

 PDF
PDF Citation
Citation Print
Print



Introduction
Male infertility incidence is rising steadily worldwide and accounts for more than 50% of infertility cases [1, 2]. Equally, there is a worldwide increase in human immunodeficiency virus (HIV) infection [3] and alcohol abuse [4]. For instance, about 33% of South African (SA) population consumes alcohol regularly and the SA’s average consumed alcohol per capita (APC) almost doubles the global average APC [5, 6]. Equally, the country is burdened with high HIV prevalence and has the biggest combination antiretroviral therapy (cART) program globally [7, 8]. Currently, cART prophylaxis is encouraged among healthcare workers following accidental needle prick, and amongst prostitutes and individuals after unsafe and casual sex with unknown persons or HIV infected individuals [3, 9]. While the influence of alcohol on risky sexual activity is associated with an increase in the spread of HIV infection [10-12] and the consequent use of cART, many HIV patients on cART therapy continue to consume alcohol regularly for euphoric purposes [13, 14]. Furthermore, alcohol has been associated with reduced viral suppression dose-dependently [15, 16]; aggravated HIV progression, and reduced cART efficacy [14].
Alcohol and cART have each been shown to have negative long-term consequences on both male and female reproductive organs and other body systems [17-20]. Excessive alcohol consumption is reckoned as a leading cause of male impotence and infertility [21]. Several testicular and sexual dysfunctions have been reported in habitual/chronic alcohol consumers [22-24]. Also, cART has been shown to alter reproductive hormones and sperm parameters [25, 26]. A reduction in testosterone levels, sperm concentration, and DNA integrity have been implicated in long-term use of cART [27, 28]. Only two reports in literature have shown adverse effects of alcohol and cART interaction on testicular histology [29, 30]. The present study evaluated specifically the impact of this interaction on Sertoli and Leydig cells and immunohistochemical localization of the oxidative stress, inflammatory, and apoptotic markers to further delineate possible physio-biochemical impact of alcohol plus cART treatment on the testis. Currently, increasing HIV infection and male infertility are reported among reproductive age individuals, with high desire for procreation, and might also be regular alcohol consumers [30]. This implies increased alcohol and cART interaction amongst the age group, making their reproductive health a huge health concern. Against this backdrop, this study evaluated testicular toxicity associated with concurrence of alcohol and cART in the body using HIV-naïve male rat model to mimic conditions of cART regimen.
Materials and Methods
Chemical and reagents
These were procured from different reliable sources. A combination antiretroviral drug (Atripla) consisting of 600 mg of Efavirenz, 300 mg Tenofovir disproxil Fumurate, and 245 mg Emtricitabine was procured from Bristol-Myers Squibb and Gilead Sciences. Rabbit polyclonal for primary antibodies interleukin-1beta (IL-1β) (ab2105), tumor necrosis factor-alpha (TNF-α) (ab6671), and caspase 3 (ab4051), rabbit monoclonal for primary antibody inducible nitric oxide synthase (iNOS) (ab115819), and mouse monoclonal for primary antibodies interleukin-6 (IL-6) (ab9324), malondialdehyde (MDA) (ab243066), and 8-hydroxydeoxyguanosine (8-OHDG) (ab62623) were procured from Abcam . Whilst biotinylated goat anti-rabbit (BA-1000) and goat anti-mouse (BA-9200) secondary antibodies, and Avidin-Biotin Complex kit (ABC) (PK-6100) were procured from Vector Laboratories.
Ethical clearance
This study was approved by the Animal Research Ethics committee (AREC) (approval number: 2018/011/58/C) of University of the Witwatersrand (Wits) and all experiments were conducted at the Wits Research Animal Facility in compliance with the guidelines set forth by AREC.
Animals
Twenty-four adult male Sprague Dawley rats (10 weeks old), weighing between 330–370 g were used in the study. Each rat was separately housed under sterile conditions in a plastic cage. Animals were fed on normal rat chow and water ad libitum and were maintained at 21°C–23°C room temperature, with lighting conditions of 12/12-hour light/dark cycle.
Experimental design
Animals were divided into four groups of six rats each as follows; control group, received no treatment; alcohol group (A), which received 10% v/v alcohol in drinking water daily [31]; the cART group, received extrapolated human equivalent dose of 23 mg/kg of cART (consisting of efavirenz 600 mg+emtricitabine 200 mg+tenofovir 245 mg) in gelatine cubes daily [32]; and alcohol plus cART group (A+cART), received both alcohol and cART daily at same doses indicated above. After 90 days of treatment, the animals were weighed, anesthetized with 240 mg/ml pentobarbitone dose, and sacrificed. The animals were perfused with 2 ml/min of 0.1 M phosphate buffer (PB) in 0.9% saline and the testes were harvested, weighed, and the wet weight recorded and thereafter fixed in 10% neutral buffered formalin for histology and further analysis.
Testicular morphometry
Testis weight and volume
The weight of the testes was recorded immediately after their extraction using an electronic scale (Kern PCB 200-2 weighing scale). Testis volume was determined using the Archimedes principle of water displacement in a measuring cylinder [33]. The initial volume (Vi) of distilled water in a graduated cylinder was recorded, and then the final volume (Vf) was determined after adding the testis to the graduated cylinder containing the known amount of distilled water. The volume of the testis (Vt) was calculated by subtracting the initial volume from the final volume as shown below.
Histology and histomorphometry (H&E)
The fixed testes were dehydrated sequentially in 70%–100% alcohol grades and embedded in molten paraffin wax and serially sectioned at 5 μm thickness. The sections were stained with H&E and examined using the Axioskop 2 plus light microscope (Nikon Eclipse Ci, 104C type). Photomicrographs of testicular tissue sections were obtained using a linked computerized Zeiss digital image system, the AxioCam 208 color (Zeiss group) for morphometric assessment. Six camera fields from 4 sections per animal, from six animals per group (i.e., 144 images per group) were captured and used for morphometry and analysis.
Testicular component area fractions
Testicular component (epithelial, luminal, connective tissue, and interstitial space) area fractions were determined using Fiji 84-intersection grid. The grid was superimposed on H&E-stained images captured at ×100 and the intersecting points for each component were counted. The average intersecting points for each testicular component were calculated in 144 microscopic field images per group and each testicular component area fraction was determined using the formula below [35].
Seminiferous tubule morphometry
Seminiferous tubule area (TA), tubule diameter (TD), epithelium height (EH), and luminal diameter (LD) were measured in 50 round or nearly round seminiferous tubules for each animal (i.e., 300 tubules per group) using Fiji software. The seminiferous TA was determined by tracing around the seminiferous tubule using the Fiji freehand selection tool. The average TD and epithelial height were calculated using the minor and major axes measurements [36]. To exclude longitudinal tubules, an average TD was considered only if D1/D2 ≥0.85, a value of D1/D2=1.0 representing a perfectly rounded circle [37].
Germinal epithelium cell quantification
The number of the different germinal epithelium cells (Sertoli, spermatogonia, spermatocytes, round, and elongated spermatids) were counted in H&E-stained sections at ×400. Ten rounded seminiferous tubules (stage VII) [38] for each animal were considered (i.e., 60 tubules per group).
Leydig cell morphometry
Using Fiji software, diameters (D) of fifty Leydig nuclei with distinct nucleoli were measured for each animal on H&E-stained images captured at ×400. The nuclear volume (an important parameter in a variety of cell functions) was determined in accordance with previously reported methods [39], with the formula below.
Histopathological evaluation
Johnsen’s testicular score
Spermatogenesis was classified in 50 stage II-VII seminiferous tubules per animal (i.e., 300 tubules per group) using Johnsen’s testicular score. Each tubule was assigned a score from 10 to 1, accordingly, thereafter the number of tubules for an individual score was multiplied by the score, and then the total for all the scores was divided by 50 (the total number of tubules scored) to obtain the mean Johnsen’s score per group for further analysis [40].
The modified Johnsen scoring criteria used is shown in Table 1.
Histological changes
Testicular H&E-stained sections were examined for the presence of histological changes. Observed changes were graded in 24 microscopic fields (six fields from each of the four sections) for each animal (i.e., 144 fields per group). The changes in each field were semi-quantitatively categorized into five grades as follows; grade 4=very severe (change seen in >75% of the field), grade 3=severe (change seen in >50% <75% of the field), grade 2=moderate (change seen in >25% <50% of the field), grade 1=mild (change seen in <25% of the field), and grade 0=none (no change in the field) as previously reported by Erpek et al. [41].
Immunohistochemistry for oxidative stress, inflammation, and apoptosis
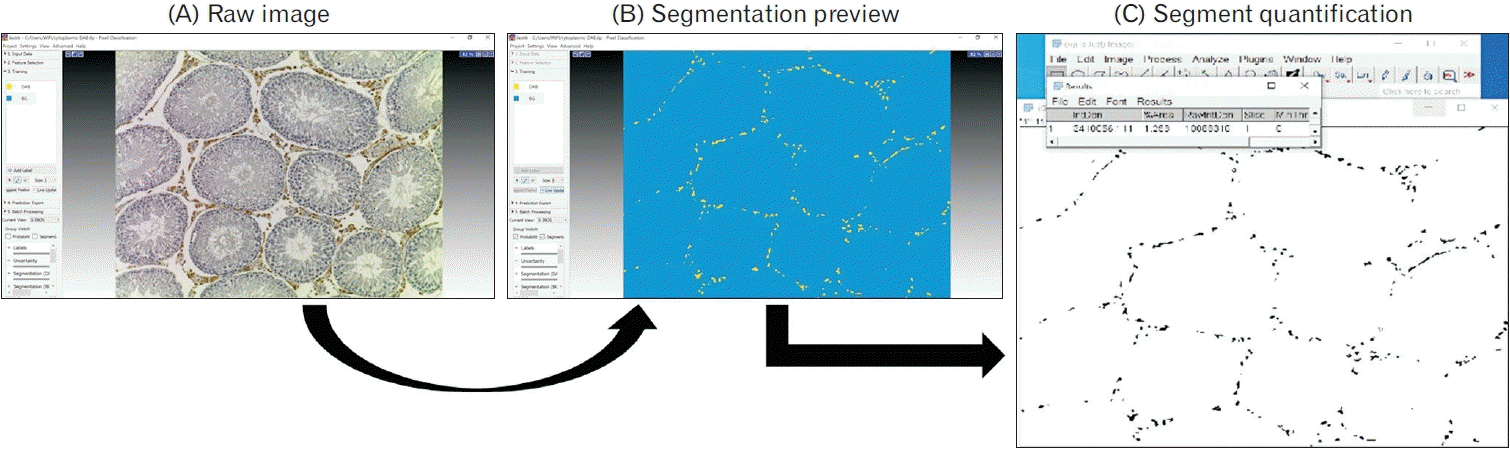
Testicular tissue sections were mounted on silane-coated slides for antibody immunolabeling. Assessment of oxidative stress using iNOS, MDA, and 8-OHDG, inflammation (IL-1β), TNF-α, and IL-6, and caspase 3 for apoptosis were carried out. The sections were incubated in citrate buffer overnight in a water bath at 60°C for antigen retrieval. Then, the endogenous peroxidase was inhibited using 1% hydrogen peroxide in methanol for 20 minutes. After rinsing in phosphate buffered saline (PBS) for three changes of five minutes each, 5% normal goat serum was added to the sections to block nonspecific antibody binding. This was tapped off after 30 minutes, and the primary antibody was added subsequently (1:100 for anti-TNF-α and anti-iNOS, 1:200 for anti-IL-1β, anti-IL-6, anti-MDA, and anti-caspase 3, and 1:1,000 for anti-8-OHDG) and left overnight (approximately 16 hours) at 4°C. Thereafter, the sections were rinsed in PBS and incubated with the secondary antibody (1:1,000 biotinylated goat anti-rabbit for the IL-1β, TNF-α, iNOS, and caspase 3 antibodies and 1:1,000 biotinylated goat anti-mouse for IL-6, MDA, and 8-OHDG antibodies) for 30 minutes. Then, after rinsing in PBS, ABC reagent was added for 30 minutes. Afterward, the sections were rinsed in PBS and incubated with 3, 3’-diaminobenzidine tetrachloride (DAB) for five minutes. DAB was then washed off under running tap water for five minutes and the slides were dipped in hematoxylin for one minute to counter stain. Followed by rinsing in running tap water to remove excess stain, dehydration of slides in alcohol series, and coverslipped with Dibutylphthalate Polystyrene Xylene. The images of antibodies with immunoreactivity localized to both cell nucleus and cytoplasm (IL-1β, TNF-α, iNOS, and MDA) were captured in 24 microscopic fields at ×100 for each animal (i.e., 144 sections per group) and were segmented using the ilastik pixel classification workflow. Fiji was used for quantification of the ilastik exported image segments to extract numerical data for statistical analysis. Details of image segmentation and quantification are shown in Appendix 1–2. The percentage area of expression for antibodies was calculated using the formula below.
For the antibodies with immunoreactivity localized to cell nuclei (IL-6, 8-OHDG, and caspase 3), the total number of cell nuclei expressing immunoreactivity were counted in 20 rounded stage VII seminiferous tubules for each animal (i.e., 120 tubules for each group).
Data analysis
GraphPad Prism 6 windows software was used for data analysis. Data were expressed as Mean±SEM. Group means of parametric data were compared using a one-way ANOVA, followed by Bonferroni’s post hoc test. Non-parametric data (Johnsen’s and histological change scores) were compared using Kruskall Wallis, followed by Dunn’s post hoc test. Deeming a P-value of <0.05 statistically significant.
Results
Testicular morphometry
All treated groups (A, cART, and A+cART) showed a non-significant decrease in testis weights, testis volume, and testis size in comparison with the control (Table 2).
Testicular area fractions
The epithelial area fraction (EAF) of treated groups significantly reduced compared to the control, (A, P=0.0012; cART, P=0.0078; and A+cART, P<0.0001) (Table 2). The luminal area fraction (LAF) of cART group decreased significantly compared to control (P=0.0123), A (P=0.0059) and A+cART (P<0.0001), but was significantly increased in A+cART compared to control (P=0.0141). The connective tissue and interstitial space area fractions (CTAF and ISAF respectively) of all the treated groups (A, cART, and A+cART) were increased compared to the control. The increase was significant in CTAF of all treated groups (A: P=0.0017, cART: P<0.0001, and A+cART: P<0.0001) and in ISAF of A (P=0.0008) and cART (P=0.0003) groups. Furthermore, CTAF of cART and A+cART treated groups was significantly higher when compared with group A (P=0.0110 and P=0.0038 respectively), while ISAF of A+cART group was significantly reduced compared to A (P=0.0025) and cART (P=0.0010) groups (Table 2).
Seminiferous tubule morphometry
All seminiferous tubule measured parameters (area, diameter, epithelium, and lumen) were decreased in all treated groups compared to control, except for lumen of A+cART treated that increased non-significantly (P>0.05) (Table 3). TA and diameter significantly decreased (P<0.0001) in A, cART, and A+cART treated groups relative to control. The TA of cART treated group significantly decreased compared to A treated group (P=0.0413). In addition, TA and diameter were significantly higher in A+cART than in A (P=0.0155 and 0.0089 respectively) and cART (P<0.0001) groups. In comparison with the control group, the epithelial height of groups A, cART, and A+cART decreased significantly (P=0.0002, 0.0001, and 0.0001 respectively) while the LD of A+cART group increased significantly (P=0.0013). However, the LD of cART group was significantly decreased compared to control (P=0.0340), A (P=0.0004), and A+cART (P<0.0001) treated groups.
Germinal epithelium cell count
The results are as shown in Table 3. The mean number of spermatogonia increased significantly in A and cART-treated groups (P<0.0001and P<0.0002 respectively) relative to control but was significantly decreased in A+cART (P=0.0032) relative to A group. However, spermatocytes were significantly reduced in cART and A+cART groups compared to control, (P=0.0043 and P=0.0007 respectively). Both round and elongated spermatids decreased significantly (P<0.0001) in all treated groups compared to the control. Furthermore, a non-significant decrease (P>0.05) in the number of Sertoli cells was found in all the treated groups (A, cART, and A+cART) compared to control.
Leydig cell
Leydig cell nuclear diameter and volume decreased significantly (P<0.0001) in all the treated groups (A, cART, and A+cART) compared to control (Table 3).
Histopathological evaluation
Johnsen’s testicular score
Johnsen’s score was used to profile the progress/status of spermatogenesis in the seminiferous tubules. Compared to the control, a significant reduction in Johnsen’s score was recorded in A (P=0.0115) and A+cART groups (P=0.0256) (Table 4).
Histological changes
The most common histological changes observed were lifting of germinal epithelium, widened intercellular space, and karyolysis (Fig. 1) and were different across the groups (Table 4). However, the difference in epithelial lifting was not significant across the groups but widened intercellular space was significantly higher in A+cART (P=0.0029) compared to control group. Additionally, karyolysis in cART and A+cART groups was significantly high when compared to the control group (P=0.0081 and 0.0346 respectively). Furthermore, seminiferous tubule shrinkage was observed in all treated groups, with sloughing of germinal epithelium into the lumen recorded in A+cART treated animals.
Immunohistochemistry: oxidative stress, inflammation, and apoptosis
Oxidative stress markers
Oxidative stress markers immunoreactivity (Fig. 2) was localized in the cytoplasm and nucleus of macrophages and Leydig cells for both iNOS and MDA. While 8-OHDG immunoreactivity was localized in the spermatogenic cell nuclei and residual bodies. All treated groups (A, cART, and A+cART) demonstrated a significant increase (P<0.0001) in expression of iNOS and 8-OHDG relative to the control group (Fig. 2). A significant increase of MDA expression was detected in cART and A+cART groups relative to the control group (P<0.0001) and in cART and A+cART groups relative to group A (P<0.0001 and P=0.0302 respectively) (Fig. 2).
Inflammatory and apoptosis markers
Expressions of IL-1β, TNF-α, IL-6, and caspase 3 were as presented in Fig. 3. IL-1β expression was significantly elevated (P<0.0001) in A and cART treated groups compared to the control. But the expression of IL-1β in cART and A+cART treated groups was significantly decreased relative to A treated group (P=0.0006 and P<0.0001 respectively) and significantly decreased (P<0.0001) in A+cART compared to cART. Although, the expression of TNF-α was increased in all the treated groups against the control, the increase was only significant in A treated animals (P=0.0001). Further, the expression TNF-α in A group increased significantly (P=0.0011) compared to A+cART treated group. IL-6 was significantly upregulated (P<0.0001) in all treated groups (A, cART, and A+cART) compared to the control. Whilst caspase 3 expression significantly increased in A (P=0.0023), cART (P<0.0001), and A+cART (P<0.0001) groups relative to control. Further, caspase 3 expression in A+cART group was significantly higher in comparison to A (P<0.0001) and cART (P=0.0013) treated groups (Fig. 3).
Discussion
Infertility is a worrisome reproductive health problem in families globally [2] with a soaring prevalence of male infertility [1]. The chronic use of combined antiretroviral (cART) blended with alcohol abuse has introduced new variables in male reproductive dysfunctions. Moreover, a high incidence of alcohol use among people living with HIV/AIDS (PLWHA) and on cART has also been reported [13, 14]. Markedly, cART and alcohol have individually been reported to negatively impact the male reproductive system [17, 19] and thus their concomitant use may pose major reproductive dysfunctional challenges, particularly amongst PLWHA that desire to have biological children [26], which is a critical societal necessity.
The result from this study showed non-significant decrease in testicular weight, volume, and size across the treatment groups compared to the control group which is consistent with previous reports [29, 37, 39, 42] and may suggest minimal perturbations of testicular architecture. However, the significantly decreased EAF, and the significantly increased LAF in A+cART treated group point to perturbations of testicular structure and cellular coherency. Furthermore, the significant increases in ISAF in A and cART treated groups and non-significant in the A+cART treated group may suggest specific treatment effects and the impact of the A+cART interaction on this parameter. Overall, these changes in the histological component of the testis reflect an alteration of the integrity of testicular morphology, pointing to disruption of the normal testicular paracrine/autocrine factors (such as cytokines) involved in regulation of spermatogenesis and consequently will lead to a decline in spermatogenesis [27, 35].
Furthermore, the increased number of spermatogonia (proliferation of spermatogenesis) observed in testis of the alcohol and cART treated groups, could be an attempt to increase the germ cell pool so as to counteract the diminishing spermatogenesis processes which is in line with a previous clinical study that reported a massive increase in the number of normally differentiated spermatogonia and elevated spermatogonia proliferative activity in a biopsy of patient with non-obstructive azoospermia [43]. Additionally, the (LAF and diameter (LD) were both slightly increased in the A group but significantly increased in the A+cART group, suggesting varying degrees of germ cell layer reduction due to different treatments as well as seminiferous tubule shrinkage.
Notably, tubular shrinkage was depicted by the significant reduction in TA and TD in testis of treated groups (A, cART, and A+cART), and was consistent with previous findings [27, 29, 44]. However, LAF and LD were also significantly decreased in the treated groups except in cART group regardless of the diminished EAF, EH, and germinal epithelium cell layers. This could have been due to the prominent seminiferous tubule shrinkage observed in cART treated group which also recorded the greatest reductions in TA and TD. Further, shrunken seminiferous tubules are suggestive of tubule degenerative changes induced by the treatments and subsequently result in germ cell death [45]. Accordingly, shrinkage of the seminiferous tubules resulted in an increase in ISAF in the treated groups compared to the control animals.
Equally, a significant increment in CTAF was recorded in the testis of treated animals, suggesting formation of fibrosis in the testis interstitium. Our findings go in line with that of a previous study that reported fibrosis led to increase in thickness of the testicular interstitium in the infertile males [46]. Testicular fibrosis usually occurs following testis trauma and inflammation amongst others [47]. Fibrosis impairs the integrity of the testicular interstitium and alters the structure and function of the interstitial cells (Leydig, macrophages, and myoid cells), and consequently adversely affects testicular structure and function [47, 48].
Furthermore, the number of Sertoli cells was not significantly reduced in the treated animals compared to control animals, suggesting that Sertoli cells were less adversely affected. Previous report indicated that Sertoli cells are resistant to apoptosis compared to germ cells [49], because they contain high levels of anti-apoptotic markers, Bcl-w and Bcl-xL [50]. It has been reported that while the number of Sertoli cells is not usually affected [50, 51], disruptions in its structure are relatively common and can alter the cell’s functions, the integrity of the blood-testis barrier [52] and consequently lead to dysregulation of spermatogenesis. Though, the present study did not include effects on the Sertoli cell cytoskeleton, previous studies show that its alteration and/or damage leads to sloughing of germ cells into the lumen and separation of germinal epithelium from the basement membrane [53-55], which were both observed in treated animals in the current study.
However, Leydig cell nuclear diameter and volume in testis of treated animals were significantly reduced relative to the control animals and may imply disturbances of Leydig cell function [37, 52, 56]. The nucleus is the lifeline of the cell, and a previous study showed that quantitative measurements of the nucleus is important in understanding the cell’s response to injury and further reported that nuclear volume correlates strongly with cell volume and function [57]. Leydig cells are present in the testis interstitium, and synthesize androgens via steroidogenesis [58]. Therefore, shrinkage of the nucleus will ultimately affect the Leydig cell’s steroidogenic capacity and subsequently lead to disruption of spermatogenesis [58-60].
Additionally, in parallel with previous findings [61, 62], Johnsen’s score of the treated groups was reduced compared with the control animal group. Reduction in Johnsen’s score is linked to interruption of normal germ cell maturation, germ cell layer loss, and tubule lesions [63, 64]. Also, seminiferous tubule germinal epithelium lesions such as lifting of epithelium, widened interstitial space, karyolysis, and sloughed germ cells were more evident in the treated groups compared to control animal group, which supports the reduction in Johnson’s score recorded in treated groups. This result confirm testicular toxicity of the treatments (alcohol or/and cART) [20, 53].
Oxidative stress is a common feature in most male reproductive system dysfunctions [65-67], occurring as a result of an imbalance between rate of production and clearance of reactive oxygen species (ROS) [68, 69]. High ROS levels have been reported in 30%–80% of infertile males [65, 69]. Our findings showed upregulation of oxidative stress markers i.e., iNOS, MDA, and 8-OHDG in the testis of treated animals. This indicates that alcohol, cART, and their combination induced oxidative stress in the testis. Reports show that iNOS upregulation leads to increased ROS formation [70], ROS have been shown to induce lipid peroxidation and DNA fragmentation in the testis [71-73]. These are the most common consequences of testicular oxidative stress resulting into increased MDA and 8-OHDG formation respectively [66, 74, 75]. Previous studies have reported that ROS-induced lipid peroxidation is implicated in male reproductive dysfunction [65-67], due to substantial amounts of unsaturated fatty acids in the testis tissue [71, 76].
Reportedly, elevation of ROS stimulates release of several inflammatory cytokines such as IL-1, TNF-α, and IL-6 mostly via the nuclear factor-kappa beta pathway [74, 75, 77]. In the testis, cytokines are majorly derived from interstitial macrophages, but Leydig and Sertoli cells also secrete cytokines [78-80]. Studies show that during testicular injury, activated macrophages, Leydig, and Sertoli cells produce a high concentration of cytokines and iNOS [75, 78, 79]. In consonance with the above, the elevated levels of pro-inflammatory cytokines observed in the present study may adversely affect the process of spermatogenesis, as reported by previous studies [80, 81]. Somade et al. [82] found that elevated cytokine levels in testis and kidney of Wistar albino rats, was associated with testiculo-renal toxicosis following acute administration of edible camphor. Additionally, testicular cytokines have been shown to affect germ cell proliferation, as well as Leydig and Sertoli cells functions and secretions [80]. This study findings show that oxidative stress and inflammation are interlinked mechanisms associated with germ cell apoptosis, which might underlie the multidimensional changes observed in testis of animals treated with alcohol or/and cART.
Overall, the study has revealed some important findings in testicular toxicity of the treatments relative to the parameters studied. Interestingly, though the levels of IL-1β and TNF-α, and MDA were elevated in A+cART treated group compared to control animals but comparatively were not as elevated as that of IL-6, iNOS, 8-OHDG, and caspase 3 expression in A+cART treated group. These results suggest possible effect of cytochrome P450 enzyme pathway involved in the metabolism of alcohol and cART [14, 30, 83], resulting in reduction of the level of these parameters (IL-1β, TNF-α, and MDA) in A+cART treated group. Additionally, it may suggest a variation in the effects of different treatment on the studied parameters.
In conclusion, our results show that consumption of alcohol or cART can cause perturbations in the testis morphometric parameters and seminiferous tubule lesions via upregulation of oxidative stress and pro-inflammatory cytokines. In addition, consumption of alcohol and cART concurrently can exacerbate testicular damage. The findings from this study are invaluable for the clinical management of male fertility challenges that could emerge in PLWHA on cART therapy who consume alcohol regularly.



 XML Download
XML Download