000, ab181205, Abcam), rabbit anti-phospho-CREB (Ser133) (1:1,000, #9198, Cell Signaling Technology), rabbit anti-CREB (1:1,000, #PB0127, Boster), rabbit anti-PKAα/β/γ (1:1,000, YP0226, ImmunoWay), rabbit anti-phospho-PKC (gamma Thr514) (1:1,000, #9379, Cell Signaling Technology), rabbit anti-phospho-PLCγ1 (1:1,000, YP0234, ImmunoWay), anti-β-Actin (1:5,000, #3700, Cell Signaling Technology) and anti-GAPDH (1:5,000, #51332, Cell Signaling Technology). The appropriate secondary antibodies used in this experiment were : IR Dye 800CW Donkey anti-Rabbit (1:5,000, #925-32213, LI-COR Biosciences); IR Dye 680RD Donkey anti-Mouse (1:5,000, #925-68072, LI-COR Biosciences). Images were observed using Odyssey Infrared Imaging System (Li-COR Biosciences).
9. RT-qPCR
We used Tissue RNA Kit (OMEGA, #R6688) to extract Total RNA and Revert Aid First Strand cDNA Synthesis Kit (Thermo ScientificTM, #K1622) to reverse transcribe. Achieved cDNA was used for PCR detection (Applied Biosystems). The following primer pairs specific for the rat GAPDH and PACAP genes were used: GAPDH(F)(5’-CATGTGTAGCGGAGCAAGGTT-3’)/GAPDH(R)(5’-TGATCCCAGGGAAGCTGAGT-3’) and PACAP(F)(5’-ACTCACCTTTGGTCAATCCC-3’)/PACAP(R)(5’-TGATCCCAGGGAAGCTGAGT-3’). The reaction conditions were as follows: 95°C for 15 minutes; 40 cycles of 94°C for 15 seconds, 55°C for 30 seconds and 72°C for 45 seconds; extentioning at 72 °C for 10 minutes. Compared with the control sample, the 2−ΔΔCT method was performed to obtain the relative fold change of the PACAP normalized by the GAPDH [20].
10. ELISA
The levels of PACAP in the serum were detected by ELISA using the rat PACAP ELISA kit (Shanghai Enzyme Link Biotechnology). Absorbance was measured in the Multiskan Go Microplate Spectrophotometer (Molecular Devices).
11. HT-22 cells
HT-22 cells (ZQ0476, Shanghai Zhongqiao Xinzhou Biotechnology Co., Ltd) were used for the in vitro study. The cells were placed in Dulbecco’s modified Eagle’s medium containing 10% fetal bovine serum, 100 μ/mL penicillin and 100 ng/mL streptomycin (HY-B0472, MCE) and maintained at 37°C under 5% CO2. The cells were seeded in 6-well plates and incubated with different drugs treatment: 100 µM perampanel [21], 100 μM SQ22536 (HY-100396, MedChemExpress), a adenylate cyclase (AC) inhibitor; 10 µM forskolin (HY-15371, MedChemExpress), a AC agonist; 10 µM H-89 (HY-15979, MedChemExpress), a protein kinase A (PKA) inhibitor; 0.1 mM 8-bromo-cAMP sodium salt (HY-12306, MedChemExpress), a PKA agonist and 25 µM KG501 (HY-103299, MedChemExpress), and a cAMP-responsive-element-binding protein (CREB) inhibitor for 24 hours. Cell lysates were collected to do western blot analysis 24 hours later.
12. Statistical analysis
All data in the study were calculated using GraphPad Prism 7.0 (GraphPad Software). All data were presented in the form of mean ± standard error of mean. Measurement data were analyzed using the Kruskal–Wallis test and two-way repeated measures analysis of variances, P < 0.05 was considered a significant difference.
Go to :
RESULTS
1. Perampanel inhibited NTG-induced photophobia, migraine-like pain and nociceptive behaviors
An NTG-induced migraine model was used to investigate whether perampanel could ameliorate the migraine-like pain. As shown in Fig. 2, our results showed that pretreatment with perampanel not only significantly increased the time rats spent in the light compartment in the light-aversive behavior test but also increased the periorbital withdrawal thresholds. Compared with the NTG group, the perampanel + NTG group exhibited a significantly longer time spent in the light compartment, a shorter latency to re-enter the light compartment, and more transitions (***P < 0.001, Fig. 2). We also investigated head grooming as a nociceptive behavior in conscious rats. As shown in Fig. 3A, after days of injection, the head grooming time of the rats treated with NTG was significantly longer than that of the rats which were also treated with perampanel at doses of 50 and 100 μg/kg on day 7 (###P = 0.001 and $$P = 0.004 respectively, Fig. 3A). After 5 injections, compared with the NTG group, the 100 μg/kg perampanel + NTG group spent a shorter time on head grooming ($$$P = 0.001, Fig. 3A). The periorbital withdrawal thresholds in the 50 μg/kg and 100 μg/kg perampanel + NTG group were higher than those in the NTG group beginning on day 3 (#P = 0.014 and $P = 0.016 respectively, Fig. 3B). After being administered 4 times, the thresholds in the NTG group were significantly lower than those in the 100 μg/kg perampanel + NTG group ($P = 0.020, Fig. 3B). In this study, the relationships between migraine and eating disorders were also tentatively explored. After 5 injections of NTG, the rats from the different groups showed no significant differences in body weight (P = 0.793, Fig. 4A) or food consumption (P = 0.091, Fig. 4B). Collectively, these results suggested that perampanel can inhibit photophobia, migraine-like pain, and nociceptive behaviors in NTG-induced migraine rats.
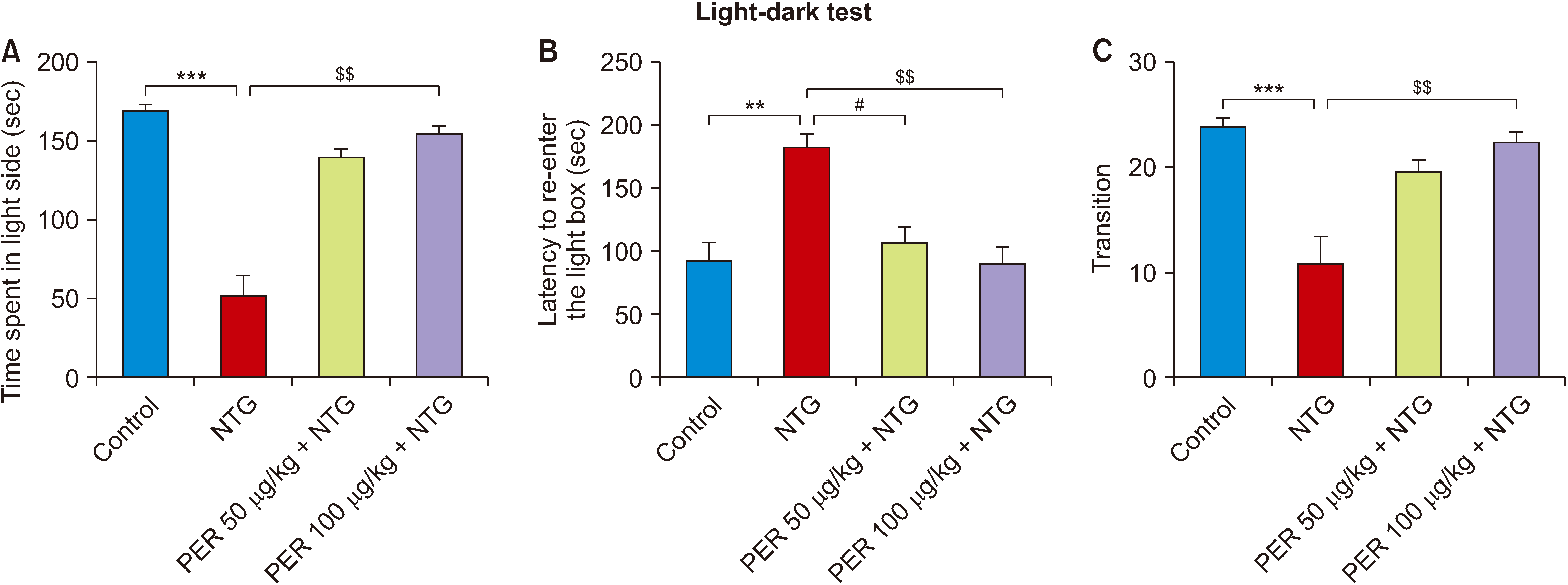 | Fig. 2Effects of perampanel treatment on photophobia. (A) Time spent in the light compartment, Kruskal–Wallis statistic = 24.30, ***P < 0.001; (B) Latency to re-enter the light compartment, Kruskal–Wallis statistic = 16.33, ***P = 0.001; (C) Transition, Kruskal–Wallis statistic = 20.55, ***P < 0.001. Control, n = 8; NTG, n = 8; PER 50 μg/kg + NTG, n = 8; PER 100 μg/kg + NTG, n = 8. Data are presented as means ± SEM. NTG: nitroglycerin, PER: perampanel. $$P < 0.01 vs. NTG group; **P < 0.01, ***P < 0.001 vs. control group; #P < 0.05 vs. NTG group in corresponding days.
|
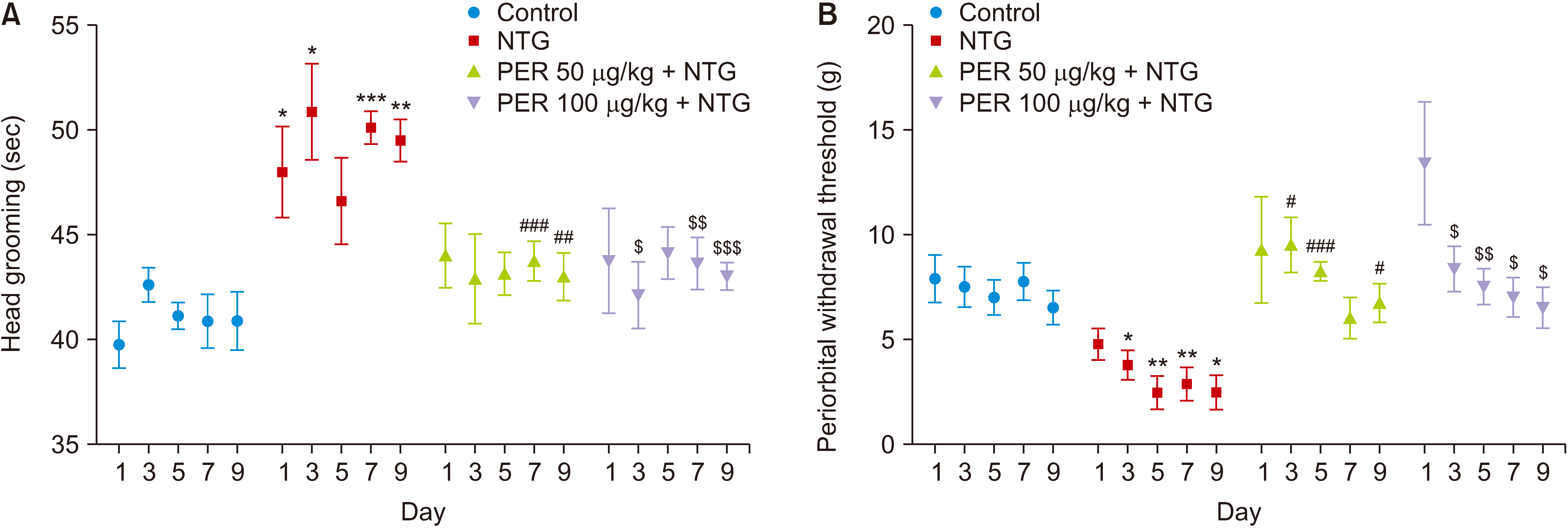 | Fig. 3Effects of perampanel treatment on NTG-induced migraine-like pain and nociceptive behaviors. (A) Head grooming, subgroups, F (3, 28) = 25.95, P < 0.001; time, F (4, 112) = 0.295, P = 0.880; subgroups × time, F (12, 112) = 0.629, P = 0.814. Control, n = 8; NTG, n = 8; PER 50 μg/kg + NTG, n = 8; PER 100 μg/kg + NTG, n = 8. In periorbital withdrawal thresholds test, NTG were decreased periorbital withdrawal thresholds, while perampanel treatment, at both doses of 50 and 100 μg/kg significantly increased the thresholds. (B) Periorbital withdrawal thresholds, subgroups, F (3, 28) = 23.28, P < 0.001; time, F (4, 112) = 4.397, P = 0.002; subgroups × time, F (12, 112) = 0.9606, P = 0.491. Data are presented as means ± SEM. NTG: nitroglycerin, PER: perampanel. $P < 0.05, $$P < 0.01, $$$P < 0.001 vs. NTG group; *P < 0.05, **P < 0.01, ***P < 0.001 vs. control group; #P < 0.05, ##P < 0.01, ###P < 0.001 vs. NTG group in corresponding days.
|
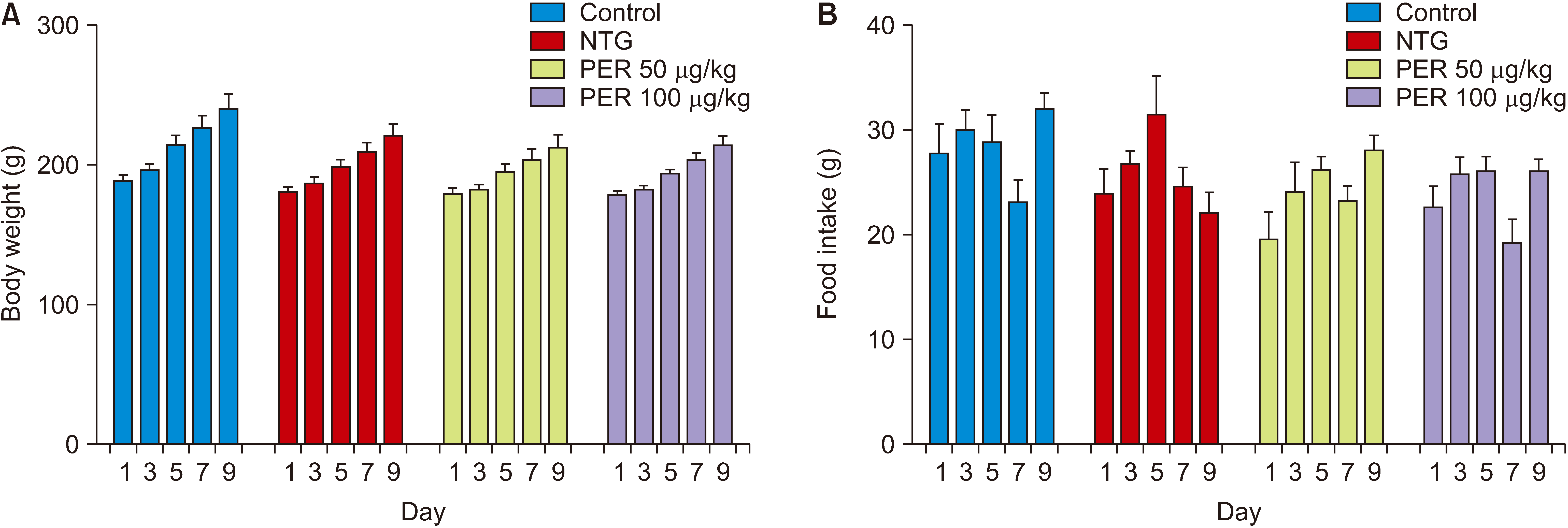 | Fig. 4Effects of perampanel treatment on eating disorders. (A) Body weight, subgroups, F (3, 28) = 4.720, P = 0.009; time, F (4, 112) = 67.16, P < 0.001; subgroups × time, F (12, 112) = 0.652, P = 0.793; (B) Food intake, subgroups, F (3, 28) = 4.311, P = 0.013; time, F (4, 112) = 5.916, P < 0.001; subgroups × time, F (12, 112) = 1.637, P = 0.091. Control, n = 8; NTG, n = 8; PER 50 μg/kg + NTG, n = 8; PER 100 μg/kg + NTG, n = 8. Data are presented as means ± SEM. NTG: nitroglycerin, PER: perampanel.
|
2. Perampanel inhibited the increase in PACAP level
As shown in Fig. 5, the expression of PACAP was increased by NTG injection, but pre-administration of perampanel suppressed the increase. In the western blot assay, PACAP expression was found to be upregulated in the NTG group compared to the control group (*P = 0.041, Fig. 5A, B); however, pretreatment with perampanel at doses of 50 μg/kg and 100 μg/kg downregulated the expression of PACAP (*P = 0.046, *P = 0.017 respectively, Fig. 5A, B). The RT-qPCR analysis revealed that the mRNA expression of PACAP was upregulated in the NTG group compared to the control group (**P = 0.001, Fig. 5C). Consistent with the results of western blot and RT-qPCR analysis, the ELISA results revealed an increased trend for the NTG group (*P = 0.020, Fig. 5D). Collectively, these results suggested that PACAP level may be correlated with migraine-like behavior, and that increased PACAP level may play an important role in the pathogenesis of the pain caused by migraine, suggesting that perampanel may relieve migraine through inhibiting the increase in PACAP in NTG-induced migraine rats.
 | Fig. 5Effects of perampanel treatment on PACAP level in NTG-induced migraine. (A) Western blot showing the protein levels of PACAP; (B) Western bolt analysis of PACAP expression in TG. Kruskal–Wallis statistic = 12.15, **P = 0.007; (C) qRT-PCR analysis of PACAP gene expression in TG. Kruskal–Wallis statistic = 14.16, **P = 0.003, P < 0.001; (D) Elisa analysis of PACAP level in serum. Kruskal–Wallis statistic = 9.828, *P = 0.020. Control, n = 4; NTG, n = 6; PER 50 μg/kg + NTG, n = 6; PER 100 μg/kg + NTG, n = 6. Data are presented as mean ± SEM. PACAP: pituitary adenylate-cyclase-activating polypeptide, NTG: nitroglycerin, TG: trigeminal ganglion, PER: perampanel. Significant differences: *P < 0.05, ***P < 0.001.
|
3. Perampanel affected the PKA/CREB signaling pathways in NTG-induced migraine
Then, we examined the expression levels of the cAMP/PKA/CREB pathway in the TG. The western blot results showed that the protein levels of PKA and p-CREB of the perampanel rats in preteatment were lower than those in NTG-induced rats (**P = 0.002, **P = 0.006 respectively, Fig. 6A–D). Our results showed that perampanel could inhibit the cAMP/PKA/CREB pathway, suggesting that perampanel may regulate the expression of PACAP through the cAMP/PKA/CREB pathway. This confirms previous research [22].
 | Fig. 6Effects of perampanel on PKA/CREB and PLC/PKC signaling pathways in NTG-induced migraine. (A) Western blot showing the protein levels of PKA, CREB and p-CREB; (B) Western blot analysis of the expression of PKA in TG. Kruskal–Wallis statistic = 17.20, ***P < 0.001; (C) Western blot analysis of the expression of P-CREB in TG. Kruskal–Wallis statistic = 11.52, **P = 0.009; (D) Western blot analysis of the expression of CREB in TG. Kruskal–Wallis statistic = 9.125, *P = 0.028; (E) Western blot showing the protein levels of PLC and PKC. (F) Western blot analysis of the expression of PLC in TG. Kruskal–Wallis statistic = 3.631, P = 0.304; (G) Western blot analysis of the expression of PKC in TG. Kruskal–Wallis statistic = 7.030, P = 0.071. Control, n = 4; NTG, n = 6; PER 50 μg/kg + NTG, n = 6; PER 100 μg/kg + NTG, n = 6. Data are presented as mean ± SEM. PKA: protein kinase A, CREB: cAMP-responsive-element-binding protein, PLC: phospholipase C, PKC: protein kinase C, NTG: nitroglycerin, TG: trigeminal ganglion, PER: perampanel. Significant differences: *P < 0.05, **P < 0.01.
|
To gain additional insights into the mechanism by which NTG induces PACAP synthesis in the TG, we examined the phospholipase C (PLC)/protein kinase C (PKC) pathway. However, no significant differences in the protein expression of PLC and PKC were observed between the control group and the NTG group (P = 0.304 and P = 0.071 respectively, Fig. 6E–G). These results showed that the PLC/PKC pathway may not be involved in NTG-mediated induction of PACAP.
4. Perampanel decreased the expression of PACAP via inhibition of the cAMP/PKA/CREB pathway in vitro
Forskolin treatment had no effect on PACAP expression in cells (P = 0.982, Fig. 7A, B), while 8-bromo-cAMP sodium salt significantly increased PACAP expression compared with that in the perampanel group (**P = 0.007, Fig. 7C, D). Furthermore, perampanel decreased the expression of PACAP compared with the control group in this vitro study. We further assessed the effect of the PKA inhibitor H-89 (10 μM), the AC inhibitor SQ22536 (100 μM), and the CREB inhibitor KG501 (25 μM). The expression of PACAP in the groups treated with the H-89 and the SQ22536 was notably decreased compared with that in the control group (**P = 0.007 and **P = 0.0051, respectively, Fig. 7E–H). The CREB inhibitor also decreased PACAP expression (**P = 0.010, Fig. 7I, J). Taken together, these results suggested that 100 μM perampanel may exert an effect on reducing the expression of PACAP through downregulating cAMP/PKA/CREB signaling.
 | Fig. 7Effects of perampanel on cAMP/PKA/CREB signaling pathways in vitro. (A) Western blot showing the protein levels of PACAP in HT-22 cell incubated with forskolin 10 µM for 24 hours; (B) Western blot analysis of the expression of PACAP. Kruskal–Wallis statistic = 7.538, *P = 0.011. Control, n = 4; Forskolin 10 µM, n = 4; PER 100 μM, n = 4; (C) Western blot showing the protein levels of PACAP in HT-22 cell incubated with 8-bromo-cAMP sodium salt 0.1 mM for 24 hours; (D) Western blot analysis of the expression of PACAP. Kruskal–Wallis statistic = 16.33, P = 0.001. Control, n = 4; 8-bromo-cAMP sodium salt 0.1 mM, n = 4; PER 100 μM, n = 4; (E) Western blot showing the protein levels of PACAP in HT-22 cell incubated with SQ22536 100 μM for 24 hours; (F) Western blot analysis of the expression of PACAP. Kruskal–Wallis statistic = 9.269, ***P < 0.001. Control, n = 4; SQ22536 100 μM, n = 4; PER 100 μM, n = 4. (G) Western blot showing the protein levels of PACAP in HT-22 cell incubated with H-89 10 µM for 24 hours; (H) Western blot analysis of the expression of PACAP. Kruskal–Wallis statistic = 9.846, ***P < 0.001. Control, n = 4; H-89 10 µM, n = 4; PER 100 μM, n = 4. (I) Western blot showing the protein levels of PACAP in HT-22 cell incubated with KG501 25 µM for 24 hours; (J) Western blot analysis of the expression of PACAP. Kruskal–Wallis statistic = 8.769, **P = 0.001. Control, n = 4; KG501 25 µM, n = 4; PER 100 μM, n = 4. Data are presented as mean ± SEM. PKA: protein kinase A, CREB: cAMP-responsive-element-binding protein, PER: perampanel. Significant differences: *P < 0.05, **P < 0.01.
|
Go to :
DISCUSSION
Migraine, one of the most common neurological diseases, contributes to significant disability and reduced quality of life [23]. The pathophysiology of migraine is not completely clear, and there is no specific cure. Therefore, medical approaches should be individualized to adequately treat migraine. Understanding of the underlying mechanism of migraine has evolved to recognize a second messenger stage. Previous studies have shown that some key signaling pathways are dysregulated in migraine [24]. However, research on signal transduction pathways in the pathogenesis of migraine is incomplete. The activity of signal transduction pathways is difficult to explain, but further in-depth research may provide useful information on the roles of signal transduction pathways in migraine. The signals that ultimately lead to migraine are transmitted to the ventroposterior thalamus from the TG [25]. Therefore, in the present study, the TG was examined to reveal the pathophysiological mechanism of migraine. In the cascade of events controlling vascular function, second order neurons also participate in the release of several neurotransmitters [26]. For example, release of PACAP, a well-known vasodilator, leads to subsequent increases in vessel diameter and blood flow in the meninges and cortex, both of which can further activate vascular and meningeal nociceptors, leading to migraine headache [27].
It has been demonstrated that the AMPA receptor plays critical roles in receptor trafficking and neuronal plasticity [18]. Moreover, migraine and epilepsy are linked by their symptom profiles, comorbidities and treatments. Therefore, the AMPA receptor inhibitor perampanel, presumed to be the first-in-class antiepileptic drug, may have antimigraine effects. In our study, the antimigraine effects of perampanel were observed with nociceptive behavior, periorbital withdrawal threshold tests, and light-aversive behaviors, which are standard behavioral tests for analgesia assessment. Our results confirmed the antimigraine effects of perampanel, which significantly attenuated the spontaneous nociception caused by NTG administration. Moreover, perampanel was able to counteract peripheral pain, as indicated by a reduction of periorbital mechanical hypersensitivity. Another feature of migraine is photophobia, in fact, exposure to light increases the severity of migraine. Nearly 90% of migraine patients with normal vision report that they experience photophobia because that the light signal from the eye converges on the trigeminovascular neurons somewhere in its path. In our study, we found that NTG-induced photophobia in rats was attenuated by perampanel pretreatment. In addition, there is a bidirectional relationship between migraine and eating disorders. Patients with eating disorders have a greater risk of developing migraine than patients without eating disorders [28]. Similarly, patients with migraine have a higher risk of developing eating disorders than patients without migraine [29]. Therefore, we preliminarily explored the ability of perampanel treatment to reduce the rates of migraine and comorbidities. We measured body weight because eating disorders can have many effects, including decreased body weight. However, in this study, a notable difference between the perampanel and NTG groups was not observed after five injections of NTG. A possible explanation is that eating disorders require a relatively long period to develop.
PACAP and its related receptors are widely distributed in the nervous system, including several areas where pain transmission is related to migraine. But its role in migraine and its pathophysiology is not fully understood. In the present study, PACAP was upregulated in the TG in rats with NTG-induced migraine compared with control rats, as evidenced by RT-qPCR, western blot, and ELISA results. PACAP is expressed in the TG and is released into the cerebrospinal fluid and blood after trigeminal stimulation and nociceptive input stimulation. We observed by ELISA that PACAP levels in serum were markedly higher in NTG-induced migraine model rats than in control rats.
Moreover, as the authors’ in vitro research suggests, for the first time, perampanel can regulate PACAP expression by inhibiting cAMP/PKA/CREB signaling. Our results showed that perampanel downregulated PACAP levels but that inhibition of PACAP with forskolin and 8-bromo-cAMP sodium salt attenuated this effect. Activated PKA phosphorylates and catalyzes the phosphorylation of regulatory proteins, causing a series of downstream changes. Phosphorylated CREB is a key transcriptional cofactor that initiates not only biological processes but also activates ion channels and cellular motor proteins, as well as regulating metabolic enzyme activity [30]. These phosphorylation processes interweave the cAMP/PKA/CREB signal transduction pathway and other signal cascades to form a complex signal network, and provide multiple feedback loops to further regulate cAMP activity, indicating that the cAMP/PKA pathway plays a key role in inducing and maintaining synaptic plasticity in the nervous system [31]. Behavioral analyses of mechanical allodynia have revealed recovery after cAMP/PKA/CREB signaling is inhibited with a PKA inhibitor, suggesting that the effects of the PKA inhibitor may occur through regulation of mechanical allodynia [32]. Interestingly, the PKA inhibitors H-89 and PKI (14-22) can block the regulation of CGRP release and regulate pain sensitization [33].
These signal transduction pathways are complex and not linear, and in some cases of migraine, crosstalk occurs between and among various pathways. In the present study, we attempted to describe the signal transduction pathways that may be involved in the pathophysiology of migraine. Luckily, we found, for the first time, that the AMPA receptor inhibitor perampanel inhibits NTG-induced migraine-like pain and that the cAMP/PKA/CREB pathway may contribute to the pathogenesis of migraine, which potentially could be important to the pathophysiology of migraine.
In summary, our study clearly showed the potential of perampanel as a therapy to prevent migraine attack. In an NTG-induced migraine model, preadministration of perampanel alleviated migraine-like pain. In an in vitro study, perampanel reduced the expression of PACAP through inhibition of the cAMP/PKA/CREB pathway. However, the exact mechanism by which perampanel acts on the cAMP/PKA/CREB pathway is complex and remains unclear. Defining the protective effects of perampanel against diseases such as migraine will provide more insights into the pathogeneses of nervous system disorders and support the development of therapeutic strategies for these disorders.
Go to :
 XML Download
XML Download