

 PDF
PDF Citation
Citation Print
Print



서 론
방사선 치료는 고형암의 치료에 있어 가장 효율적이고 중요한 치료이다. 모든 암의 50% 이상의 환자들이 방사선 치료를 받게 되고[1], 두경부 암의 경우 조기암의 경우 단독치료로, 진행암의 경우 항암치료와 병합하여 근치적 치료로 혹은 수술은 보조치료로 시행된다[2]. 방사선 치료에 대한 저항성은 방사선 치료 실패를 유발하여 잔존암을 만들거나, 치료 종료 후 재발이나 원격전이를 일으킬수 있는 중요한 요소이다[3]. 따라서 방사선 저항성에 기여하는 기전을 연구하는 것은 종양 재발을 줄이고 방사선 용량을 줄여 삶의 질 향상을 유도할 수 있다. 본 종설에서는 두경부암에서 방사선 저항성에 중요한 역할을 하는 것으로 알려진 다양한 신호전달체계 및 바이오 마커를 알아보고, 이중 중요한 치료 타깃이 될 수 있는 PI3K/AKT 및 EphA3의 역할에 대해서 자세히 기술하려고 한다.
Go to : 
DNA 손상 반응 및 복구
방사선 치료는 종양 DNA의 이중 나선을 파괴(Doublestrand break, DSB)함으로, 세포사멸을 유도한다. 하지만 암세포들은 DNA damage response (DDR)를 통해서 손상을 복구하고 세포생존을 촉진한다. DSB는 homologous recombination (HR) 또는 nonhomologous에 의해서 복구되고, HR은 정확한 DSB와 유전적 안정성 유지에 필수적이다[2,4,5]. DSB를 복구하는 기전은 방사선 저항성의 중요한 요소가 되며, DSB에 관여하는 XRCC2 (pretreatment protein expression) 의 발현이 DDR과 관련하여 불량한 예후인자와 관련되고(Fig. 1), 방사선 저항성을 예측할 수 있다고 알려져 있으나[6], 두경부암에서 그 역할은 아직 밝혀진 바가 없다. DDR은 매우 복잡한 신호전달체계들이 관련되어 있어, 임상적으로 이상적인 예측 바이오마커의 연구는 방사선 치료 반응을 예측하는 데 도움을 줄 수 있을 것이다.
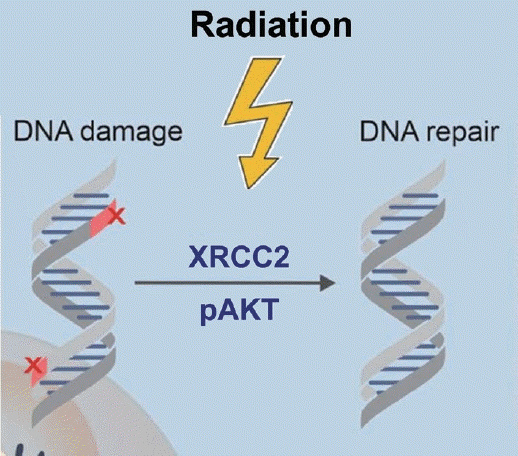 | Fig. 1.Irradiation results in DNA double strand break and in response to irradiation, DNA damage response is DNA repair is activated. Adapted from Larionova, et al. J Cancer Res Clin Oncol 2022;148(5):1015-31 [5].
|
Go to :
세포사멸 및 세포 포식
세포사멸이나 세포분열의 조절에 있어 중요한 tyrosine kinases (TKs)는 방사선 치료와 같은 스트레스 상황에서 세포가 이에 대응하여 생존하게 하는 역할을 한다[7]. TKs는 receptor tyrosine kinases (RTKs) (e.g., EGFR, PDGFR, EPHA, HER-2)와 non-receptor thyrosine kinase (NRTK: SRC, ABL, FAK, AKT)로 분류할 수 있다. 이들은 방사선에 의한 DNA 복구 및 세포생존에 필수적인 역할을 하여, 세포 내 발현이 증가되어 있는 경우 방사선 저항성 및 불량한 예후 그리고 재발 예측 바이오마커가 될수 있다[2,3,5,7]. 또한 TK inhibitor들은 다양한 표적치료제로 개발되어 사용되어 있다[7]. 그 외에 자가포식(autophagy)은 방사선과 같은 세포 스트레스 상황에서 세포 생존의 메커니즘이 될 수 있어, 자가포식이 과도하게 활성화되는 경우 방사선의 효능이 떨어져서 저항성의 원인이 되기도 한다. 이전 연구에서 두경부암 세포 및 동물 모델에서 지속적인 자가포식증상의 활성화가 방사선에 의해 세포 사멸이 유도되는 것을 막고, 방사선 저항성이 발생하게 되는 중요한 요인이 됨을 밝힌 바 있다[8].
Go to :
저산소증
Hypoxic cancer cells (저산소성 암세포)는 암세포의 공격성 및 방사선 치료에 대한 저항성을 유도하는 것으로 알려져 있다. Hypoxia inducible factor (HIF)는 저산소 상태에서 다양한 세포 생존에 필요한 여러 유전자들을 조절하는 중요한 단백질이며, α subunit (HIF-1α, HIF-2α and HIF-3α), 그리고 the β s ubunit (HIF-1β)으로 분류되며, HIF-1α와 HIF-1β 결합체가 저산소 상태에서 증가하는 중요한 전사인자이고, 저산소 상태를 나타내는 바이오마커이기도 하다. HIF-1α은 방사선에 의한 혈관 손상이나 reactive oxygen species (ROS)에 의해 증가하는 것으로 알려져 있다[9,10]. 다양한 암종에서 HIF-1α의 증가는 항암방사선에 대한 저항성 예측 및 예후 인자로 알려져 있다(Fig. 2). 또한, HIF-1α는 fibroblast growth factor and vascular endothelial growth factors 등의 신생혈관인자들을 자극하고 이를 통해 주변 조직의 침범을 유도한다[9,10]. HIF-1α와 달리 HIF-1β는 세포 내 에서 일정하게 발현되고, 세포 내에서 존재하고 HIF-1α의 결합 파트너이다. 추가적으로 HIF-2α도 저산소 상태에서 타깃이 될수 있는 mRNA의 전사를 촉진하는 역할을 하는 것으로 알려져 있다[11-13]. 하지만 이러한 저산소성 반응이 두경부암모델에서 방사선 저항성에 미치는 영향이 분명하지 않고, 이를 타깃으로 하는 약물의 유용성은 아직 명확하지 않으며, 환자의 종양에서 저산소성과 그렇지 않은 종양을 구분하는 방법에 대한 연구 또한 필요한 상황이다.
 | Fig. 2.Radiation promotes hypoxia inducible factor (HIF)-1 activation by reactive oxygen species (ROS) in hypoxic condition for activation of cell survival pathway. Adapted from Larionova, et al. J Cancer Res Clin Oncol 2022;148(5):1015-31 [5].
|
Go to :
종양미세환경
방사선 치료는 암세포뿐만 아니라 주변의 종양미세환경(tumor microenvironment, TME)에도 영향을 준다. 종양미세환경은 cancer-associated fibroblasts (CAFs), 내피세포, 면역세포, 혈관, extracellular matrix (ECM, 세포외부의 matrix ECM) 구조 등으로 구성되어 있다(Fig. 3). 암세포와 TME 구성요소들 간의 다양한 상호작용을 통해 종양의 성장, 혈관 재생, 방사선 저항성 획득 등을 촉진한다[14-16]. CAFs는 ECM 리모델링, 대사 및 혈관생성조절, 다양한 chemokine과 cytokine의 생성에 의해 암세포와 주변 면역세포와의 crosstalk 같은 다양한 기능을 하는 중요한 TME의 구성요소이다[15,16]. 또한, CAF는 DNA repair response 및 anti-apoptotic signaling pathway, epithelial mesenchymal transition (EMT)의 활성화를 통해 항암 방사선, 면역치료에 대한 저항성을 유발하는 중요한 역할을 하고, 이외에 암세포의 이동과 침습적 특성을 촉진하여 전이를 증가시키는 역할을 한다. 방사선 치료 후 증가된 CXCL-1, CXCL-12, FAP, TGF-β와 procollagen-lysine, 2-oxoglutarate 5-dioxygenase 3의 발현이 CAF의 증가를 나타내는 바이오마커이고 다양한 암에서 불량한 예후와 관련되어 있다[15,16]. CAF을 타깃으로 하는 항암기전은 일차적으로 특이적 표면 마커를 타깃으로 하여 CAF를 제거하는 것에 중점을 두고 있다. Fibroblast activation protein-α (FAP5)-DM1은 FAP와 CAF를 동시에 타깃으로 하는 monoclonal antibody로 동물실험에서 유의하게 종양 억제 혹은 완전 관해를 유도하였다. 그 외에도 CAF에 의한 하위 신호 물질들 예를 들면 IL-6, IL-6R, JAK/STAT3 등을 타깃으로 하는 약물들이 FAP와 CAP에 의한 다양한 cytokine들을 억제하기 위해 개발되고 있다. 병합치료로써의 다양한 역할이 기대되고 있으나, 현재 여러 임상시험이 진행 중이다[15,16]. Tumor-associated macrophages은 항암방사선 치료 후 전이나 종양의 재성장에 중요한 역할을 하는 innate immunity의 중요한 요소이다[17]. Macrophage의 종류는 macrophage 분극화(polarization)에 의해 그 타입이 나누어지고, M1 macrophage는 기본적으로 항원을 제시할 수 있는 능력을 가지고, 염증성 사이토카인의 활성화을 통해 암을 억제하는 능력을 가지는 반면, M2 macrophage는 항염증 성질을 가지며 항원을 제시하는 능력이 낮고, 암면역을 억제하며, 신생혈관형성, 저산소증 유도, 면역 억제, 세포증식 및 전이를 유도하는 알려져 있다. 이러한 macrophage의 분극화는 종양미세환경에 의해 조절되는 것으로 알려져 있다[18]. 또한, 기존의 연구들에서 M2 macrophage의 축적은 종양의 신생혈관 형성 및 macrophage의 정상적인 포식작용을 억제하여 방사선 치료 효과를 제한하는 것으로 알려져 있다[14]. 본 연구자들은 macrophage에 의한 포식작용을 억제하는 CD47의 발현이 방사선 저항성 모델에서 증가되어 있는 것을 발견하였고[19], 이는 종양 내 CD47이 발현증가가 미세종양환경 내 macrophage의 기능을 억제하므로, 방사선에 대한 저항성이 생길 수 있음을 시사하는 결과이다.
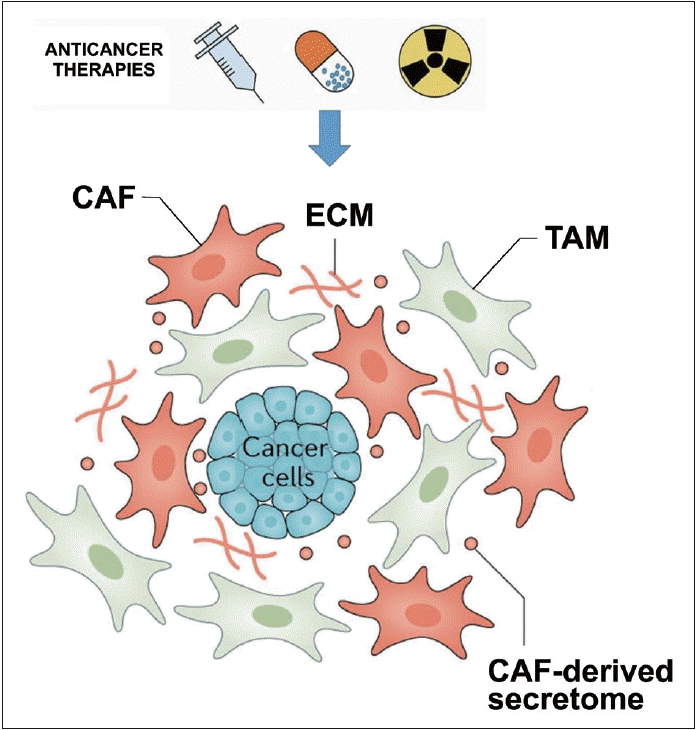 | Fig. 3.Anticancer therapy including radiation induces the change of tumor microenvironment such as cancer associated fibroblast (CAF), extracellular matrix (ECM) and immune cells and tumorassociated macrophages (TAM). Adapted from Chen, et al. Nat Rev Clin Oncol 2021;18(12):792-804 [16].
|
Go to :
PI3K/AKT pathway
Phosphatidylinositol-3-kinase (PI3-K)/protein kinase B(AKT) 신호전달체계는 방사선저항성을 획득함에 있어 내재적 저항성, 종양 세포 성장 그리고 저산소증의 3가지 메커니즘에 의해서 이루어진다. PI3K/AKT pathway는 두경부암을 포함한 고형암에서 활성화되어 세포의 성장과 침범, 세포사멸 억제 그리고 저산소 관련 단백질의 활성화 유도에 관여한다. 일반적으로 상위신호로는 EGFR과 같은 receptor t yrosine kinase에 의해 활성화되는 것으로 알려져 있다(Fig. 4) [20-25]. PI3K/AKT 신호체계는 두경부암의 90% 이상에서 활성화되어 있고, 이는 EGFR 활성화(47%), PIK3CA 돌연변이(8.6%), PIK3CA 증폭(14.2%), PI3K 과발현(27.2%) 등에 의해 유도된다[25]. 그외에 phosphatase and tensin homolog (PTEN)은 이 신호전달체계의 가장 강력한 억제 인자로, 두경부암의 15%에서 PTEN의 돌연변이가 있고, 다양한 전사인자로 인해 단백질 발현 저하가 있는 경우 phosphatidylinositol (3,4,5)-trisphosphate (PIP3)의 비활성화가 감소하여, AKT의 활성화가 유도되면서 항암 방사선 치료 저항성이 발생하는 것으로 알려져 있다[21]. Class IA PI3K는 a catalytic subunit (p110)와 a regulatory subunit (p85)으로 구성된 heterodimeric protein으로 p85 regulatory subunit은 phosphatidylinositol(4,5)-bisphosphate을 PIP3로 전환하는 역할을 통해 AKT를 활성화하는 기능을 한다[22,26]. 두경부암에서 AKT, PI3K 억제제를 이용한 임상연구는 현재 phase I I까지 진행되어 있다. 주로는 기존의 항암제나 cetuximab과의 병합 요법에 대한 연구로 진행되어 왔다. PI3K inhibitors는 2가지 종류가 있고, alpelisib (BYL-719), buparlisib (BKM-120), copanlisib (BAY 80-6946), LY294002, pilaralisib (XL147) and taselisib (GDC 0032) 등이 있으며, 대부분 pan-class l PI3K inhibitors이며, BYL-719은 p110α에 특이적인 inhibitors이다[21,27,28]. PI3K inhibitors는 이론상으로 보면 방사선 저항성을 극복할 수 있는 매력적인 약물이지만 실제로 고형암에서 PI3K inhibitors 단독 치료는 약물에 대한 저항성이 발생하여 실망스러운 치료 결과를 보이고 있다. 이러한 저항성 기전으로 PI3K pathway의 불완전한 억제나 재활성화 또는 보완적인 pathway의 활성화 등이 원인으로 생각되고 있다. 또한 in vitro나 in vivo 동물실험에서는 cisplatin, 5-FU, docetaxel 등과의 병합치료에서 효능이 있음이 증명되었으나, 환자에서의 사용시 잦은 부작용이 있으며, 두경부암에서는 아직 효과적인 결과를 확인하지 못하였다[21,27]. 이전 연구에서 방사선 저항성 두경부암에서 PI3K p85의 과발현이 중요한 역할을 함을 확인하였고, 흥미롭게도, PI3K p85 과발현이 발견되었으나 PI3K inhibitor에 저항성이 있음을 발견하였다. 이는 PI3K pathway에 수많은 crosstalk signal이 존재하기 때문에 PI3K 자체를 억제해도 다른 pathway의 활성화로 저항성이 발생하는 것으로 생각되었고, 두경부암에서는 Src의 지속적 활성화를 통한 PI3K/AKT pathway의 활성화가 유지된다는 것을 발견하여, 이를 근거로 Src inhibitor를 병합 처리하였을 때 PI3K i nhibitor에 대한 저항성이 극복됨을 확인하였다. 이는 p85의 활성화가 PI3K inhibitor에 대한 저항성을 예측할 수 있는 바이오마커가 될 수 있음을 시사하는 결과이기도 하다[29]. 또한 PTEN이 돌연변이에 의해 기능이 없어지거나 혹은 후성학적으로 억제되는 경우에도 PI3K의 활성화를 유도할 수 있다. PTEN의 유전자 돌연변이를 평가하는 것만으로 복잡한 조절기전을 완전히 밝혀내기가 어렵고, 유전자의 변화, 전사조절, 비암호화 RNA 등 다양한 조절 기전이 존재하는 것으로 알려져 있다[30-32]. 저자는 이전에 이러한 복잡한 조절 기전 중 후성 유전학적 기전에 초점을 두고 연구를 하였고, PTEN의 CpG hypermethylation과 H3K27me에서의 histone methylation 의 강력한 후성학적 억제를 통해서 방사선 저항성 두경부암 모델에서 PTEN의 상실이 발생함을 증명하였다[33]. 이러한 사실들을 통해서, PI3K/AKT signal 전달 체계의 복잡성 및 다양한 crosstalk signal의 존재가 PI3K 혹은 AKT inhibitors에 대한 저항성을 유발하게 되고, 단독 치료로써의 유용성의 한계를 가져오게 한다는 것을 알수 있다. 따라서, 방사선 저항성이 있는 두경부암에서 단독치료보다는 병합치료로써 역할을 기대할 수 있다.
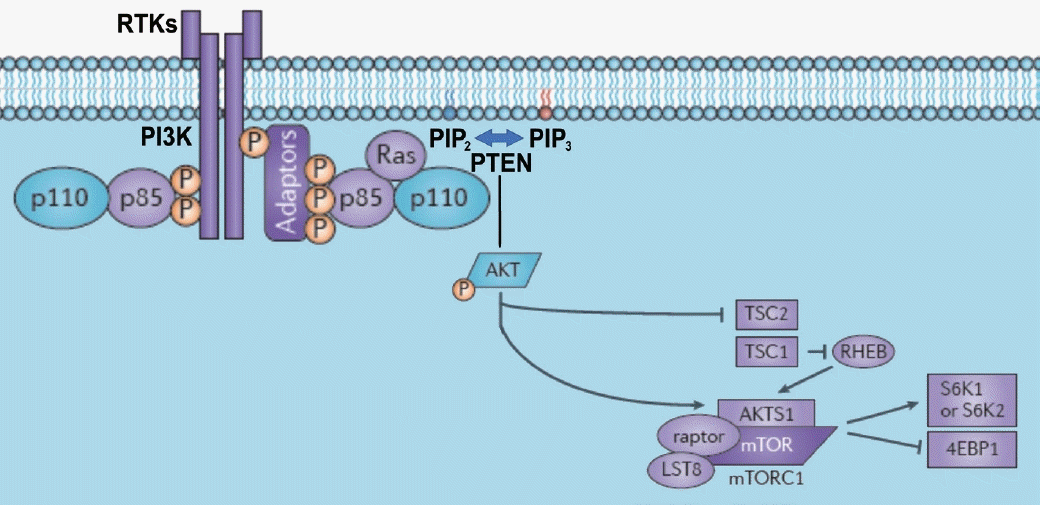 | Fig. 4.Activation of phosphatidylinositol-3-kinase (PI3-K)/protein kinase B (AKT) cascade in aspect of cancer aggressiveness and radioresistance. Adapted from Burris HA 3rd. Cancer Chemother Pharmacol 2013;71(4):829-42 [26].
|
Go to :
EphA3
Eph receptors는 RTK에 속하며, 세포 간 결합과 EMT를 유도하여 종양 진행, 주변조직 침범 및 전이에 중요한 역할을 하는 것으로 알려져 있고, 다양한 하위 신호체계를 조절할 수 있는 것으로 알려져 있다[34-39]. Eph receptor들은 2개의 하위분류(A and B)와 16개(A1-A10, B1-B6)의 멤버로 나누어진다. EphA3 (3p11.2)는 983개의 아미노산을 가진 막통과 단백질(transmembrane protein)로 교모세포종을 포함한 다양한 고형암에서 발현이 증가하여 공격적인 성향을 유도하고, 치료의 저항성을 유발한다고 알려져 있다[40,41]. 또한 agonistic α-EphA3 antibody를 사용하여 EphA3의 기능을 억제한 경우 종양의 실질 및 미세혈관 생성을 방해하여 종양성장을 효과적으로 억제할 수 있음이 보고되었다[42]. 두경부암에서 EphA3의 발현과 관련된 연구나 방사선 저항성에 관한 연구가 전무한 상황에서 저자는 EphA3의 역할을 연구하고 밝혀내었다[33,39]. 본 연구그룹은 두경부암 모세포에 2 Gy의 방사선을 70 Gy까지 매일 조사하고 이 과정에서 살아남은 세포들이 이전의 모세포와는 다른 방사선에 저항성을 가짐을 확인하여, 안정적인 방사선 저항성 세포주를 확립하였고 이에 기존 세포에 R 을 붙여 명명하였다[43]. 그리고 AMCHN-3 세포주와 AMC HN3R 세포주 간에 달라진 DNA 발현을 확인하고자 microarray를 시행하였다. 두 세포주 간에 방사선 용량에 따라 그 발현이 증가하는 몇 가지 물질을 발견하였고, 그중 방사선 저항성과 관련하여 EMT를 유도하는 중요한 물질이 EphA3임을 발견하였다. EphA3는 다양한 방사선 저항성 세포주에서 발현이 증가되어 있었으며, EphA3 siRNA를 이용하여 억제시 방사선 감수성이 증가하는 사실을 확인할 수 있다. 또한 이러한 방사선 저항성 조절에 가장 중요한 요인이 EMT를 조절함으로 기인된다는 것을 밝혀내어, EphA3가 방사선 저항성이 있는 두경부암에서 중요한 치료 타깃이 될수 있음을 확인하였다[39]. 이외에, EphA3가 PI3K의 활성화 물질인 p-AKT를 지속적으로 활성화시킴을 발견하고 그 기전에 PTEN의 억제에 중요한 역할을 함을 발견하였는데, DNA methylation과 histone methylation의 두 가지 기전을 통해 PTEN을 후성유전학적으로 강하게 억제하고 있었다[33]. PTEN의 강력한 억제는 AKT를 지속적으로 활성화시켜 방사선 저항성에 중요한 기전이 됨을 알 수 있었다(Fig. 5). 이런 두 가지 연구 결과는 EphA3가 방사선 저항성이 있는 두경부암 혹은 방사선 치료후 재발한 암에서 중요한 치료 타깃이 될 수 있음을 시사하는 결과이다. 또한 PI3K/AKT가 활성화되어 있는 암종에서 EphA3가 활성화되어 있는 경우 지속적인 PTEN 억제로 AKT 활성화가 강력하게 유도되므로, PI3K 혹은 AKT inhibitors들에 대한 저항성이 생기는 기전이 될 수도 있다. 따라서, 방사선 저항성 암종에서 표적 치료제를 고려할 때 EphA3의 활성화를 확인하여, 방사선 혹은 표적치료제에 대한 저항성을 예측하는 바이오마커로 이용가능하다. 최근 Eph/ephrin을 타깃으로 하는 약물들은 항체, 재조합 단백질 등이 있으며, 다양한 암종 동물 모델에서 효과가 입증되고 있다[44]. 현재 tyrosine kinase EphA3 수용체를 타깃하는 monoclonal antibody 인 KB004가 대표적으로 사용되고 있다. 이전 연구에서 EphA3 mAb IIIA4가 acute lymphoblastic leukemia 세포주와 동물모델에서 종양 성장과 침습을 억제하였고[45], 다발성 골수성 동물모델에서도 유의한 종양 억제를 보였다고 보고하였다[46]. 현재 EphA3 monoclonal antibody를 고형암에서 사용한 임상연구는 없으나, 혈액암에서 KB004 monoclonal antibody를 이용한 경우 1주일에 한 번씩 3주간 주사한 경우 급성백혈병 환자의 대부분에서 반응을 보이는 긍정적인 결과를 보고하였다[47]. 두경부암에서 방사선 저항성과 관련된 중요한 역할이 증명된 바를 바탕으로 방사선 치료와 병합 또는 추가적인 표적 항암제 혹은 면역치료제와의 병합치료를 통한 역할이 기대되는 바이다.
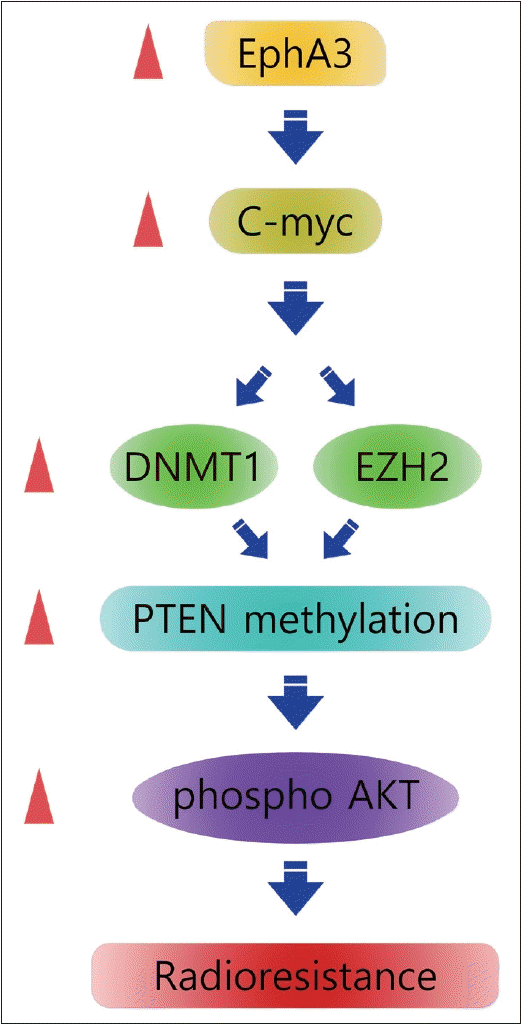 | Fig. 5.Phosphatase and tensin homolog (PTEN) expression is downregulated by CpG hypermethylation in the PTEN promoter and histone hypermethylation in H3K27me3 and results in constitutive activation of the protein kinase B (AKT) pathway in radioresistant head and neck cancer. Adapted from Kim, et al. Biomolecules 2021;11(4):599 [33].
|
Go to :
종양 줄기 세포
Cancer stem cell (CSC)은 방사선에 대한 세포 독성에 적응한 일부 세포군이 자가재생과 분화 능력을 가지고, 전이 및 종양재생 그리고 항암방사선에 대한 저항성을 갖게 되는 것을 일컫는다. 두경부암에서 stem cell 마커로, CD44, ALDH, 그리고 CD133이 밝혀져 있고, 여러 임상 연구에서도 방사선 저항성에 중요한 역할을 하는 것으로 알려져 있다[2,48]. 임상적으로 CD44의 발현증가가 방사선치료 후 재발이나 치료 실패를 예측할 수 있는 위험 인자가 됨이 보고되었다[49]. CSC는 DDR, 활성 산소(ROS), 그리고 Wnt, Notch 등 여러가지 시그널을 통해서 방사선 저항성을 획득하는 것으로 알려져 있다[26]. 방사선 저항성에서의 CSC의 역할은 추가적인 연구가 필요한데, 다양한 세포군의 표현형과 기능적 특징이 다양하기 때문에 그 차이에 따른 방사선 저항성에서의 역할 규명이 해결해야 할 문제이다.
Go to :
결 론
방사선 저항성은 두경부암 환자의 예후에 가장 큰 영향을 주는 요인으로 방사선 저항성은 하나의 핵심 조절 물질이나 신호전달체계만으로 설명하기 어려운 복잡하고, 다양한 유전자들의 변이에 의한 기능의 상실 혹은 획득으로 이루어진다. 가장 대표적인 신호전달체계가 EGFR, EphA3, PI3K/AKT 이지만, 최근에는 이러한 신호전달체계뿐 아니라 방사선에 의한 종양 미세환경의 변화 즉 CAF의 증가 등이 중요한 요인으로 거론되고 있다. 즉 두경부암에서 방사선 저항성에 기인하는 메커니즘은 너무나 많고, 최근에 많은 방사선 감수제들이 연구가 되고 있지만, 아직까지도 많은 치료제들이 임상 단계에서는 큰 효과를 보지 못하고 있는 실정이다. 최근에는 암 세포의 면역환경들의 변화도 방사선 저항성과 관련되어 연구들이 이루어지고 있으며, 아직 획기적인 방사선 저항성 극복 방안이 없는 상황에서, 방사선 저항성에 대한 깊이 있는 연구들이 이루어져야 할 것이다.
Go to :
 XML Download
XML Download