

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
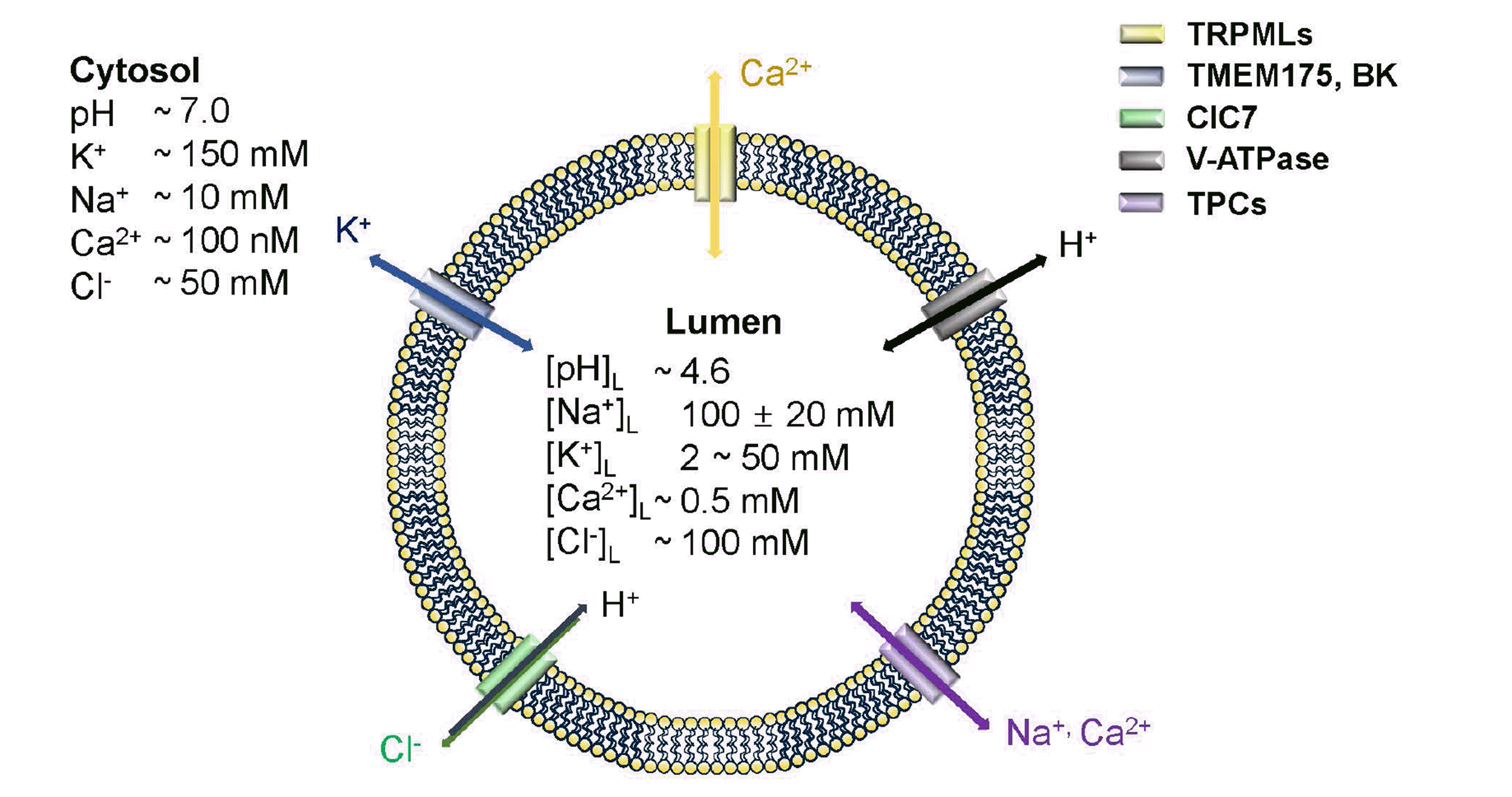
Lysosomes play vital roles in maintaining extracellular and intracellular macromolecules while regulating various degradation processes. They modulate ion-related functions within endocytic, exocytic, and other signaling pathways, thereby contributing to cellular signaling. Lysosomes are distributed throughout the body, and defects in lysosomal function are associated with lysosomal storage disorders (LSDs). To regulate lysosomal function, ion channels, and transporters are necessary to maintain ion concentration gradients of Na+, K+, Ca2+, H+, and Cl-. Each ion channel operates independently and in correlation with the others, responding to changes in individual ion concentrations and aiding in the establishment of the lysosomal membrane potential. Lysosomes maintain an acidic pH range (approximately 4.5–5.5) to support the enzymatic activity required for degradative functions. Ion channel activity influences acidic conditions and plays crucial roles in diverse cellular processes, including exocytosis, calcium release, wound repair, and recycling. Autophagy and cellular signaling can be severely affected by lysosome dysfunction. Dysfunctions in lysosomal ion channels contribute to the development of various diseases, including LSDs, neurodegenerative diseases, metabolic diseases, such as Mucolipidosis IV (MLIV), and autoimmune diseases. This highlights the importance of identifying novel therapeutic targets as increased lysosomal dysfunction may contribute to the prevalence of these diseases.
Numerous reviews have indicated the potential of lysosomal ion channels as therapeutic targets in these diseases [1-3]. In the context of the Coronavirus disease 2019 (COVID-19) era, the lysosomal two-pore channel (TPC) has emerged as a novel target for regulating virus-associated diseases such as severe acute respiratory syndrome (SARS), Ebola, and COVID-19.
HISTORY OF LYSOSOMES
The first descriptions of LSD phenotypes were made in the 19th century [4,5]. However, it wasn't until 1955–1956 that lysosomes were identified. de Duve [6] played a crucial role in this discovery by identifying lysosomes as intracellular catabolic stations. This has marked the beginning of extensive research on lysosomes. In one of his studies, he examined the localization of glucose-6 phosphatase in hepatic fractions, thereby providing mechanistic insights. This allowed him to isolate and identify non-specific acid phosphatases and glucose-6-phosphate. Christian discovered acid phosphatase-rich regions between microsomes and mitochondrial fractions, which he termed the "L-fraction" [7]. They named it the lysosome and hypothesized it functioned as a protein degradative organelle. Initially, lysosomes were considered garbage boxes that were primarily involved in recycling and digestion. However, recent studies have provided new insight into their roles in autophagy and mitochondrial function. Furthermore, there has been a renewed emphasis on understanding their function as nutrient sensors.
BIOLOGICAL FUNCTIONS OF LYSOSOMES
Numerous lysosomal-resident proteins play significant roles in cellular signaling pathways associated with organelle function. Approximately 50 proteins were categorized as acidic hydrolases [8-10]. Most of these proteins are localized within the lysosomal lumen and function as sulphatases or exoglycosidases. However, some proteins found in the lysosomal membrane contribute to various functions, including acidification, transport, facilitation of interactions between lysosomes and other organelles, and maintenance of membrane stability [11,12]. Notably, lysosomal proteins do not operate in isolation but rather interact with other proteins. These interactions are essential for preserving the integrity of lysosomes and ensuring the proper functioning of lysosomes, both internally and externally. Some of the proteins involved in these interactions include protein kinase B (AKT), mechanistic targets of rapamycin complex 1 and 2 (mTORC1 and mTORC2), transcription factor EB (TFEB), folliculin (FLCN), FLCN-interacting protein, signal transducer and activator of transcription-3, and energy-sensing complex AMP-activated kinase (AMPK) [13-29].
Lysosomes play a primary role in the degradation process by influencing various components such as nucleic acids, sphingolipids, proteins, glycosaminoglycans, and complex lipids. They break down the extracellular and intracellular materials that are old or no longer useful, resulting in the production of amino acids, fatty acids, and saccharides. These dissolved components and substrates are then directed to their designated destinations via diverse routes, such as endocytosis, exocytosis, phagocytosis, and autophagy [2,30-32].
Autophagy, also known as the autophagy-lysosomal pathway, plays a crucial role in regulating cellular processes, such as intracellular clearance, recycling of molecules and cellular components, and preservation of intracellular energy metabolism. It involves the formation of phagophores and the recruitment of cargo, eventually leading to the fusion of lysosomes to form autolysosomes [9,16]. Lysosomal degradation is essential for autophagy and the elimination of organelles from cells. Defective lysosomes can induce cytotoxicity and trigger unexpected inflammatory responses. Over 60 known nucleases, phosphatases, lipases, and proteases degrade organelles and proteins through degradation steps [33-35]. During these degradation steps, lysosomes release essential components synthesized from new molecules or associated with cellular signaling pathways via the diffusion or transport of proteins, such as the cholesterol transporters Niemann-Pick type C protein 1, 2 (NPC1, 2) and nicotinic acid adenine dinucleotide phosphate (NAADP) [36-38].
An acidic environment within lysosomes is critical for activating lysosomal enzymes and facilitating the conversion of their pro-forms into the mature forms necessary for degradation. Lysosomal enzymes involved in the degradation process are highly pH-dependent and are influenced by designated pH regulation for proper functioning. Disruptions in pH can interfere with substrate clearance, potentially linking diseases such as Parkinson's disease (PD) and MLIV, in which abnormal autophagy pathways contribute to lysosomal damage and improper protein aggregation.
Recently, lysosomes have been recognized as nutrient sensors that monitor cellular nutrient conditions and help maintain stable intracellular metabolism by adjusting various components.
THE ROLE OF ION CHANNELS IN MAINTAINING LYSOSOMAL FUNCTION
Emerging lysosomal ion channels: unveiling new therapeutic targets for lysosomal dysfunction
Lysosomal ion channels have only been discovered in the past decade, posing challenges in their study owing to the limitations of electrophysiological techniques. However, these channels play crucial roles in maintaining lysosomal homeostasis. By regulating ion channel activity, lysosomal dysfunction can be rescued, and promising therapeutic targets for diseases associated with impaired lysosomal function can be identified. Various techniques have been developed to investigate ion channel activity under different lysosomal conditions. Specific ions, such as Na+, K+, Ca2+, and Cl-, have been identified as key players in the maintenance of lysosomal homeostasis. Although the number of reported lysosomal ion channels is limited, their dysfunction has been strongly linked to impaired lysosomal function. This review focuses on the functions of lysosomal TPC, calcium-activated large-conductance K+ channel (BK), TMEM175, ClC-7, and TRPML channels (Fig. 1) and highlights their potential implications in lysosomal-related diseases.
Lysosomal sodium ion channels
TPCs in lysosomal function: TPCs are characterized by a dimeric structure with two-pore domains. Three types of TPC channels have been identified: TPC1, TPC2, and TPC3 (absent in mice, rats, and primates). TPC3 is currently under investigation because it localizes to the plasma membrane and is found in Na+-selective and Ca2+-permeable endosomal and lysosomal membranes [39-41]. TPC1, in contrast, is a voltage-dependent channel that is activated by PI(3,5)P2, the second messenger of NAADP, and changes in pH [41-45]. TPC2, although independently activated by voltage [40,42,46], has caused some problems regarding its activator. Some studies have suggested that TPC channels are activated by PI(3,5)P2 and increase Na+ influx [43], whereas others have proposed that NAADP activates Ca2+ influx, which is associated with TFEB translocation [47]. Structural studies conducted by Guo et al. [47] have indicated that TPC channels exhibit Na+ and Ca2+ sensitivity. PI(3,5)P2 has been shown to directly bind to TPCs and open channels, based on structural findings [45,48,49]. However, NAADP activation remains a subject of debate and requires further investigation, as it possibly differs among various organs. TPCs function as intracellular sensors during nutrient deprivation [43] and interact directly with mTOR, influencing channel activation and lysosomal function based on nutrient status. They are involved in pH homeostasis and membrane trafficking, affecting the pathogenicity of viruses, such as pigmentation-related diseases, Ebola, SARS, and COVID-19 [50-53]. TPC1 also plays a crucial role in the endosomes and participates in protein processing and uptake. Defects in TPC1 hinder toxin and protein uptake by weakening their interaction with syntaxins (SNARE syntaxins 7 and 8), leading to aberrant autophagy pathways due to lysosomal dysfunction from endosomes to lysosomes. TPC2 regulated lysosomal pH and activity. Overexpression of TPC2 led to an increase in lysosomal pH, suggesting its involvement in lysosomal homeostasis. Additionally, it modulates lysosomal membrane potential and channel activity [43,54]. Some studies have demonstrated that TPC2 knockout fibroblasts do not exhibit changes in lysosomal pH, but reveal increased lysosomal Ca2+ levels and reduced cholesterol degradation during low-density lipoprotein (LDL)-derived cholesterol transport [55,56]. Disruption of TPC activity can affect endosomal trafficking, thereby inhibiting viral spread [57,58], indicating a role for TPCs in lysosomal trafficking. Collectively, the regulation of lysosomal ion channels represents a promising novel therapeutic target.
Lysosomal potassium ion channels
Potassium influx helps maintain homeostasis by compensating for the loss of Na+ and Ca2+. Similar to the plasma membrane, the movement of K+ ions is closely linked to lysosomal membrane potential to ensure normal lysosomal function. Although the BK is believed to be the sole K+ channel in endolysosomes, a novel transmembrane protein, TMEM175, has been identified as a potassium-selective channel [13].
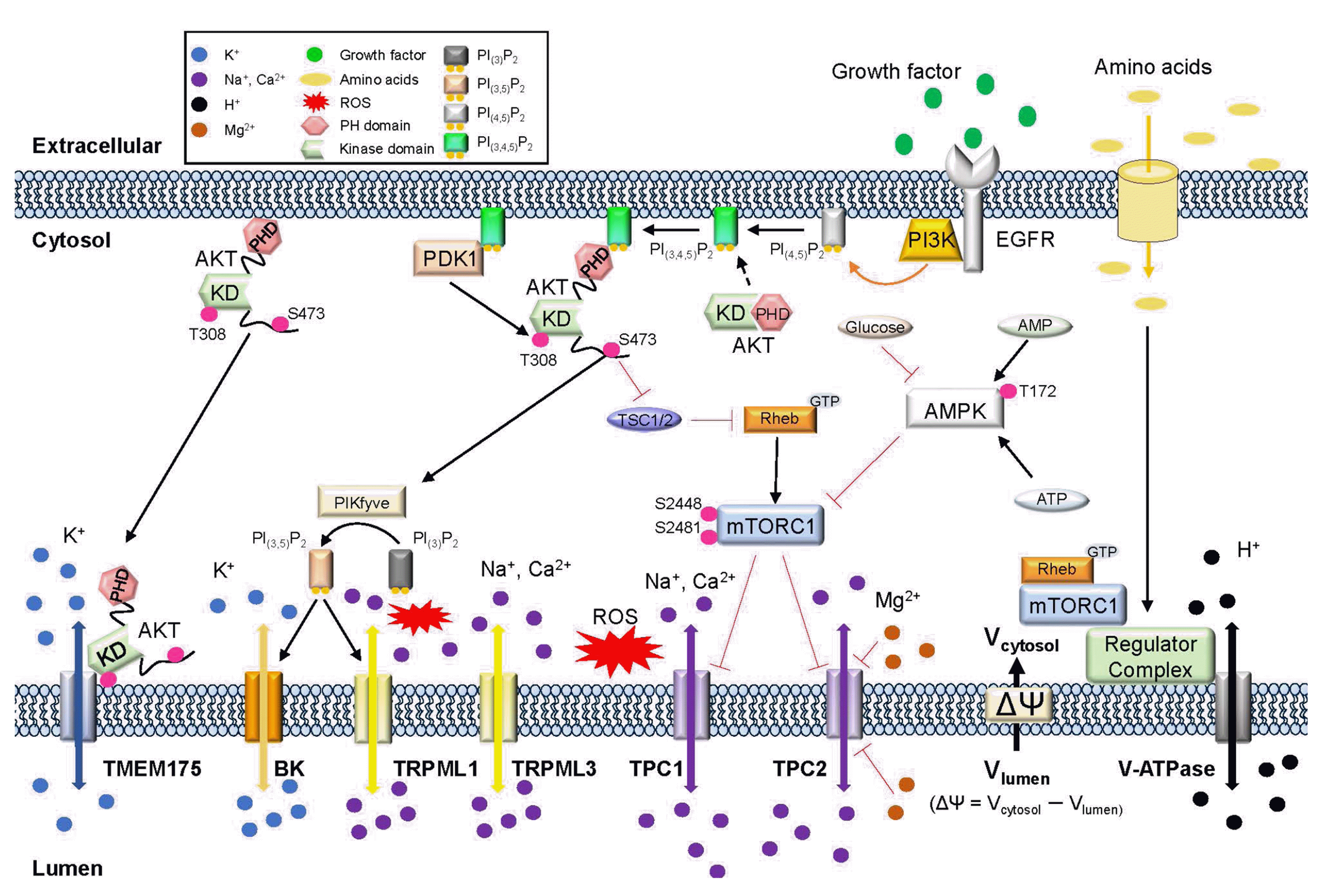
TMEM175: TMEM175 localizes within endolysosomes and was initially believed to exist only in prokaryotes. However, after the development of mass spectrometry, it was recognized as a mammalian protein [59]. TMEM175 acts as a K+ channel with the selective movement of K+ ions [60]. Unlike canonical K+ channels, TMEM175 does not inhibit tetraethylammonium (TEA) or quinine but suppresses Zn2+ and 4-AP. Although a P-loop selectivity filter is lacking in commercial K+ channels, structural studies have suggested that threonine and serine residues play a role in K+ selectivity [61]. Defects in TMEM175 have been found to regulate lysosomal pH and autophagy stages in RAW246.7 macrophage cells [60]. TMEM175 is strongly associated with neurodegenerative diseases like PD and may be involved in various LSDs [62,63]. It has emerged as a major protein connecting these diseases, as revealed by studies comparing Lewy body dementia patients and controls [62,64]. Additionally, TMEM175 deficiency affected mitochondrial function and autophagy in neuroblastoma cells [65] (Fig. 2).
BKs: In contrast, BK channels are calcium-activated large-conductance channels comprising six transmembrane segments and four pore-forming BK α-subunits (BKβ1-4). These channels were initially identified at the Slowpoke locus Slo1 in Drosophila [66]. BK channels not only respond to cytosolic Ca2+ but also influence membrane depolarization. Unlike other channels, BK channels are widely expressed and have been extensively studied in the plasma membrane. When activated, they facilitate K+ export from the cytosol, leading to hyperpolarization and increased lysosomal membrane. Some studies have suggested that BK channels are associated with TRPML1-mediated Ca2+ release. TRPML1 activation leads to Ca2+ release, which subsequently activates BK channels, allowing the influx of K+ ions into the lysosomal lumen [67]. This sustains the membrane potential necessary for the continuous activation of TRPML1 and V-ATPase. Inhibition of BK channels reduces lysosomal Ca2+ release, but co-treatment with a TRPML1 activator can rescue this effect.
Lysosomal calcium ion channels
TRPML channels: function, localization, and implications in lysosomal disorders: TRPML channels, initially discovered in Drosophila melanogaster, belong to the transient receptor potential (TRP) superfamily. These channels are nonselective cation channels that allow the permeation of calcium ions. The TRP superfamily comprises six main subgroups based on their functions: TRPA (ankyrin), TRPC (canonical), TRPM (melastatin), TRPML (mucolipin), TRPP (polycystin), and TRPV (vanilloid) [68,69]. TRPML isoforms include TRPML1-3, which are six transmembrane proteins with N- and C-termini. TRPML1 and TRPML2 are located on chromosome 19, whereas TRPML3 is located on chromosome 1 [70-72]. These isoforms share approximately 40% amino acid sequence similarity [73,74], and the localization of TRPML3 depends on the presence of TRPML1 and TRPML2. Controversially, it has been reported that both TRPML1 and TRPML2 bind to TRPML3 [75].
Among TRPML channels, TRPML1 is widely expressed [55], and localized in endosomes and lysosomes. The activation of TRPML1 channels leads to increased ion transport from the lysosomal lumen to the cytosol [76]. These channels are activated by PI(3,5)P2 and ML-SA1 but inhibited by ML-SL1. TRPML channels function within the lysosomes and interact with each other. TRPML1 has been extensively studied because of its association with LSDs. Mutations in TRPML1, also known as MCOLN1-3, are linked to LAMP-1 in the lysosomal membrane [77,78]. TRPML1 defects disrupt lysosomal conditions and lead to autophagic dysfunction and neurodegeneration. This highlights the importance of TRPML1 in organelle homeostasis and cellular function and suggests its role in lysosomal pH regulation. Interestingly, TRPML1 exhibited the opposite behavior of other ion channels. Dysfunction of TPCs and TMEM175 or nutrient deficiency increases lysosomal pH [13,42,79], whereas TRPML1 moves towards an acidic pH when depleted. TRPML1 plays a crucial role in sustaining the lysosomal function by interacting with lysosomal enzymes. However, some studies have suggested that pH can also influence TRPML channels [74]. TRPML1 contains an intraluminal pore with aspartate residues that modulate Ca2+ conductance depending on the surrounding pH [79].
TRPML2 and TRPML3: tissue localization and functions: TRPML2 exhibits tissue-specific expression and is primarily found in the heart, kidney, thymus, liver, immune cells, endosomes, and lysosomes [80]. Although the exact functions of TRPML2 channels are not well understood, some studies have suggested their association with glycosylphosphatidylinositol-anchored proteins (GPI-APs), a major histocompatibility protein class I, and ADP-ribosylation factor (ARF6), a small G protein involved in membrane trafficking and recycling. TRPML2 localizes to the ARF6-modulated pathway and regulates GPI-AP sorting [81].
In contrast, TRPML3 is found in various tissues, including the spleen, lungs, eyes, thymus, and melanocytes [82,83]. It is primarily localized in the endosomes, lysosomes, and intracellular vesicles [84,85]. TRPML3 plays a crucial role in regulating sodium ions (Na+) and cytosolic hydrogen ions (H+). However, it exhibited distinct pH regulation compared to TRPML1. The functional pH of TRPML3 is approximately 6.4, and its activity is inhibited at an acidic pH [85]. TRPML3 is present in multiple cellular compartments including the plasma membrane, endocytosis, and autophagy. Its activity is inhibited by acidic pH; however, when overexpressed or activated, it enhances autophagy and regulates its status. TRPML3 interacted with GATE16, an ATG8 homolog [86]. During autophagy induction, TRPML3 undergoes palmitoylation, a post-translational modification that facilitates the movement of specific components within the autophagosome, activating the process [87]. Previous studies have suggested a correlation between the number of autophagosomes and TRPML3 expression during cellular stress and nutrient deprivation. Cells lacking TRPML3 show reduced autophagosome formation, potentially leading to nutrient deficiency [88]. TRPML3 also interacts with autophagosome markers, indicating its direct involvement in the regulation of Ca2+-mediated membrane fusion. Structural studies have identified specific PIP2 binding site mutations (K52AR58AK62A, R305A, and F524A) that abolish TRPML3 currents, whereas K52AR58AK62A and R305A completely block PIP2 binding [83]. Constitutively activating isoforms of TRPML3, including the A419P and I362T mutants, have been reported, although the status of I362T remains controversial. Variants such as A419P are associated with conditions such as deafness and vascular problems because these mutations cause continuous Ca2+ release from lysosomes into the cytosol, leading to elevated cytosolic Ca2+ levels and disruption of intracellular signaling pathways. In the A419P mutant, sensory hair cell death results in hearing loss. Another mutation, I362T, reduced TRPML3 currents, resulting in a milder phenotype when co-mutated with A419P. Recent findings indicated that phosphatidylinositol-3-phosphate (PI3P), a crucial lipid for autophagosome formation, specifically interacts with TRPML3. The binding of TRPML3 to PI3P increases Ca2+ release from phagophores and promotes autophagy. Furthermore, TRPML3 inhibition abolishes autophagy, even in the presence of PI3P [89].
Lysosomal chloride-permeable channels: ClCs
The ClC family encompasses the voltage-gated Cl- channels that distinguish themselves from others by functioning as Cl-/H+ antiporters [90,91]. These channels are present in the plasma membrane and intracellular organelles. Depending on their location, ClC channels contribute to various processes, such as resting membrane potential, pH regulation, and overall homeostasis [90,91].
Among them, ClC-7 is widely expressed and prominently found in the central and peripheral nervous systems. It localizes to late endosomes and lysosomes, alongside Lamp-1 [92-99]. For its distinct functioning and localization, ClC-7 requires co-localization with the β subunit of osteoclastogenesis-associated transmembrane protein 1 [100,101]. ClC-7 is expressed at the ruffled borders of osteoclasts [92]. However, its specific role in the lysosomes remains debatable.
Recent findings have suggested that ClC-7 defects have a minimal impact on phagosomal acidification. Instead, ClC-7 primarily contributes to the accumulation of lysosomal Cl-, while V-ATPases are responsible for maintaining a low pH necessary for degradative hydrolase activity. ClC-7 indirectly activates V-ATPase by providing a driving force for Cl- accumulation [102].
Other studies have proposed that ClC-7 is predominantly associated with lysosomal hyperacidification, leading to the depletion of PI(3,5)P2 by inhibiting PIKfyve (an FYVE finger-containing phosphoinositide kinase) [65,103]. Notably, PIKfyve potently activates ClC-2 [104]. This suggests that ClC-7 is directly inhibited by PI(3,5)P2, thereby preventing functional mutations (such as Y715C) and regulating lysosomal pH [103,105].
LSDs linked to ion channels
LSDs are inherited metabolic disorders with a prevalence of approximately 1 in 5,000 live births [106], indicating the critical role of lysosomes in numerous cellular processes. Lysosomal dysfunction leads to various defects in lysosomal properties including pH imbalance, membrane potential disruption, enzymatic activity impairment, and abnormal autophagy. These lysosomal-mediated diseases encompass a broad range of conditions, from LSDs to neurodegenerative diseases, such as Parkinson's and Alzheimer’s disease, hearing loss, obesity, osteopetrosis, fatty liver, cancers (lung, breast, prostate, and colon), and viral diseases (Ebola and SARS) [107-118].
LSDs, which involve mutations or defective channel function, are common lysosomal dysfunctions. One such example is MLIV, a neurodegenerative and neurodevelopmental disease caused by mutations in TRPML1. Patients with MLIV exhibit dysfunctions in mTOR, trafficking, metabolism, lysosomes, and autophagy. Vardi et al. [119] conducted a study using TRPML1 knockout mice and obtained proteomic data through pathway analysis. The study revealed upregulated pathways in the cerebellum and cerebral cortex (integrin pathway, innate immune system, lysosome, and sphingolipid metabolism, among others) as well as downregulated pathways (GABAergic synapses). TRPML1 knockout mice displayed upregulated immune response, cytokine production, coagulation cascades, and viral susceptibility [119].
Niemann-Pick disease, another type of LSD, is associated with TRPML channels. This group of disorders causes cellular changes due to the accumulation of cholesterol and other lipids in endocytic compartments. In Niemann-Pick disease, cholesterol trafficking from the lysosome to the endoplasmic reticulum leads to dysfunction in transcriptional cholesterol metabolism and esterification. The improper regulation of lysosomal calcium release by TRPML1 mutations is thought to be associated with these issues [120].
Over the past few years, COVID-19 has become a pandemic, primarily causing respiratory syndromes related to SARS-CoV-2. While some individuals recover or improve naturally, others develop acute respiratory distress syndrome [121-123]. However, a few individuals may experience primary cardiovascular dysfunctions [124,125]. TPC channels, which localize in endosomes, lysosomes, and Na+ and Ca2+ channels, have been identified as potential candidates. It is suggested that TPCs regulate Ca2+ associated with SARS-CoV-2 through endocytosis and endolysosomal trafficking [50,126]. However, this hypothesis warrants further investigation. TPC1 is associated with systemic anaphylaxis and mast cell activity. TPC1-deficient mice exhibited enhanced passive systemic anaphylaxis, lower body temperature, slower recovery, reduced histamine release, decreased mast cell number and size, and impaired regulation of exocytosis [127]. TPC2, in contrast, modulates the endolysosomal degradation pathway by regulating trafficking. TPC2 dysfunction impairs epithelial growth factor (EGF)/EGF-receptor trafficking and LDL-derived cholesterol in mouse fibroblasts and hepatocytes. This indicates that TPC2 plays a role in regulating the trafficking mechanism of endolysosomal degradation pathways, which affect metabolite and macromolecule homeostasis. Recently, TPC2 has been shown to rescue lysosomal storage in MLIV and Batten disease. This channel activation improves the cellular phenotypes associated with lipofuscin accumulation, lactosylceramide buildup, cholesterol storage, and abnormal vacuole formation. TPC2 activation promotes lysosomal exocytosis and autophagy, providing a potential therapeutic avenue for these diseases [128].
Although BK channels were initially studied in the context of cancer, they have now gained attention for their use in neurological diseases. Mutations in KCNMA1, which encodes the pore-forming-subunit of BK channels, have been identified as crucial points. BK channel conductance plays a role in the regulation of breast cancer progression, and its localization and activation are linked to cancer development. Low expression of KCNMA1 has been associated with shorter recurrence-free survival in patients with breast tumors, regardless of treatment or estrogen receptor status. LRRC26, an auxiliary subunit, is required to activate BK channels in non-excitable tumor cells at a depolarized resting membrane potential [129]. In terms of neurological diseases, mutations in KCNMA1 (BKG354S) located in the selectivity filter and pore region have been linked to accelerated cognitive impairment in children with congenital and progressive cerebellar ataxia. These mutations significantly reduce ion selectivity and channel conductance and impair cell viability, mitochondrial content, and neurite outgrowth; however, they do not affect channel trafficking. However, the BK activator, NK1619, selectively blocks mutated BKG354S channels, suggesting a potential therapeutic strategy [130]. Another study proposed a correlation between KCNMA1 mutations and paroxysmal non-kinesigenic dyskinesia 3, which may occur with or without generalized epilepsy in mice and in three KCNMA1 patient variants. The BKN999S and BKD434G channels exhibited a gain-of-function phenotype, whereas the BKH444Q channel was negatively activated in heterologous cells. Heterozygous mutant mice with activating mutations show increased action potential firing and reduced seizure thresholds. However, this effect was not observed in mice harboring the H444Q mutation. N999S heterozygous mice demonstrate severe behavioral impairment in paroxysmal dyskinesia tests, suggesting its role in promoting seizure propensity (Table 1) [131].
TMEM175 has recently been implicated in PD. Unlike most ion channels that exhibit heterozygous activity, TMEM175 activation is dose-dependent. TMEM175 knockout mice display impaired clearance of phosphorylated α-synuclein (S129), a marker of PD, and increased galectin-3 levels, indicating lysosomal membrane damage. TMEM175 also functions as a nutrient sensor and its knockout negatively affects lysosomal pH, enzyme function, and autophagy. Furthermore, TMEM175 knockout mice were more susceptible to damage induced by starvation and neurotoxins (MPP+ and H2O2) than wild-type mice. Mutations in TMEM175 (M393T and Q65P) are associated with PD. The M393T mutation increased the likelihood of developing PD by more than 25%. Patients with PD and the M393T mutation experience decreased motor and cognitive functions. In vitro studies have shown that the M393T mutation reduces channel activity, impairs α-synuclein clearance, causes nutrient deficiency, and increases sensitivity to neurotoxins. In contrast, the Q65P mutation affected channel activity differently and was more resistant to nutrient deficiency. After deficiency, growth factors can rescue channel activity, indicating a connection between growth factors and TMEM175 channels. Protein kinase B (AKT), which is involved in multiple signaling pathways including cell proliferation, apoptosis, and metabolism, directly influenced TMEM175 activity through conformational changes in the AKT PH domain [13].
The industrial application of TMEM175 as a therapeutic target is supported by collaborations such as that between Caraway and AbbVie, which aim to develop therapies targeting TMEM175 with a funding allocation of up to $267 million. This highlights the potential significance of lysosomal ion channels as targets for therapeutic interventions. Another channel, TRPML1, has also been investigated as a potential target for agonist development to treat neurodegenerative diseases and muscular dystrophy. Merck has invested up to $576 million in this study owing to the close relationship between TRPML1 and lysosomal calcium. Reactive oxidative stress (ROS), TFEB, and GSK3 signaling are additional possible connections that warrant further investigation [132-138]. Reports have suggested that Mucopolysaccharidosis and Krabbe disease may be caused by ROS, indicating the potential involvement of TRPML and TPC channels. These channels can modulate lysosomal calcium release, affect local calcium levels, and potentially influence intracellular signaling. TFEB-mediated autophagy is linked to LSDs through the inhibition of mTOR pathways by drugs such as rapamycin. However, it has been observed that rapamycin can independently activate TRPML1 channels, leading to lysosomal calcium release and affecting TFEB. This interaction elucidates the coordination between lysosomal function and autophagy regulation. GSK3, a member of the AKT signaling pathway, is associated with familial Alzheimer's disease. Inhibition of GSK3 increases lysosomal activity, although its connection lies with plasma membrane ion channels rather than lysosomal ion channels.
The emerging understanding of lysosomal ion channels and their involvement in various diseases, such as LSDs and neurodegenerative conditions, opens exciting possibilities for the development of novel therapeutic strategies. Targeting lysosomal ion channels may provide a new approach to modulating lysosomal function, restoring cellular homeostasis, and alleviating disease symptoms.
Further research is required to elucidate the precise mechanisms and interactions of lysosomal ion channels with other proteins and signaling pathways. Investigating these complex networks can provide deep insights into underlying disease processes and identify novel therapeutic targets. Additionally, exploring the functional roles of lysosomal ion channels in different cellular contexts and organ systems can help to develop individualized and effective treatment approaches. Continuous research in this field holds significant potential for the development of innovative therapies that mitigate the impact of lysosomal dysfunction and neurodegeneration, thereby improving the quality of life of individuals suffering from these diseases.
CONCLUSION
Lysosomes play a crucial role in cellular signaling and maintaining cellular homeostasis. Dysfunctional lysosomes disrupt cellular homeostasis and contribute to the development of various diseases including neurodegenerative disorders and LSDs. Lysosomal ion channels, such as lysosomal membrane potential channels, are primarily responsible for regulating lysosomal conditions. The pathogenesis of these diseases has been linked to dysfunctions in these ion channels and their related signaling pathways. Therefore, a comprehensive understanding of the correlation between lysosomal ion channels, their regulatory mechanisms, and their impact on intracellular signaling is essential for the development of novel therapeutic approaches. Further investigations are highly recommended to unravel the precise roles of lysosomal ion channels and ion-related signaling in intracellular processes. In conclusion, this review aimed to provide valuable insights into the significance of lysosomal ion channels and ion-related signaling in intracellular signaling and their potential as therapeutic targets. A deeper understanding of the mechanisms involving lysosomal ion channels warrants continued research with the ultimate objective of developing effective therapies for LSDs and other related disorders.
 XML Download
XML Download