

 PDF
PDF Citation
Citation Print
Print



Introduction
The classical descriptive study by Schunke [1] characterized the morphology of the fetal sacroiliac joint (SIJ) during development as an unequal development of the two hard tissues, a bony ilium and a cartilaginous sacrum; followed by formation of interarticular septa with synovial folds after 20 weeks gestational age (GA); and then a very slow and incomplete cavitation, with a thin periosteum interposed at full term. Based on these specific features, Vleeming et al. [2] hypothesized a similarity between the SIJ and the temporomandibular joint (TMJ). The TMJ contains a definite disk and, in contrast to the early ossification of the temporal bone, the mandibular condyle cartilage forms later in a specific manner [3-6].
Schunke [1] also reported significant inter-individual variations in the advancement of joint cavitation in the SIJ, and this started as early as 10 weeks GA in one case. However, other studies estimated that the GA when joint cavitation occurred varied by 3 to 4 weeks [1, 7-10]. Thus, there appear to be significant inter-individual variations in the timing and sequence of development and growth of the SIJ. The adult SIJ has two specific morphologies. In one morphology, the posterior capsule is rudimentary or absent, in contrast to the anterior capsule that is reinforced by the anterior sacroiliac ligament [11]; in the other morphology, the so-called “fibrous ankylosis”, the intra-articular fibrocartilage bands connect between the ilium and sacrum [12, 13].
Consequently, despite many studies of this topic, there appears to be no uniform timing and sequence in morphogenesis of the SIJ, and individual variations may mask underlying similarities. To determine the common features of SIJ morphogenesis, we aimed to clarify the timing and sequence of three key developmental events: joint cartilage development, cavitation, and advancing ossification of the ilium. We also compared SIJ morphology with the morphology of the humeroradial joint of two fetuses and with the TMJ of five fetuses. Like the TMJ, the humeroradial joint has a thick and tight synovial fold, even in fetuses [14].
Go to : 
Materials and Methods
The study was performed in accordance with the provisions of the Declaration of Helsinki, as revised in 2013. Paraffin-embedded histological sections of the pelvis were obtained from 24 human fetuses at GAs of approximately 10 to 30 weeks (crown-rump length [CRL]: 49–250 mm). These specimens were from three different institutions, and each institution provided independent approval.
All 13 specimens smaller than 104 mm CRL (Table 1) were part of the large collection at the Institute of Embryology, Universidad Complutense Madrid. These specimens were the products of miscarriages or ectopic pregnancies and are managed at its Department of Obstetrics. While in Madrid, these sections were stained with hematoxylin and eosin (H&E) or azan. All sections were serial and the sectional planes were horizontal. The use of the sections in Madrid was approved by the local university ethics committee (No. B08/374).
Table 1
Developmental sequence among the sacroiliac joint cartilage, the cavitation and the advancing ossification of the ilium
| Size of fetus | Joint cartilage | Cavitation | Ossification of ilium |
|---|---|---|---|
| 49 mm CRL | None | None | Anterior side of the future joint |
| 52 mm (Fig. 1A) | None | None | Anterior side of the future joint |
| 56 mm | None | None | Anterior side of the future joint |
| 62 mm | None | None | Anterior side of the future joint |
| 76 mm (Fig. 1B) | None | None | Reached the future jointb |
| 80 mm | None | None | Reached the future joint |
| 82 mm (Fig. 1C) | Small | None | Reached the future jointb |
| 85 mm | None | None | Reached the future joint |
| 90 mm (Fig. 1D) | Extended anterior 1/3a | None | Covered the future joint |
| 92 mm | Small | None | Covered the future joint |
| 97 mm | Extended anterior 1/3 | None | Covered the future joint |
| 97 mm-2 | Small | None | Covered the future joint |
| 103 mm | Extended anterior 1/3 | None | Covered the future joint |
| 107 mm | Small | None | Covered the future joint |
| 115 mm | Extended anterior 1/3 | None | Covered the future joint |
| 120 mm (Fig. 2A) | Extended anterior 1/4 | None | Covered the future joint |
| 145 mm (Fig. 2B) | Extended almost halfa | Present | Extended over the joint |
| 150 mm | Extended almost half | Present | Extended over the joint |
| 155 mm (Fig. 2C) | Extended almost half | None | Covered the future joint |
| 160 mm | Extended almost half | None | Covered the future joint |
| 165 mm | Extended anterior 2/3 | Present | Covered the future joint |
| 210 mm | Fully extended | Present | Extended over the joint |
| 232 mm (Fig. 2D) | Fully extended | Present | Extended over the joint |
| 250 mm | Fully extended | Present | Extended over the joint |
aThe joint cartilage of the ilium extended along the bony ilium from the anterior end of the future joint area and covered the anterior 1/3 (or the anterior half) of the area. bOssification of the ilium started at the far anterior side of the future joint, extended posteriorly to reach the future joint area and, further extended over the area.
![]()
Unilateral pelvises of 8 larger fetuses (>119 mm CRL, and one with a CRL of 155 mm; Table 1) were prepared as paraffin-embedded sections at 100-µm intervals. The sectional planes were horizontal, and the sections were stained with H&E. These specimens were in the collection of the Department of Anatomy, Akita University, Japan. All specimens were donated by their families to the Department from 1975 to 1985 and preserved in a 10% (w/w) neutral formalin solution for more than 30 years. The date of donation and GA of each fetus were available, but there was no information on family name, name of the obstetrician or hospital, or reason for the abortion. The use of these specimens for research was approved by the local ethics committee (No. 1428).
Sagittal sections (H&E staining) that included the TMJ were prepared from the heads of 5 of the 21 specimens described above (CRL 92, 120, 150, 210 and 250 mm; approximately 15–30 weeks GA). These sagittal sections were also used in our recent studies [15, 16]. Among these 5 specimens, sagittal sections of the humeroradial joint were examined in 2 fetuses (CRL 210 and 250 mm), sections that were used in our previous study of muscles covering the elbow joints [17].
The other three fetuses (CRL 107, 115, and 155 mm; approximately 14, 15, and 18 weeks GA) were donated by their families to the Department of Anatomy, Yanbian University Medical College, Yanji, China. Their use for research was approved by the local university ethics committee (No. BS-13-35) while the second author was on its faculty. These fetuses were obtained by induced abortion, after which the obstetrician at the college teaching hospital orally informed the mother that the fetus could be donated for research, but made no attempt to actively encourage donation. After a mother agreed, the fetus was assigned a specimen number and stored in a 10% w/w neutral formalin solution for more than 1 month. Because of specimen number randomization, there was no possibility of contacting the family at later time.
After dividing the body into parts, pelvic samples were decalcified by incubation at 4°C in a 0.5 M EDTA solution (pH 7.5; Decalcifying Solution B; Wako) for 3 to 5 days, depending on sample size. Most sections were stained with H&E or Masson’s trichrome. Some samples were used for immunohistochemical analysis of osteoclasts and macrophages using the mouse monoclonal anti-human CD68 KP1 (Dako M0814, 1:100). After washing, the sections were incubated with the secondary antibody (incubation for 30 minutes; dilution 1:1,000; Histofine Simple Stain Max-PO, Nichirei), labeled with horseradish peroxidase (HRP), and antigen-antibody reactions were detected by the HRP-catalyzed reaction with diaminobenzidine (incubation for 3–5 min; Histofine Simple Stain DAB, Nichirei). All samples were counterstained with hematoxylin. Each experiment included a negative control (tissue specimens without a primary antibody).
Sections were observed and photographed using a Nikon Eclipse 80 microscope. However, photographs at ultra-low magnification (objective lens <1×) were taken using a high-grade flat scanner (Epson GTX970) with translucent illumination.
Go to :
Results
Observations of the SIJ
We examined 24 specimens whose GA ranged from about 10 to 30 weeks. We found that the entire sacrum was cartilaginous, but the ilium consisted of regions with different levels of ossification. The membranous ossification of the ilium started in the lateral and medial aspects, near the anterior end of the bone at 10 weeks GA (Fig. 1A). The endochondral ossification started in the anterolateral side of the future SIJ at 12 weeks GA (Fig. 1B). Both ossifications extended posteriorly and reached the future joint area at 12 weeks GA (Fig. 1B, C), and extended over the joint at 15 weeks GA (Fig. 2A). A small mass of the joint cartilage appeared at the anterior end of the future SIJ at 12 weeks GA (Fig. 1C). The cartilage tissue extended posteriorly along the bony ilium, reached the anterior half of the joint area at 18 weeks GA (Figs. 1D, 2A–C), and was covered at 26 weeks GA (Fig. 2D). Notably, the joint cartilage of the ilium was always distant from and anterior to the hypertrophied cartilage tissues at the ossification center of the ilium.
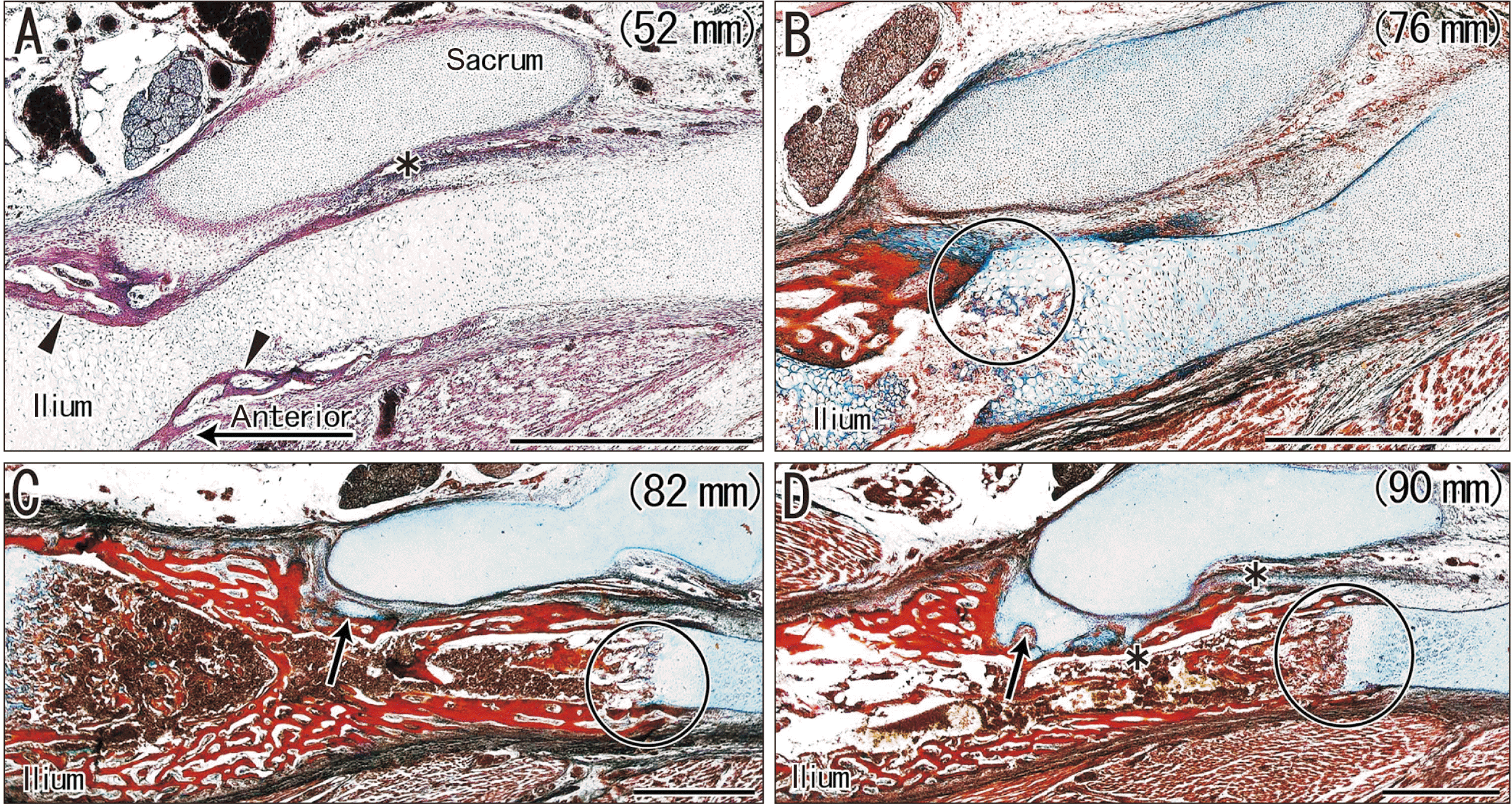 | Fig. 1Horizontal sections of sacroiliac joints in four fetuses (crown-rump length: 52–90 mm) that were stained with H&E (A) or azan (B–D). (A, B) show that ossification of the ilium occurs in the anterior part, but does not reach the future joint area. (A) shows membranous bones (arrowheads), not cartilaginous bones. (C) shows a small cartilage mass attached to the ilium (arrow). (D) shows that the cartilage covers almost one-third of the future joint area (arrow). The circles in (B–D) indicate endochondral ossification of the ilium. The asterisks in (A, D) indicate an artifact space produced during histological procedure. (A–D) Scale bars=1 mm.
|
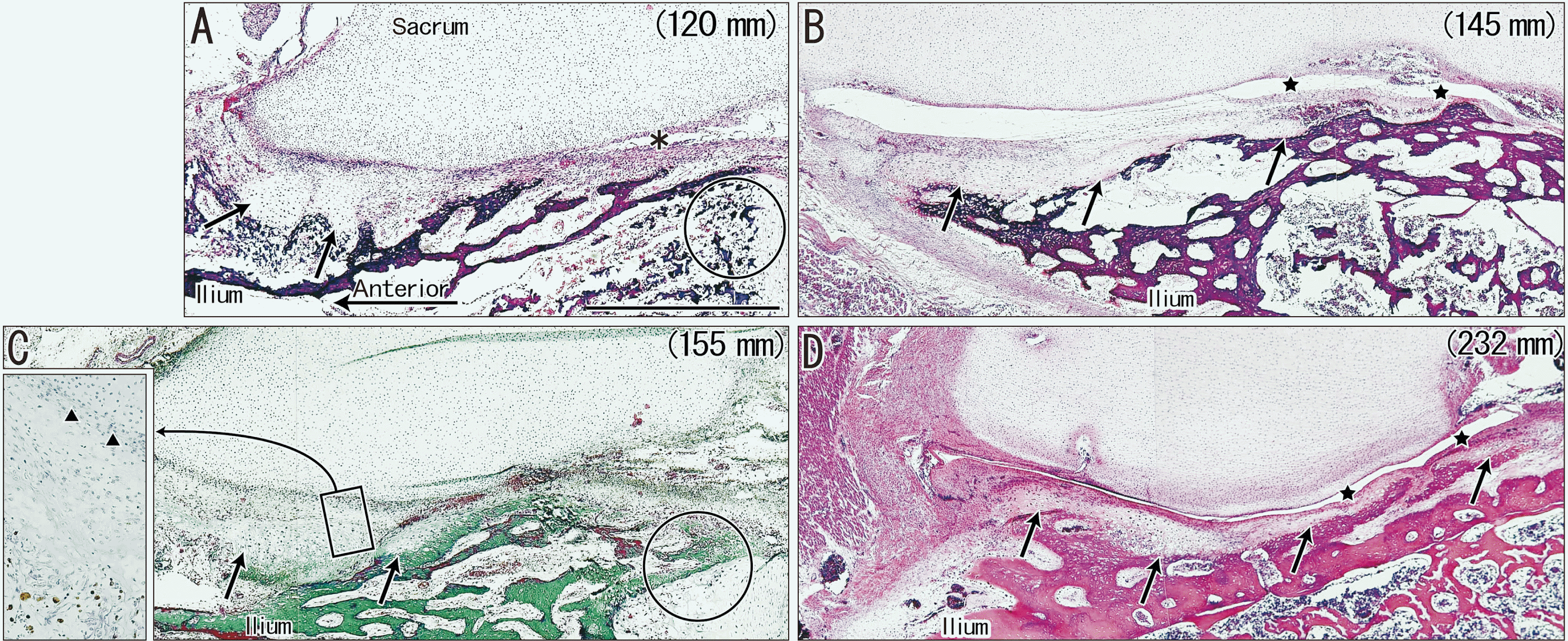 | Fig. 2Horizontal sections of sacroiliac joints in four fetuses (crown-rump length: 120–232 mm) that were stained with H&E (A, B, D) or Masson’s trichrome (C). All panels show the bony ilium covering the joint area. (A–C) show that the joint cartilage of the ilium (arrows) covers the anterior half of the joint area. (D) shows the cartilage extends posteriorly over the joint. (B, D) show the joint cavities (stars). The insert on the left of (C) (corresponding to the rectangle in the main panel) shows CD68-positive cells at higher magnification, in which the future joint space (triangles) contains no macrophages. The circles in (A, C) indicate endochondral ossification of the ilium. The asterisk in (A) indicates an artifact space produced during the histological procedure. All panels (except the C insert) were at the same magnification. (A) Scale bar=1 mm.
|
One specimen had evidence of cavitation at 15 weeks GA (Fig. 2B), but two other specimens had no cavitation at 18 weeks GA (Fig. 2C). After 19 weeks GA, all specimens had joint cavities (Fig. 2D). During development, the joint cavity (if present) contained no septum or disk-like structure. Immunostaining of CD68 showed clear evidence of osteoclasts in the bony ilium, but no macrophages in the future joint or in the interzone between the sacrum and ilium (insert of Fig. 2C). Table 1 summarizes our observations of these specimens. In the ilium, ossification and joint cartilage growth advanced from the anterior to the posterior site. Notably, the posterior extension of the cartilage along the medial aspect of the ilium followed the advancing ossification.
The ilium was covered by the gluteus and back muscles from the superior, posterior and/or lateral sides as well as the iliacus muscle from the anteromedial side (details, described in our recent publications [18, 19]). The iliacus muscle appeared to be thicker at the more anterior origin from the iliacus at any stages examined. The myofibers were composed of the secondary myotube in fetuses larger than 80 mm CRL (figures, not shown).
Observations of the TMJ and humeroradial joint
Our observations of the TMJ in five specimens (CRL 92, 120, 150, 210, and 250 mm; approximately 15–30 weeks GA) indicated the temporal bone was bony near the mandibular condyle (Fig. 3). To facilitate comparison of the cartilaginous condyle with the sacrum, we presented these panels up-side-down, so the condyle is on the top of each panel, even in sagittal view. The cavitation appeared to start at 15 to 16 weeks GA (Fig. 3A).
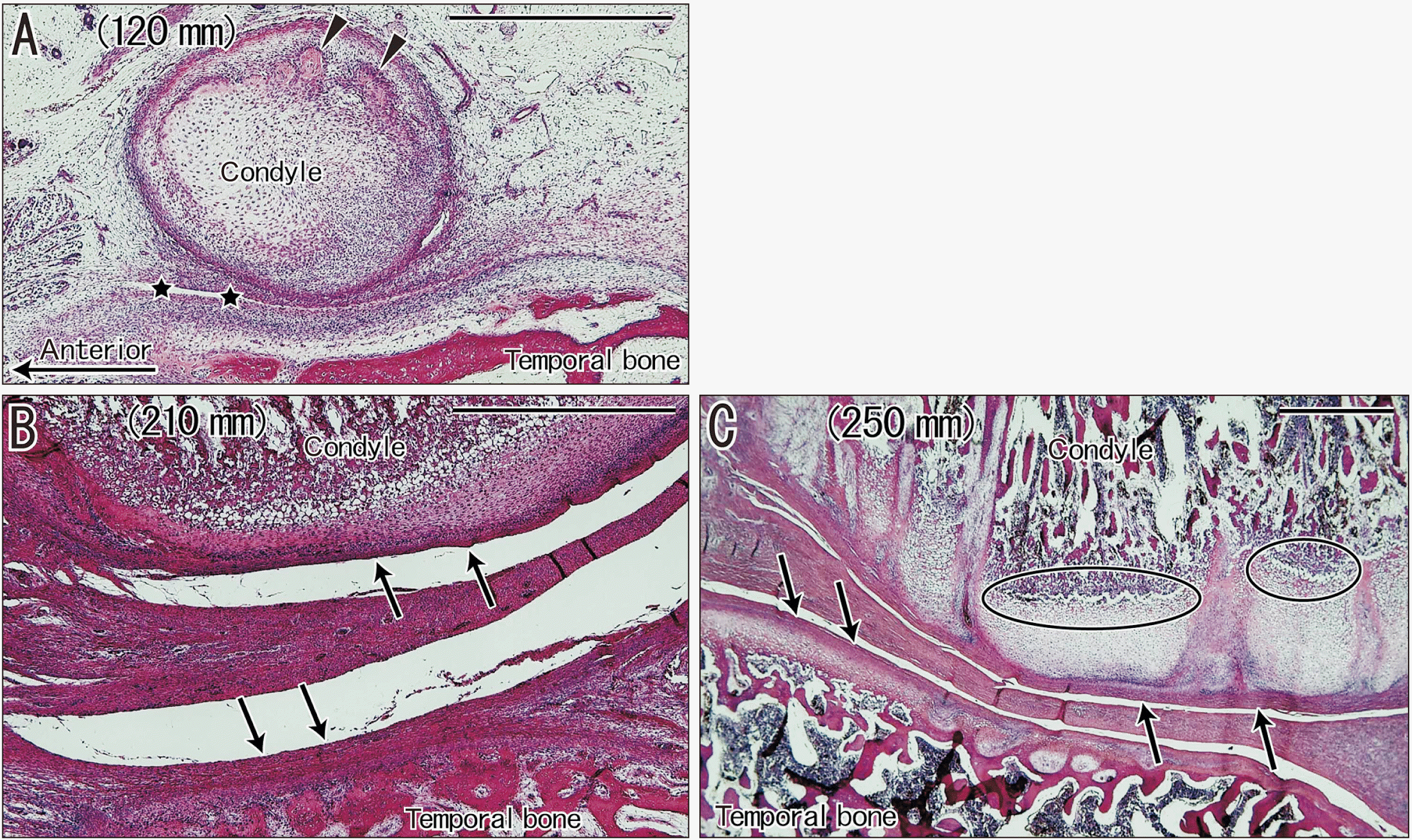 | Fig. 3Sagittal sections of temporomandibular joints in three fetuses (crown- rump length: 125, 210, and 250 mm) that were stained with H&E. (A) shows attachment of the condylar cartilage to the bony temporal bone, with initiation of ossification in the condyle (arrowheads) and cavitation (stars). (B, C) show that the joint cavity is large and the disk is thick, but a periosteum-like membrane (arrows) covers the joint surface of the condyle and temporal bone, instead of the joint cartilage. Ovals in (C) show endochondral ossification occurs in the condyle. (A–C) Scale bars=1 mm.
|
There was evidence of several foci of ossification in the condyle in two of the five specimens (Fig. 3A; CRL: 120 and 150 mm; approximately 15–18 weeks GA). A joint disk was not evident in the smallest specimen (Fig. 3A). The other two larger fetuses (CRL: 210 and 250 mm; Fig. 3B, C) had advanced endochondral ossification in the condyle but, instead of joint cartilage, a periosteum-like membrane covered the pair of joint surfaces of the condyle and temporal bone. Therefore, TMJ and SIJ morphogenesis were quite different, except at an early stage when the cartilaginous condyle was attached to the bony temporal bone.
The humeroradial joint of two fetuses had joint cartilages facing other cartilages (Fig. 4A). The periosteum ended near the proximal and distal margins of the joint capsule, with no continuation to the joint cavity (Fig. 4B, C). There was endochondral ossification at the distal end of the periosteum in the humerus (Fig. 4B), but it was covered by the periosteum in the radius (Fig. 4C). In contrast to the TMJ, a periosteum-like membrane did not cover the joint surface; instead, a cartilage tissue was exposed to the joint cavity and it faced a thick synovial fold (Fig. 4D).
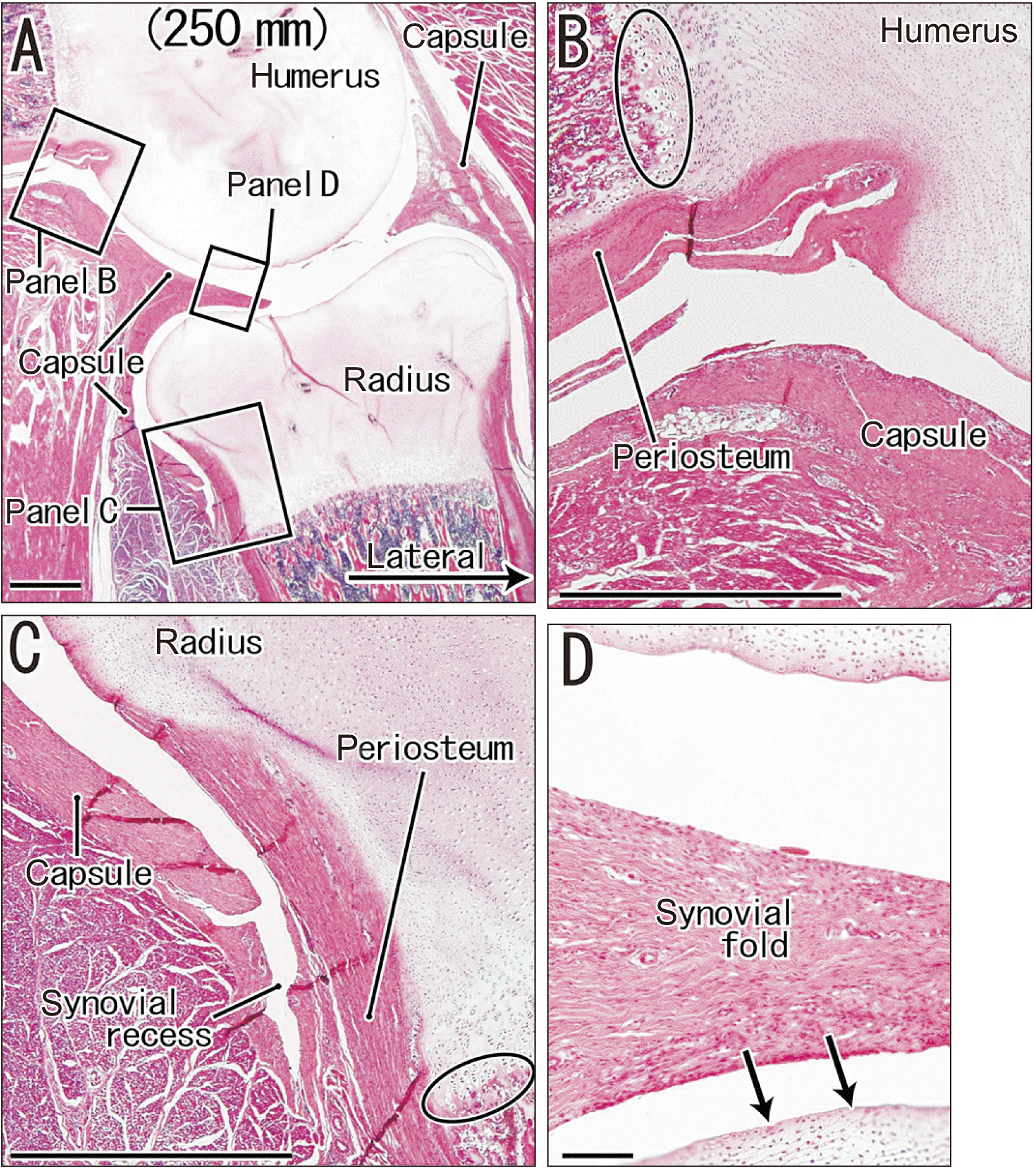 | Fig. 4Sagittal sections of the humeroradial joint in a fetus (crown-rump length: 250 mm) that were stained with H&E. (A) shows the topographical anatomy around the humeroradial joint. (B–D) are higher magnification views of three rectangles in (A). (B, C) show the periosteum and joint capsule at recesses of the joint cavity. (D) shows a thick synovial fold and its adjacent joint cartilage, and that the periosteum does not continue to the joint surface in the humeroradial joint (arrows). The oval in (B) shows the endochondral ossification is at the distal end of the periosteum in the humerus, and the oval in (C) shows it is covered by the periosteum in the radius. (A) Scale bar=1 mm; (B–D) Scale bars=0.1 mm.
|
Go to :
Discussion
Many studies have examined the development and growth of the SIJ (reviewed by Vleeming et al. [2]). However, no previous studies have examined why the joint cartilage of the ilium is always distant from and anterior to the hypertrophied cartilage tissues at the ossification center of the ilium. The ileal joint cartilage is not continuous with the cartilaginous ilium, and it develops as a mass at the “bony” ilium. The posterior extension of the joint cartilage occurs along the medial aspect of the ilium after completion of endochondral ossification. The sequential development and growth pattern of “ossification, followed by joint cartilage formation, and then cavitation” in the SIJ was quite different from that of in TMJ. After muscle contraction starts, it is likely that mechanical stress from the bony ilium induces joint cartilage development. Indeed, a joint between the cartilage and bone is common in the early SIJ and early TMJ, as previously hypothesized (Vleeming et al. [2], see Introduction), but the thick disk in the TMJ seemed to interfere with development of the joint cartilage along the aspect of the temporal bone.
The sacral joint cartilage is normally white and smooth, and resembles typical hyaline cartilage; the iliac cartilage is duller in appearance and consists of a dense bundle of fibrocartilage [8, 11]. Our observations suggest the sacral joint cartilage might differentiate after birth, in contrast to the unique posterior advancement of the iliac cartilage. Notably, we demonstrated that a pair of articular surfaces of the TMJ were not covered by cartilage, but were instead covered by a periosteum-like membrane, even at near-term. This periosteum-like covering was not on the humeroradial joint cartilage, although it did have a thick and tight synovial fold. Like the SIJ, the TMJ also seemed to develop a hyaline cartilage after birth, although descriptions of this are limited [20, 21]. Therefore, at least in fetuses, the TMJ and SIJ were quite different from the typical cartilage-to-cartilage joint (e.g., the humeroradial joint).
Cavitation of the SIJ occurs much later than in other joints. For instance, cavitation in the hip and shoulder joints is complete at 8 to 9 weeks GA [7, 22]. We found no evidence of macrophages at the future SIJ, so mechanical forces may be responsible for the cavitation, rather than programmed cell death in the interzone tissue. This delayed cavitation seemed to result from a specific posterior extension of the iliac joint cartilage. However, the delayed ossification of the sacrum might not be responsible for the unique morphology of the fetal SIJ, because ossification commonly occurs later than joint cavitation in the hip [7], knee [6], and shoulder [22]. Development and growth of the ilium from the anterior part to the posterior part seemed to connect to the anatomy of the iliacus muscle in fetuses: the latter muscle became thicker at the more anterior origin possibly because the insertion (i.e., femur trochanter) was situated far anterior to the SIJ.
The interarticular septa or disk divides the SIJ into cranial and caudal parts, and it usually disappears during the first postnatal year [2]. It is likely that we did not observe this structure because we only examined horizontal sections. Nevertheless, the septum (if present) seemed to have no effect on the sequential pattern of development and growth of the SIJ — ossification of the ilium, followed by joint cartilage formation, and then cavitation.
Go to :
 XML Download
XML Download