

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
A subtype of colorectal cancer (CRC) called rectal cancer (RC) is the 4th most common cause of cancer-related morbidity and mortality worldwide [1]. According to statistics, there are about 39,000 new cases of RC annually in the United States [2]. Due to early symptoms of RC are not obvious, many RC patients are diagnosed at advanced stages [3,4]. Current treatment strategies for RC mainly include surgical operation, innovative chemotherapy, radiotherapy and immunotherapy [5-7]. Although much advance has been achieved in therapeutic options, prognosis is still not optimistic, especially for RC patients with tumor metastasis or at advanced stages [8-10]. Notably, lung metastasis is the most common in patients with advanced RC, affecting the survival rate of RC patients [11,12]. Therefore, it is important to explore the mechanisms of lung metastasis and to seek new treatment approaches for RC.
Exosomes refer to micro vesicles from 30 to 100 nm in diameter [13]. They develop as poly vesicles, and they are fused with cytomembrane to be released into the extracellular medium [14]. It is reported that oncogenic proteins in tumor exosomes can spread to non-tumor cells in the tumor microenvironment and accelerate tumor progression [15,16]. Among enriched proteins of exosomes, integrin beta-1 (ITGB1) has caught our attention [17]. An integrin known as ITGB1 is mainly expressed in tumor-related cells, affecting the growth and spread of tumors [18,19]. Many reports have demonstrated the important role of ITGB1 in CRC [20,21]. For instance, Ropivacaine represses proliferation, migration and invasion of CRC cells via reducing ITGB1 expression [20]. ITGB1 overexpression attenuates the anti-tumor impact of cetuximab on CRC cells [21]. Nevertheless, there is lack of exploration in the influence of exosome-derived ITGB1 on lung fibroblasts in RC.
Carcinoma-associated fibroblasts (CAFs) are activated population of fibroblasts that exhibit increased production of pro-inflammatory cytokines, which apparently promote the development and metastasis of tumors [22,23]. Many tumors have persistent activation of the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB) pathway in CAFs, which promotes tumor growth by producing pro-inflammatory cytokines such as interleukin-8 (IL-8) and interleukin-6 (IL-6) [24,25]. For example, Su et al. [24] have shown that CAFs can facilitate expression levels of interleukins through activating NF-κB pathway in lung cancer. Goulet et al. [25] have demonstrated that CAFs-secreted IL-6 helps bladder cancer cells go through tumor epithelial-mesenchymal transition. However, underlying activation mechanisms of fibroblasts remain unclear in RC.
In the present study, the expression of ITGB1 in tumor-derived exosomes and the impact of exosomal ITGB1 on activation of lung fibroblasts were investigated. Besides, whether exosomal ITGB1 could activate lung fibroblasts through regulating the NF-κB pathway was explored.
Go to : 
METHODS
Cell culture
Human lung fibroblasts (MRC5), a RC cell line (SW837) and a human normal colorectal epithelial cell line (FHC) were purchased from the Chinese Academy of Sciences. All cells were cultured in Dulbecco's modified Eagle's medium (Invitrogen) containing 10% fetal bovine serum (Gibco). All cells were incubated at 37°C with 5% CO2. For investigating involvement of NF-κB, MRC5 cells were treated with 18 μM NF-κB inhibitor (SN50; MedChemexpress) for 24 h [26].
Exosome isolation and analysis
Exosomes from FHC and SW837 cells were extracted using an ExoQuick precipitation kit (System Biosciences). In brief, collected culture medium was centrifuged for 15 min. Then supernatant (250 μl) was mixed with ExoQuick precipitation (63 μl) to incubate for 30 min, followed by centrifugation at 4°C. Subsequently, cell suspension was removed and centrifuged for another 5 min. Finally, the exosome pellet was re-suspended in 200 μl of phosphate-buffered saline.
To identify exosomes, exosome suspension was dropped onto copper grids with a mesh size of 300 (Agar Scientific). These samples were then fixed with 1% osmium tetroxide for 1 h and 3% paraformaldehyde overnight. Samples were stained with uranyl acetate for 10 min and lead citrate for 5 min after being washed with sterlized water. The last step was to view exosomes under a 120 kV transmission electron microscope (JEOL). The amount of exosomes was examined via a bicinchoninic acid protein assay kit (Novagen). Additionally, Western blot analysis was used to determine expression levels of exosome marker proteins (CD63 and CD9).
Cell transfection
Small interfer RNA against ITGB1 (si-ITGB1) and si-negative control (NC) were bought from RiboBio. The sequences of si-ITGB1 and si-NC were shown as:
si-ITGB1-F: GCGAGUGUGAUAAUUUCAATT
si-ITGB1-R: UUGAAAUUAUCACACUCGCTT
si-NC-F: UUCUCCGAACGUGUCACGUTT
si-NC-R: ACGUGACACGUUCGGAGAATT
SW837 cells in six-well plates (5 × 105/well, 80% confluency) were transfected with si-ITGB1/si-NC plasmids using Lipofectamine 3000 (Invitrogen) for 48 h.
Quantitative real-time polymerase chain reaction (qRT-PCR)
Total RNAs were extracted from SW837 cells using TRIzol reagent (Invitrogen), after which these RNAs were reverse-transcribed to complementary DNAs (cDNAs) through a PrimeScript II 1st Strand cDNA Synthesis Kit (Applied Biosystems). Afterward, PCR was performed according to instructions of a SuperReal SYBR Green kit (Applied Biosystems). The reaction condition was listed as follows: 2 min at 94°C, 40 cycles of 20 sec at 94°C, 30 sec at 58°C, and 2 min at 72°C. The following primers (Sangon) were used:
ITGB1-F: AACCAGGCCCCTCCTTACTC
ITGB1-R: GATGTGTCCGAAGGCTAGGA
GAPDH-F: CCATCTTCCAGGAGCGAGAT
GAPDH-R: TGCTGATGATCTTGAGGCTG
Using the 2−ΔΔCt method [27], relative mRNA expression of ITGB1 was calculated and normalized against GAPDH.
Western blot analysis
RIPA lysis buffer was used for protein extraction (Beyotime). Electrophoresis on 10% sodium dodecyl sulphate polyacrylamide gel was then used to separate proteins. Proteins were blocked with 5% milk after they had been transferred to PVDF membranes (Millipore). The secondary antibody was then added to the membranes after the primary antibodies had been incubated with them at 4°C overnight. Protein blots were determined using the ECL chemiluminescent system, and their intensity was evaluated using Image J software from the National Institutes of Health in the United States. All antibodies were bought from Abcam, including CD63 (1:2,000, ab68418), CD9 (1:1,000, ab236630), ITGB1 (1:2,000, ab52971), p65 (1:5,000, ab16502), phosphorylated (p)-p65 (1:5,000, ab86299), β-actin (1:2,000, ab8227) and the secondary antibody (1:2,000, ab6721).
Enzyme-linked immunosorbent assay (ELISA)
The levels of interleukins in the culture medium of cell lines were measured via R&D Systems' commercial ELISA kits.
Statistical analysis
GraphPad Prism 7.0 (GraphPad Software Inc.) was utilized for performing statistical analysis. The mean and standard deviation for all three experiments' data were shown. The Student's t-test was used to compare two groups' differences. To examine differences between multiple groups, a one-way ANOVA was used, followed by a Tukey's test. The threshold for statistical significance was p < 0.05.
Go to :
RESULTS
ITGB1 expression is increased in exosomes from SW837cells
Firstly, exosomes isolated from FHC and SW837 cells were identified using a transmission electron microscope. As expected, similar morphological characteristics of FHC cells-derived exosomes and SW837 cells-derived exosomes were observed (Fig. 1A). Next, expression levels of exosome marker proteins (CD63 and CD9) and ITGB1 were examined using Western blot. We found that CD9 and CD63 were all highly expressed in exosomes isolated from FHC and SW837 cells without significant difference (Fig. 1B). In contrast, ITGB1 displayed higher expression in exosomes from SW837 cells than that in exosomes from FHC cells (Fig. 1B, p < 0.01).
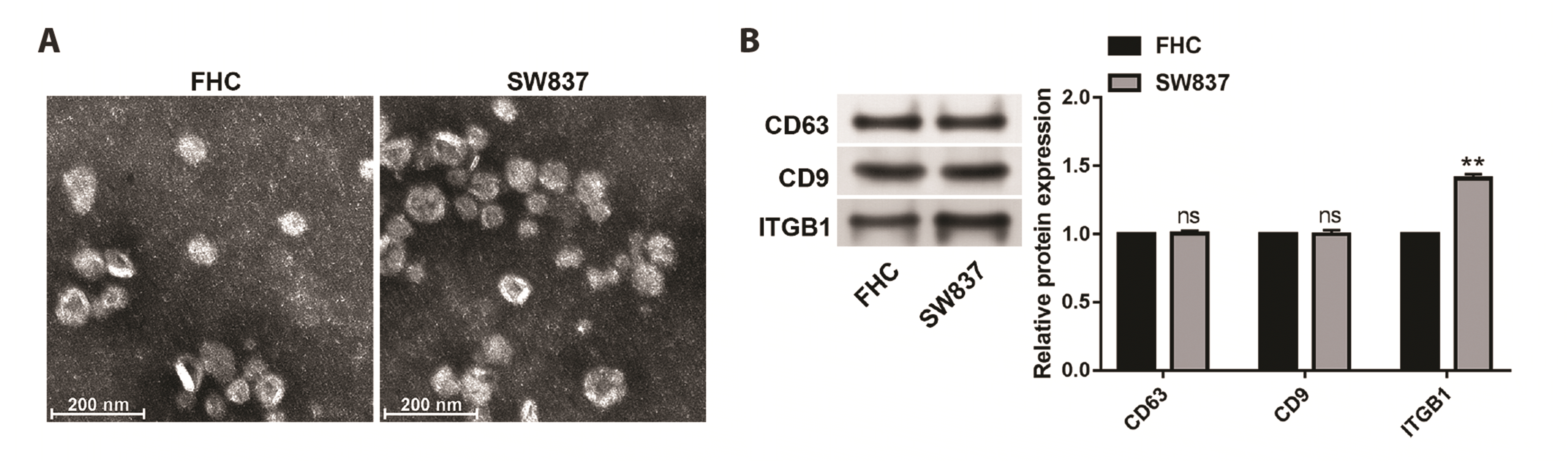 | Fig. 1ITGB1 expression is increased in exosomes from SW837 cells.(A) Transmission electron microscopy images of exosomes isolated from FHC and SW837 cells. (B) The protein expression of CD63, CD9 and ITGB1 was detected by Western blot. Values are presented as mean ± SD. ns, not significant. **p < 0.01.
|
Exosomes derived from SW837 cells activate the NF-κB pathway and lung fibroblasts
In order to explore whether tumor-derived exosomes made impact on human lung fibroblasts (MRC5), subsequent experiments were carried out. First of all, exosomes derived from FHC cells or SW837 cells were co-cultured with MRC5 cells, and protein expression of ITGB1 was determined. It turned out that ITGB1 expression was evidently increased in MRC5 cells co-cultured with exosomes from SW837 cells compared to MRC5 cells co-cultured with exosomes from FHC cells (Fig. 2A, p < 0.01). Next, whether the NF-κB pathway was activated by exosomes derived from SW837 cells was validated through Western blot. It was found that the ratio of p-p65/p65 was raised in MRC5 cells co-cultured with exosomes from SW837 cells compared to MRC5 cells co-cultured with exosomes from FHC cells (Fig. 2B, p < 0.01). More importantly, higher levels of pro-inflammatory cytokines (IL-8, IL-1β and IL-6) were observed in MRC5 cells co-cultured with exosomes from SW837 cells than MRC5 cells co-cultured with exosomes from FHC cells, which played important roles in regulation of inflammation microenvironment and facilitated carcinoma development (Fig. 2C, p < 0.01).
 | Fig. 2Exosomes derived from SW837 cells activate the NF-κB pathway and lung fibroblasts.(A) The protein expression of ITGB1 was detected by Western blot in the co-culture system of MRC5 cells and exosomes derived from FHC or SW837 cells. (B) The protein expression of p-p65 and p65 was detected by Western blot in the co-culture system of MRC5 cells and exosomes derived from FHC or SW837 cells. (C) Protein levels of IL-6, IL-8 and IL-1β were measured using ELISA in the co-culture system of MRC5 cells and exosomes derived from FHC or SW837 cells. Values are presented as mean ± SD. **p < 0.01.
|
ITGB1 down-regulation blocks activation of the NF-κB pathway and lung fibroblasts induced by RC-derived exosomes
After that, we explored whether ITGB1 affected activation of the NF-κB pathway and lung fibroblasts caused by RC-derived exosomes. Firstly, ITGB1 was silenced via transfection of si-ITGB1, and transfection efficiency was tested via qRT-PCR and Western blot. As manifested in Fig. 3A and 3B, both mRNA expression and protein expression of ITGB1 were reduced in response to transfection of si-ITGB1 in SW837 cells (p < 0.001). Then exosomes from si-NC or si-ITGB1 transfected SW837 cells were collected and identified. We found that protein levels of CD63 and CD9 did not show significant changes between exosomes from si-NC transfected SW837 cells (si-NC-Exo) and exosomes from si-ITGB1 transfected SW837 cells (si-ITGB1-Exo), while the protein level of ITGB1 was markedly reduced in si-ITGB1-Exo compared to si-NC-Exo (Fig. 3C, p < 0.001).
 | Fig. 3ITGB1 down-regulation blocks activation of the NF-κB pathway and lung fibroblasts induced by RC-derived exosomes.(A) The mRNA expression of ITGB1 was detected by qRT-PCR in SW837 cells. (B) The protein expression of ITGB1 was detected by Western blot in SW837 cells. (C) The protein expression of CD63, CD9 and ITGB1 was detected by Western blot in exosomes isolated from SW837 cells. (D) The protein expression of ITGB1 was detected by Western blot in MRC5 cells. (E) The protein expression of p-p65 and p65 was detected by Western blot in the co-culture system of exosomes isolated from SW837 cells and MRC5 cells. (F) Protein levels of IL-6, IL-8 and IL-1β were measured using ELISA in MRC5 cells. Values are presented as mean ± SD. ns, not significant. **p < 0.01, ***p < 0.001.
|
When MRC5 cells were co-cultured with si-NC-Exo or si-ITGB1-Exo, we found that ITGB1 expression was higher in MRC5 cells co-cultured with si-NC-Exo compared to the control group (MRC5 cells without any treatment) and lower in MRC5 cells co-cultured with si-ITGB1-Exo compared to MRC5 cells co-cultured with si-NC-Exo (Fig. 3D, p < 0.01). Also, the ratio of p-p65/p65 as well as protein levels of pro-inflammatory cytokines (IL-8, IL-1β and IL-6) was decreased after ITGB1 knockdown in the co-culture system of SW837 cells-derived exosomes and MRC5 cells (Fig. 3E, F, p < 0.01).
Inhibition of NF-κB pathway reverses the activation effect of exosome ITGB1 on lung fibroblasts
At last, the involvement of the NF-κB pathway in exosomal ITGB1-mediated activation of lung fibroblasts was verified. It was found that addition of NF-κB inhibitor (SN50) did not affect protein expression of ITGB1 in MRC5 cells, whereas addition of SW837 cells-derived exosomes up-regulated protein expression of ITGB1 in MRC5 cells (Fig. 4A, p < 0.01). Addition of SN50 resulted in the decreased p-p65/p65 ratio and decreased levels of pro-inflammatory cytokines (IL-8, IL-1β and IL-6) in MRC5 cells, and addition of SW837 cells-derived had opposite roles (Fig. 4B, C, p < 0.01). At the same time, the promoting impacts of SW837 cells-derived exosomes on p-p65/p65, IL-6, IL-8 and IL-1β were reversed by addition of SN50 in MRC5 cells (Fig. 4B, C, p < 0.01).
 | Fig. 4Inhibition of NF-κB pathway blocks the activation effect of exosome ITGB1 on lung fibroblasts.(A) The protein expression of ITGB1 was detected by Western blot. (B) The protein expression of p-p65 and p65 was detected by Western blot. (C) Protein levels of IL-6, IL-8 and IL-1β were measured using ELISA. Values are presented as mean ± SD. ns, not significant. **p < 0.01, ***p < 0.001.
|
Go to :
DISCUSSION
In recent years, with the improvement and changes of life standards, risk factors of RC also increased and the age of onset is getting younger [28]. Thence, developing new ways to prevent RC development is imperative. Existing evidence has indicated that exosomes derived from cancer cells can convert different cell types to CAF-like cells, thereby promoting tumor metastasis and development [29-31]. Of note, exosomes from the CRC cells are demonstrated to promote fibroblast-mediated invasion of cancer cells [32]. Here, our data indicated that exosomes from SW837 cells activated the NF-κB pathway and MRC5 cells, manifesting as increased ratio of p-p65/p65 as well as increased levels of IL-8, IL-1β and IL-6. These results suggested that exosomes from RC cells activated lung fibroblasts, and may promote lung metastasis of RC cells.
Furthermore, previous studies have shown that ITGB1 is aberrantly expressed and possesses oncogenicity in CRC [20,21]. Wang and Li [20] have uncovered that ITGB1 expression is up-regulated in CRC cells, and Ropivacaine suppresses metastasis of CRC cells via reducing ITGB1 expression. Yang et al. [21] have demonstrated that ITGB1 overexpression attenuates the anti-tumor impact of cetuximab on CRC cells. In this study, we also discovered that exosomes from SW837 cells had higher levels of ITGB1 expression than exosomes from FHC cells. More significantly, we found that ITGB1 knockdown prevents RC-derived exosome-induced activation of NF-κB pathway and lung fibroblasts. Taken together, above findings suggested that treatments targeting ITGB1 may be a meaningful option for preventing RC development.
In cancer-associated fibroblasts, the NF-κB pathway is reported to be constantly activated [33]. In this study, for verifying whether the NF-κB pathway was responsible for exosomal ITGB1-induced activation of lung fibroblasts, rescue assays were performed. It turned out that addition of SN50 resulted in significant reduction of the p-p65/p65 ratio as well as IL-8, IL-1β and IL-6 levels in MRC5 cells. At the same time, the promoting effects of SW837 cells-derived exosomes on p-p65/p65, IL-8, IL-1β and IL-6 were reversed by addition of SN50. Based on these results, we deduced that exosomal ITGB1 facilitated activation of lung fibroblasts through activating of the NF-κB pathway.
In summary, our findings indicated that ITGB1 expression is increased in exosomes from SW837 cells. Functionally, down-regulation of exosomal ITGB1 repressed activation of the NF-κB pathway and lung fibroblasts induced by RC-derived exosomes. Our study elucidated a new crosstalk between ITGB1 from tumor-associated exosomes and activation of lung fibroblasts, which contributed to development of RC treatment.
Go to :
 XML Download
XML Download