

 PDF
PDF Citation
Citation Print
Print


Introduction
During the process of orthodontic tooth movement (OTM), the periodontal ligament cells (PDLCs) sense the mechanical signals and then translate them to biological signals (1). Li et al. (2) summarized that the receptors, channels, cellular signal pathways, intranuclear signal pathways, and some new mechanisms, including the myofibroblast, noncoding RNAs, hypoxia, and autophagy are involved in mechanical signal transduction in PDLCs. Tension strain-stimulated PDLCs can not only differen-tiate into osteoblasts to synthesize bone tissue (3) but also synthesize and release cytokines, growth factors, neurotransmitters and colony-stimulating factors (4-6). These factors may regulate the cell migration, proliferation and differentiation of other types of cells in the microenvironment (3, 6). Orthodontic mechanical loading can accelerate transplanted bone marrow cell migration into periodontal tissues in mice (7). Bone marrow mesenchymal stem cells (BMSCs) can migrate to the tension side of the orthodontic moving tooth and differentiate into osteoblasts (8, 9). In a previous summary, human MSCs migrate in response to some chemokines, cytokines, growth factors and bioactive peptides such as SDF-1, MCP-1, RANTES, MIP-1α, IP-10, IL-8, HGF, C1q, C3a and C5a (10). To study the effect of tension on PDLCs, Flexcell is the most widely adopted method to apply intermitted (“dynamic”) tension force to cells via the substrate deformation in vitro (11). Previously, we also proved that tension-stretched PDLCs could release SDF-1 into the medium to recruit BMSCs (9). However, the specific mechanism of BMSC recruitment in orthodontic treatment remains unclear.
There are some other intercellular communication factors, such as microvesicles (MVs) and exosomes (12). Exosomes are cell-derived vesicles with a diameter of 40∼200 nm and are essential for intercellular communication (13). PDLCs can secrete exosomes to regulate angiogenesis and osteogenesis as well as affect the inflammatory microenvironment (14, 15). Exosomes can help recruit BMSCs into a specific microenvironment (16) through enter BMSCs and affect their response to chemokines, thus, enhance the migration ability (17). Exosomes derived from Schwann cells can promote the migration, proliferation, and differentia-tion of BMSCs in bone repair (18). The function of exosomes depends on their contents. The contents of exosomes, such as protein, RNA, and DNA, are of importance for the biological effects mediated by exosomes (19).
miRNAs are small non-coding RNAs (∼22 nucleotides long). In the miRNA/mRNA interaction network, one miRNA can bind to multiple mRNAs and regulate multiple signal pathways (20). It has been reported that mi-RNAs are highly enriched in extracellular vesicles (21). The miRNAs in exosomes are important components for cell-to-cell communication (22) and may vary with culture conditions (23). miR-155 and miR-146a are released from active dendritic cells in exosomes and are taken up by recipient bone marrow dendritic cells. The transfer of these two miRNAs can mediate target gene repression and alter the ability of recipient cells to respond to a pro-inflammatory stimulus (24). Endothelial progenitor cell-derived exosomes can enhance proliferation and migration of the endothelial cells to the damaged site and accelerate reendothelialization (25). We sought to elucidate the role of stretched PDLCs-derived exosomes in regulating BMSC migration and the changes in exosomal miRNA profiles.
In this study, we isolated and identified PDLC-derived exosomes. We explored the effect of the exosomes from stretched PDLCs on the migration of BMSCs. We then identified the alternation of miRNAs in stretched-PDLC-derived exosomes, core proteins and key signal pathways in this process through a microarray and a series of bioinformatics analyses. This study will aid us to elucidate the mechanisms of BMSC migration affected by stretched PDLC-derived exosomes, which may provide further insight into the improvement of orthodontic treatment.
Materials and Methods
Cell culture
Primary human PDLCs (hPDLCs) were isolated from healthy PDL tissues of premolars extracted from 8 patients aged 11∼15 years undergoing orthodontic treatment, as described previously (26). This study was approved by the review board of the local hospital ethics, and written informed consent was obtained from each participant. Briefly, periodontal ligament tissues were carefully scraped from extracted teeth and cut into pieces. Tissues were further digested in a solution containing 3 mg/ml collagenase type I (Sigma-Aldrich) and 4 mg/ml dispase II (Sigma-Aldrich) for 1 h at 37℃. The digest was then filtered through a 70 μm cell strainer (BD Biosciences, Franklin Lakes, NJ, USA). Cell pellets were resuspended and cultured in alpha-modified Eagle’s medium (α-MEM, HyClone, Logan, UT, USA) supplemented with 20% foetal bovine serum (FBS) and 1% penicillin-streptomycin solution (HyClone). hPDLCs between passages 5 and 10 were used.
Bone marrow mesenchymal stem cells were purchased from Cyagen Biosciences (HUXMF-01001, Santa Clara, CA, USA) and cultured according to the manufacturer’s instructions. BMSCs at passages<5 were used for all ex-periments.
Application of cyclic tension stretch (CTS)
CTS imitates the tension force loaded on PDLCs during OTM (27). CTS was applied to hPDLCs following the methods reported previously (28). Briefly, hPDLCs were seeded into BioFlex six-well plates (Flexcell International Corp, Hillsborough, NC), which have flexible bottoms and are coated with type I collagen. Custom-built equipment was used to apply deformation to the flexible bottom. When reaching 80% confluence, PDLCs were washed three times with PBS and incubated in fresh exosome-free and serum-free α-MEM. CTS (5% elongation rate) was applied to hPDLCs at 0.1 Hz (5 s elongation and 5 s rest) for 48 h. Cells that were cultured with an identical protocol but without stretch served as the control.
Exosome isolation and characterization
Conditioned medium (CM) collection: After the stretch procedure, cell culture conditioned mediums (CMs) were collected for the extraction of exosomes. Microvesicles (MVs) and exosomes were isolated by differential centrifugation, which is the most commonly used method (29). First, CMs were centrifuged at 300×g for 10 min and at 2,000×g for another 10 min to remove cellular debris. Then, the supernatant was centrifuged at 10,000×g for 70 min at 4℃ to separate MVs (Beckman J20XP centrifuge, JA25.5 rotor, Indianapolis, IN, USA). Exosomes were then extracted by ultracentrifugation at 120,000×g for 110 min at 4℃ (Beckman Optima L-100XP ultracentrifuge, SW32Ti rotor). The pellets containing MVs or exosomes were resuspended in PBS. The supernatant after 10,000×g centrifugation and 120,000×g ultracentrifugation were also collected for further study.
Transmission electron microscopy (TEM): The morphology of exosomes was observed by transmission electronic microscope. Exosomes were resuspended in 100 μl of PBS. Then, a 10 μl suspension was placed on a cold and clean Parafilm. A 230 mesh Formvar carbon-coated grid was floated on the drop for 3 min with its coated side facing the suspension. Then, the grid was washed with 20 μl of distilled water for 2 min and stained with 20 μl of 2% phosphotungstic acid solution for 3 min. Thereafter, the grid was left to dry and immediately imaged by a TEM (Tecnai, FEI, Hillsboro, OR, USA).
Nanoparticle tracking analysis (NTA): NTA was used to measure the particle size and concentration of exoso-mes. The samples were diluted to the recommended concentration and loaded into the ZetaView PMX 120 (Par-ticle Metrix, Meerbusch, Germany). Each sample was shot three times for measurement using the corresponding software (ZetaView 8.04.02).
Western blotting analysis: PDLCs, exosomes, MVs, and supernatant after ultracentrifugation were lysed with RIPA Lysis Buffer. The concentration of the protein was measured by a bicinchoninic acid protein assay kit (Pierce, Thermo Fisher Scientific, Rockford, IL, USA). Equal amounts of proteins (20 μg) were loaded onto 10% Bis-Tris gels and separated by SDS-PAGE then transferred to polyvinylidene fluoride membranes (Millipore, Bedford, MA, USA). The membranes were blocked with 5% skim milk in 1×TBST solution and then incubated with each of the following primary antibodies overnight at 4℃: anti-CD63 (1:1,000, ab134045, Abcam, Cambridge, UK), anti-CD9 (1:1,000, ab92726, Abcam), anti-CD81 (1:1,000, ab109201, Abcam), anti-Alix (1:1,000, ab186429, Abcam), anti-TSG101 (1:1,000, ab125011, Abcam), anti-Calnexin (1:1,000, CST #2679, Cell Signalling Technology, Danvers, MA, USA), and anti-Vimentin (1:1,000, ab92547, Abcam). Immunoreacted bands were visualized using HRP secondary antibodies and the ECL method.
Exosome uptake assay: Exosomes were labelled by the PKH26 Red Fluorescent Cell Linker Mini kit (Sigma-Aldrich) according to the manufacturer’s instructions. Exosomes were then recollected using the ExoQuickTM Exosome Precipitation Solution (System Biosciences, Mo-untain view, CA, USA) and resuspended in basal medium. Meanwhile, BMSCs was stained with carboxyfluorescein diacetate succinimidyl ester (CFDA-SE) (eBioscience, Thermo Fisher Scientific, Rockford, IL, USA) and cultured overnight. Then, the culture medium was changed to basal medium with exosomes (100 μg/ml). After 1, 2, and 6 h of incubation, cells were gently washed in PBS and fixed in 4% paraformaldehyde for 20 min then stained with DAPI (Beyotime, China). A laser scanning confocal microscope (Leica TCS SP8, Leica Ltd, Germany) was used to capture images.
Cell viability assay: BMSCs were seeded in 96-well plates at the density of 4×103 cells. The cell viability was evaluated by the colorimetric MTS assay using CellTiter 96Ⓡ Aqueous One Solution Reagent (Promega, Madison, WI). Cells were treated with exosomes derived from cyclic tension stretched PDLCs (Exos_CTS, 30 μg/ml), unstretched PDLCs (Exos_US, 30 μg/ml) or PBS. After culturing for 2 days, the cells were incubated with 20 μl MTS reagent for 2 hours at 37℃. The optical density of supernatant was recorded at the wavelength of 490 nm by a plate reader (Agilent Technologies, Santa Clara, CA, USA).
Cell migration assay: Transwell assays were set up in 24-well plates using inserts with membranes of 8 μm pore size (3422, Corning, NY, USA) and split into two groups. In the first group, 4×104 BMSCs was seeded into the upper chamber. The CMs before centrifugation (CM_B), after 10,000×g centrifugation (CM_C), and after 120,000×g ultracentrifugation (CM_UC) were placed into the bottom wells. In the second group, 500 μl of α-MEM supplemented with 2% FBS was placed into the bottom well. BMSCs was pretreated with 100 μg/ml exosomes derived from unstretched PDLCs (Exos_US) and stretched PDLCs (Exos_CTS) for 4 h. Then, BMSCs was seeded into the upper chamber. After 20 h, all insert membranes were fixed with ethanol for 15 min. Non-migrated cells on the upper side were gently wiped off. Membranes were dyed with Crystal Violet Staining Solution and washed three times with distilled water. Migrated cells were observed and quantified in nine random fields of view on each membrane using a microscope (OLYMPUS BX51, Japan).
RNA interference: SiRNAs of Drosha and a scrambled negative control (NC) siRNA were designed and synthesized by Genepharma (Shanghai, China). siRNA against human Drosha (sense strand 5’-GGGAGAUUCUACAGU-GGUUTT-3’ and antisense strand 5’-AACCACUGUAG-AAUCUCCCTT-3’) or negative control siRNA (sense stra-nd 5’-UUCUCCGAACGUGUCACGUTT-3’ and antisense strand 5’-ACGUGACACGUUCGGAGAATT-3’) were transfected into PDLCs at a final concentration of 10 nm or 20 nm by using Hiperfect Transfection Reagent (Qiagen, Hamburg, Germany). After 48 hours, the transfection efficiencies were validated by Western blot analysis.
miRNA expression profiling: RNA in exosomes of each sample was isolated using the Qiazol RNA Isolation Kit (Qiagen). There were three biological replicates of CTS-PDLC-derived exosomes and unstretched PDLC-derived exosomes. Total RNA was quantized using the NanoDrop ND-2000 (Thermo Scientific) and RNA integrity was detected by the Agilent Bioanalyzer 2100 (Agilent Technologies). The miRNA microarray was performed using an Agilent miRNA Array (Agilent Human miRNA Microarray Kit, Release 21.0,8x60K; DesignID:070156) that can detect 2,570 human mature miRNAs (OE Biotech, Shanghai, China). Analyses were performed by OE Biotech. The original hybridization images were scanned by the Agilent Scanner G2505C (Agilent Technolo-gies) and analysed using the Feature Extraction software (Version 10.7.1.1, Agilent Technologies). Quantile standardization of the original data was then performed. Differentially expressed miRNAs were then identified through fold-change as well as the p-values calculated by Student’s t-test, and the criteria were upregulated or downregulated based on the change value >2.0 with p-value<0.05. A Volcano plot and cluster analysis plot were also drawn.
Bioinformatics analysis
Predicting target genes: The target genes of miRNAs were predicted using public prediction tools miRWalk3.0 and miRDB. The criteria were binding probability of 1 in miRWalk and prediction score greater than 80 (total score 100) in miRDB. Finally, the intersection genes of the two databases were obtained for subsequent analysis.
Protein-protein interaction (PPI) network and modules analysis: STRING (string-db.org/) is a web-based tool for the evaluation of PPI network information. The potential relationship among the predicted genes was detected by STRING and imported into Cytoscape software. A confidence score of >0.4 was set as the cutoff criterion. The Molecular Complex Detection Program (MCODE) in Cytoscape was used to display modules of the PPI network with node score cutoff=0.2, k-core=2, max. Depth from seed=100, and degree cutoff=2. Proteins in the top four interaction networks were then imported into STRING, and the interactions involved in each protein were counted to obtain the top 20% of proteins that had the most interactions in the network, named the hub proteins.
Gene Ontology (GO) and the Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis of hub genes: GO enrichment analysis is a common method for annotating genes. The KEGG is a database used to conduct searches regarding biological pathways, diseases, drugs, and chemical substances. The functional annotation of these hub genes was analysed using the ClueGO functional module in Cytoscape and was used to test the statistical enrichment of the hub proteins in the KEGG pathway and GO enrichment analysis.
Statistical analysis: Each experiment was performed three times. Results were described as mean±SEM. SPSS 16.0 software was used for statistical analysis using Student’s t-tests and one-way analysis of variance. p<0.05 was considered statistically significant.
Results
Characteristics of PDLC-derived exosomes
The ultrastructure of the PDLC-derived exosomes was detected by the TEM, which showed a cup-shaped morphology with a lipid bilayer (Fig. 1A). The average diameter of the vesicles was approximately 130 nm (Fig. 1B and 1C). The western blot results showed that the vesicles from ultracentrifugation expressed exosomal surface mar-kers CD63, CD81, CD9, ALIX, and TSG101 and were negative for the nonexosomal marker calnexin and PDLC marker vimentin (Fig. 1D).
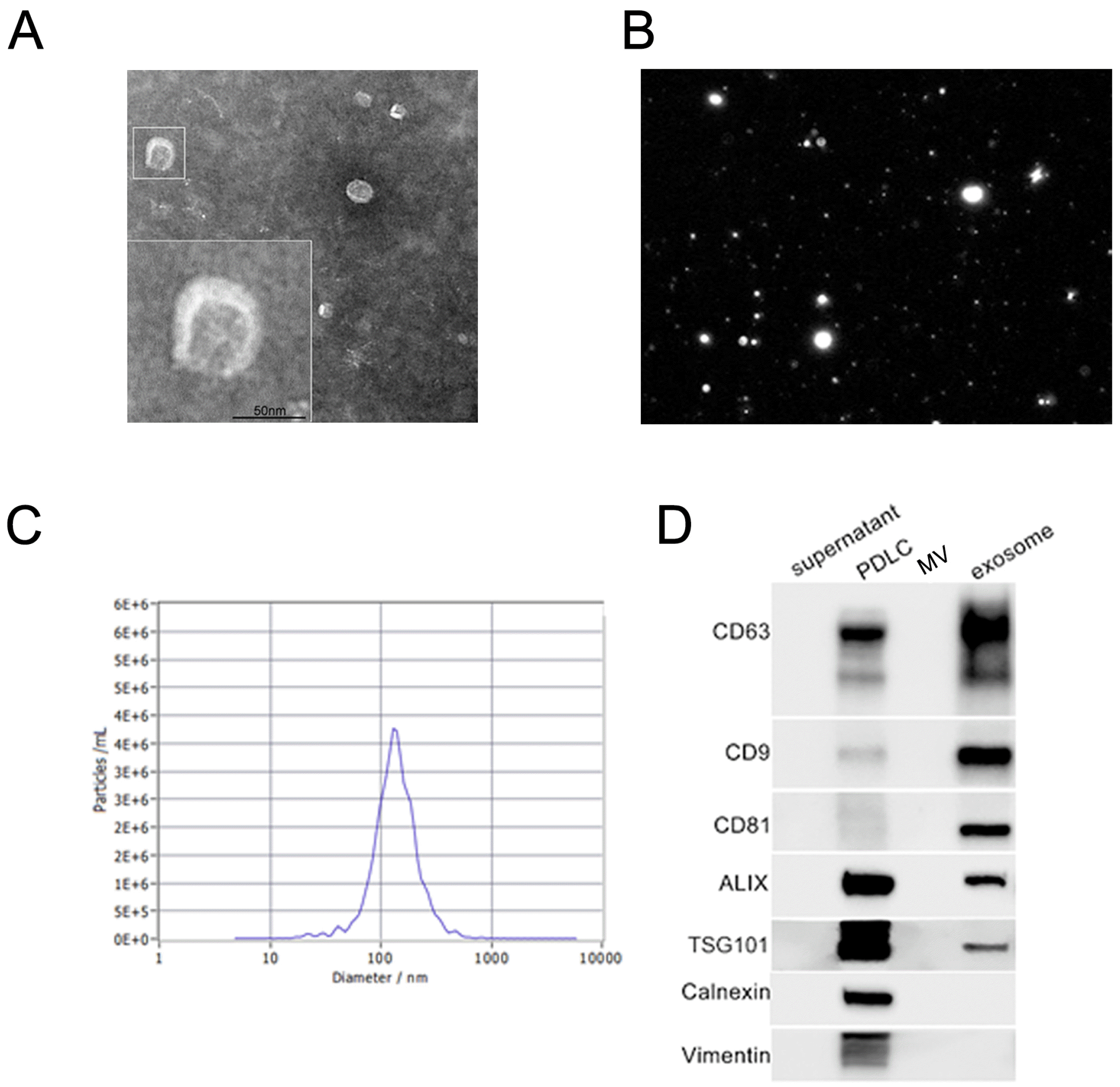
Fig. 1
Characterization of exosomes derived from human periodontal ligament cells (PDLCs). (A) Transmission electron microscope images confirmed the presence of exosomes, seen as cup-shaped vesicles. Scale bar: 50 nm. The picture in the white square is an enlarged photo. (B, C) The particle size distribution and concentration of exosomes were measured by nanoparticle tracking analysis. (D) Western blot analysis of the exosome-specific markers (CD63, CD9, CD81, ALIX, and TSG101), endoplasmic reticulum protein (calnexin), and fibroblastic marker (vimentin). PDLCs, microvesicles, and supernatant were used as the control.
![]()
Internalization of exosomes by BMSCs
Fluorescence microscopy analysis showed that PKH26-labelled exosomes (red dots) could be detected in the cytoplasm (green) of BMSCs after incubation (Fig. 2). Com-pared to 1 h of incubation, more exosomes were inter-nalized in BMSCs after 2 and 6 h of incubation.
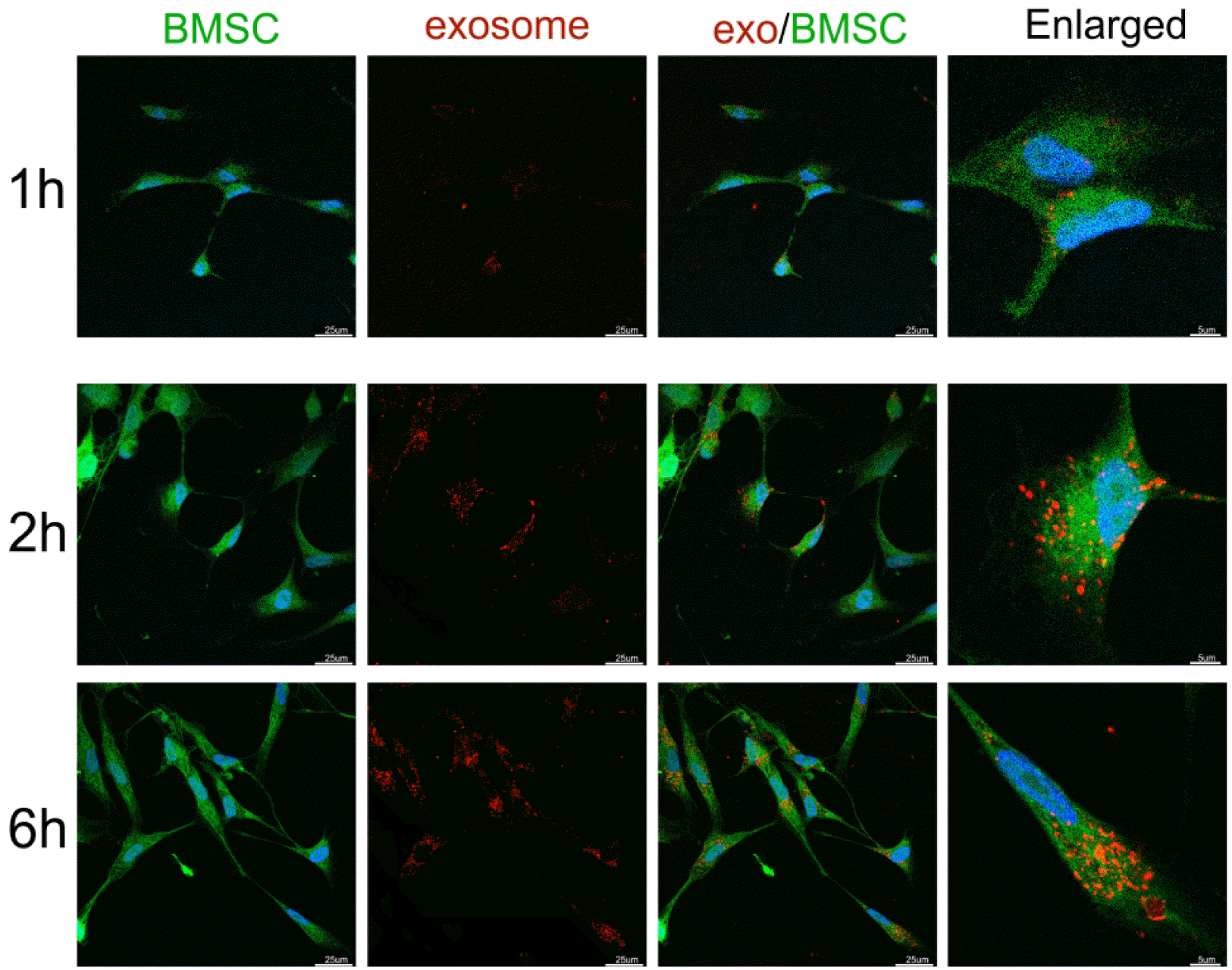
Fig. 2
Internalization of exosomes by bone marrow mesenchymal stem cells. PKH-26-labelled exosomes (red) were internalized in the cytoplasm of BMSCs after 1 h and accumulated after 2 and 6 h of coincubation. The nuclei of the BMSCs were stained with DAPI (blue). The cytoplasm of the BMSCs was stained with CFDA-SE (green). Scale bar: 25 and 5 μm.
![]()
The effects of CTS on the PDLCs-derived exosomes
Some membrane marker proteins’ expressions but not BMSC uptaken rate of exosomes were altered by CTS application. As shown in Fig. 3A, Western blot results showed that the expression of CD63 and CD81 increased in Exos_CTS compared with Exos_US, while Alix and TSG101 showed no significant difference between these two groups, and calnexin and vimentin were not detected in them. We also observed the internalization of Exos_CTS and Exos_US by BMSCs after 2 h of coincubation. Approximately equal amounts of of Exos_CTS and Exos_US were taken up by BMSCs (Fig. 3B).
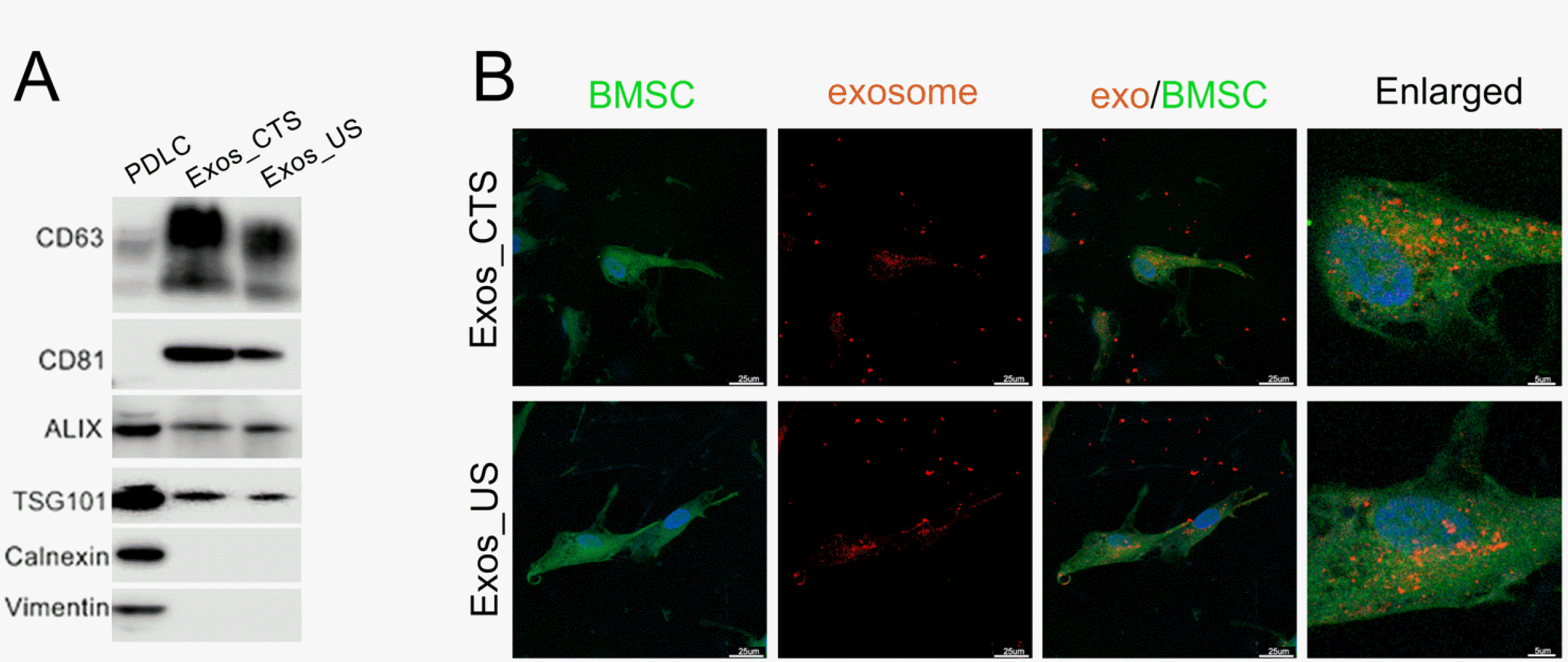
Fig. 3
Exosomes were collected from CTS-applied PDLCs (Exos_CTS) and unstretched PDLCs (Exos_US). (A) Western blot analysis of CD63, CD81, ALIX, TSG101, calnexin and vimentin in Exos_CTS and Exos_US. PDLCs were used as the control. (B) Internalization of Exos_CTS and Exos_US by bone marrow mesenchymal stem cells. PKH-26-labelled exosomes (red) were internalized in the cytoplasm of BMSCs after 2 h of coincubation. The nuclei of the BMSCs were stained with DAPI (blue). The cytoplasm of the BMSCs was stained with CFDA-SE (green). Scale bar: 25 and 5 μm.
![]()
The effects of exosomes from CTS-applied PDLCs on the migration of BMSCs
The cell viability of BMSCs showed nonsignificant change between the groups with Exos_CTS and Exos_US (Supplementary Fig. S1). The Transwell assays showed that the migration activity of BMSCs exposed to Exos_CTS was enhanced compared to BMSCs exposed to Exos_US (Fig. 4A). Further, there was no significant difference in BMSCs migration between the groups with CM before centrifugation (CM_B, containing MVs and exosomes) and after centrifugation at 10,000×g (CM_C, containing exosomes but not MVs). However, a significant decrease was observed in the group with CM after centrifugation at 120,000×g (CM_UC, no MVs or exosomes) (Fig. 4B). Removing MVs from the medium did not alleviate BMSC migration. However, further removal of the exosomes weakened their functions. Therefore, BMSC migration is affected by exosomes but not MVs derived from CTS-applied PDLCs.
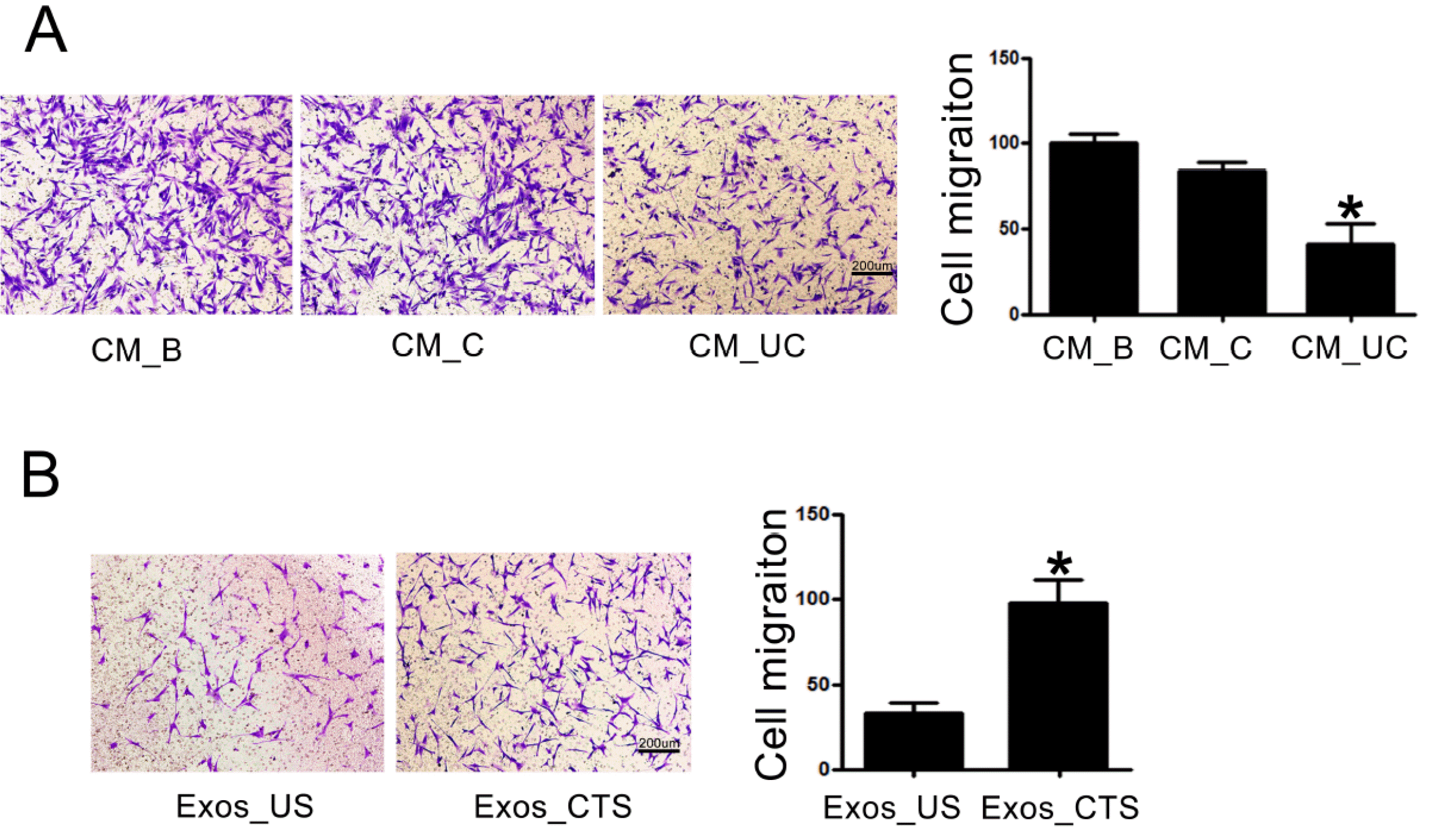
Fig. 4
Periodontal ligament cells (PDLCs) were applied 48 h after cyclic tension stretching. (A) Conditioned medium (CM_B) was collected from the cyclic tension stretch (CTS)-applied PDLCs and removed microvesicles (CM_C) and exosomes (CM_UC). BMSCs were stained with Crystal Violet solution. Representative images and the migration rate of BMSCs for different CMs. (B) Exosomes were collected from CTS-applied PDLCs (Exos_CTS) and unstretched PDLCs (Exos_US). Representative images and migration rate of migrated BMSCs treated with different exosomes. *p<0.05.
![]()
miRNAs are the key functional components in CTS-PDLC-exosomes to enhance BMSC migration
To demonstrate the function of miRNAs in CTS-PDLC-exosomes, we conducted small interfering RNA (siRNA) knockdown of Drosha (an essential polymerase for miRNA synthesis) in PDLCs. When siRNA was used to deplete Drosha in CTS-PDLCs (Fig. 5A), the exosomes had less effect to enhance BMSC migration in transwell than siRNA negative control group (Fig. 5B). The result indicates that miRNAs are responsible for the ability of CTS-PDLC-exosomes to modulate BMSC migration.
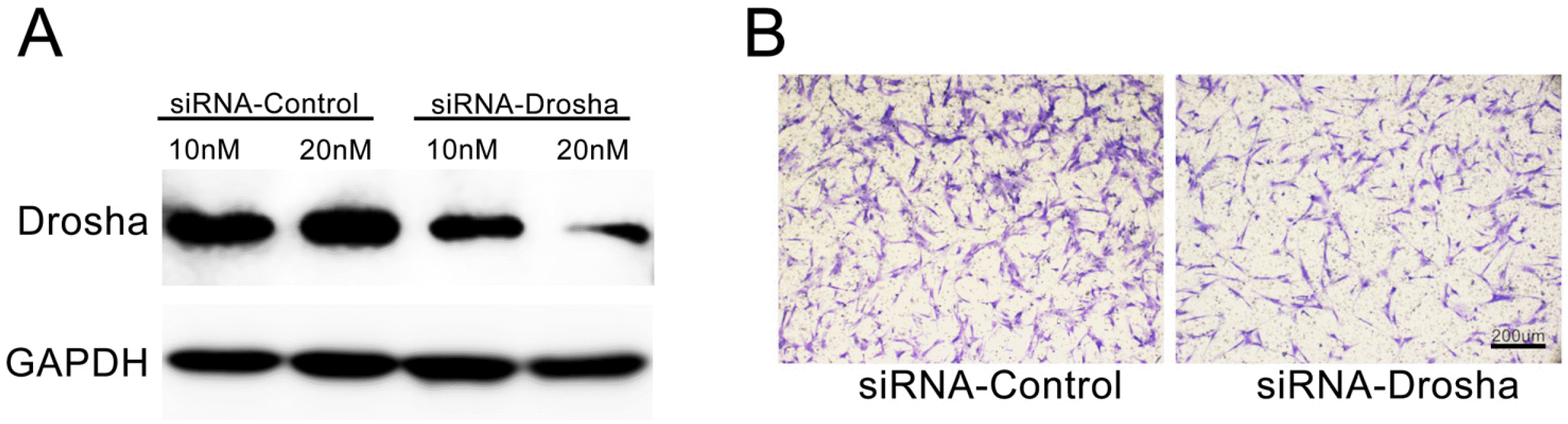
Fig. 5
(A) After siRNA-Drosha transfection for 48 hr, the knockdown efficiency of Drosha protein in PDLCs was evaluated by western blot analysis. Exosomes were collected from CTS-induced PDLCs after siRNA-mediated knockdown of Drosha and then cultured with BMSCs in Transwell plates. (B) Representative images of migrated BMSCs treated with Exos_CTSsiRNA-Drosha and Exos_CTSsiRNA-control.
![]()
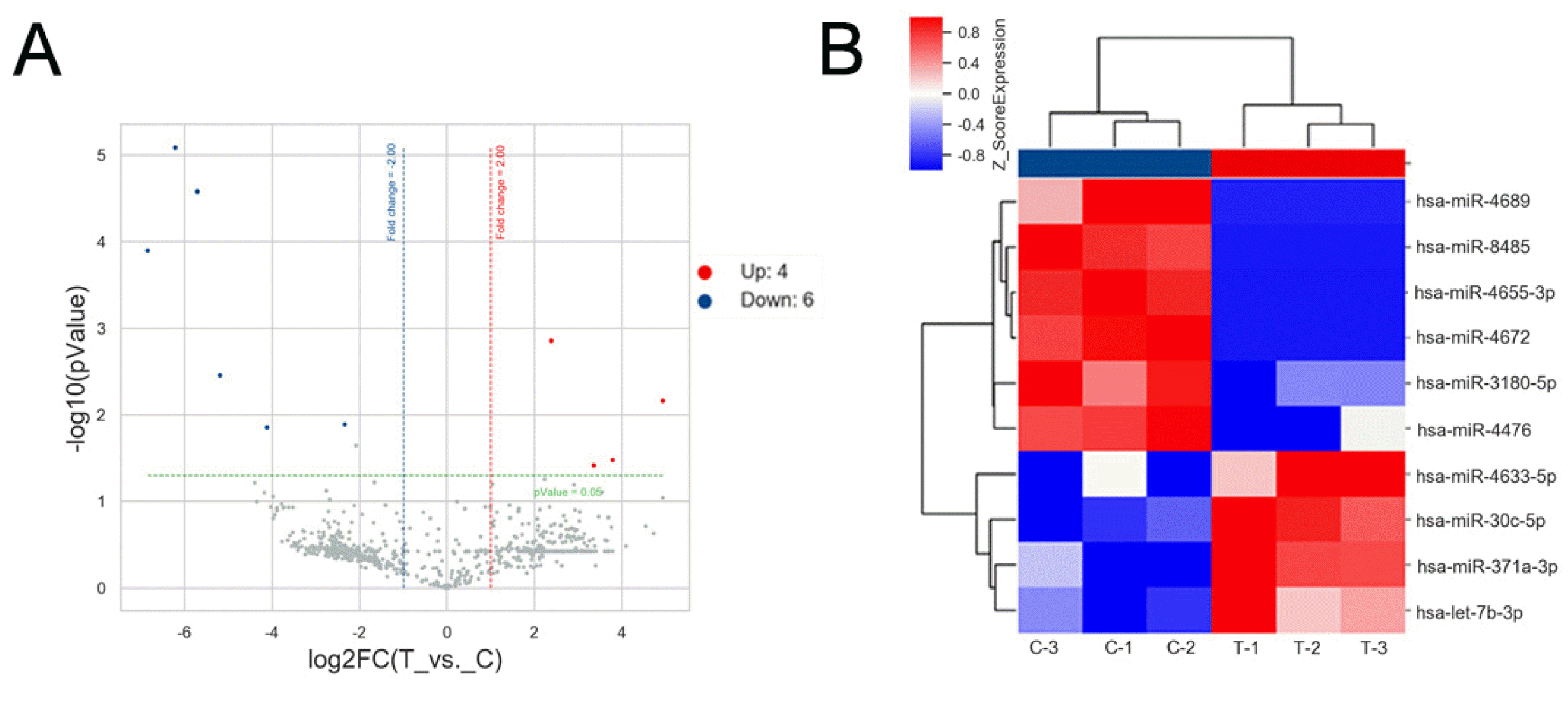
miRNA profiles of CTS-PDLC-derived exosomes were altered
A total of 769 known mature miRNAs were found in PDLC-derived exosomes through miRNA microarray. The Volcano plot shows a fold-change >2, p-value<0.05, and the distribution of differentially expressed miRNAs (Fig. 6A). The heatmap shows the clusters (Fig. 6B). Four miRNAs (miR-4633-5p, miR-30c-5p, miR-371a-3p and let-7b-3p) were upregulated and six miRNAs (miR-4689, miR-8485, miR-4655-3p, miR-4672, miR-3180-5p and miR-4476) were downregulated in exosomes derived from CTS-PDLCs compared to those derived from unstretched PDLCs.
Predicted target genes & hub genes in the protein-protein interaction network
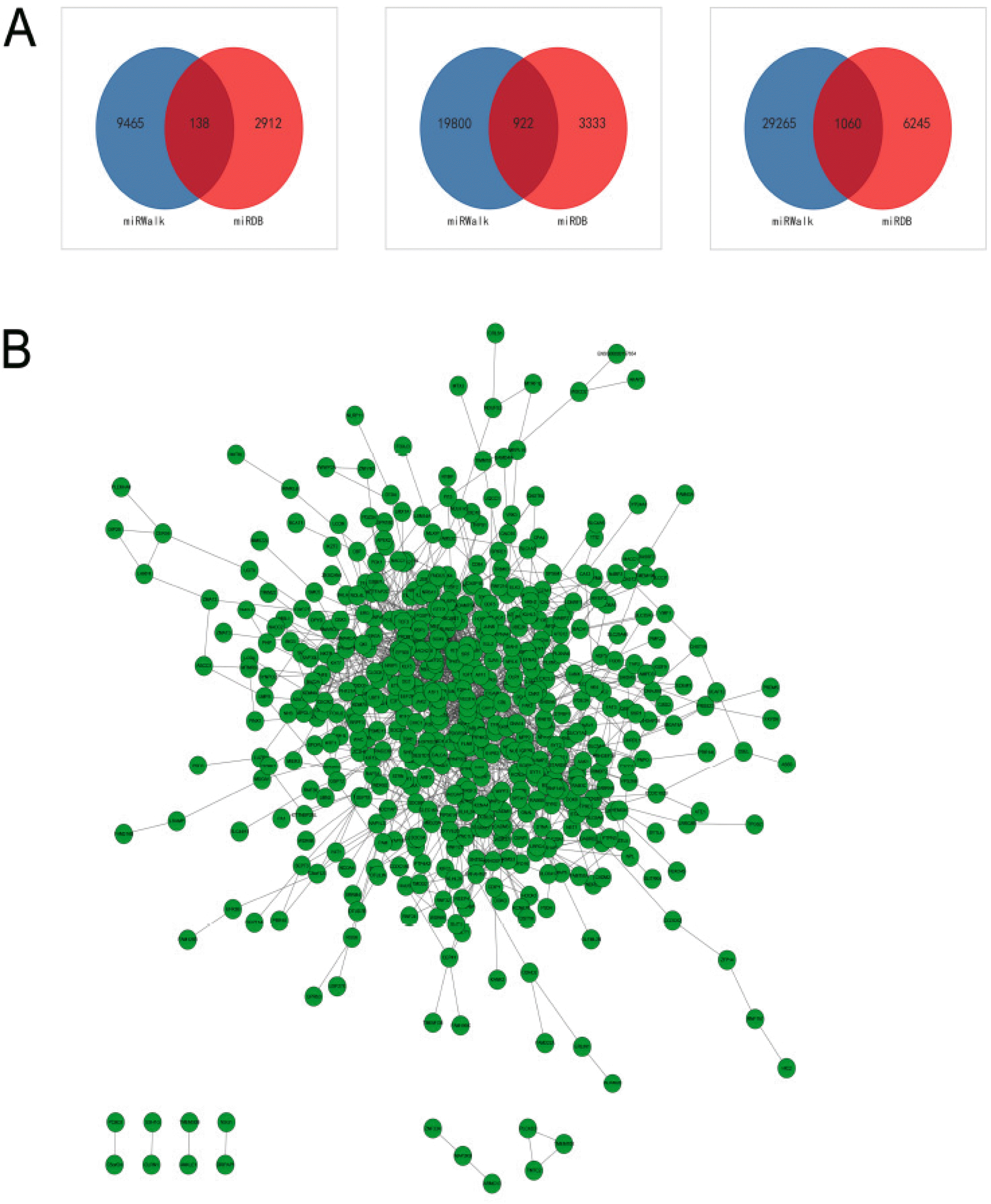
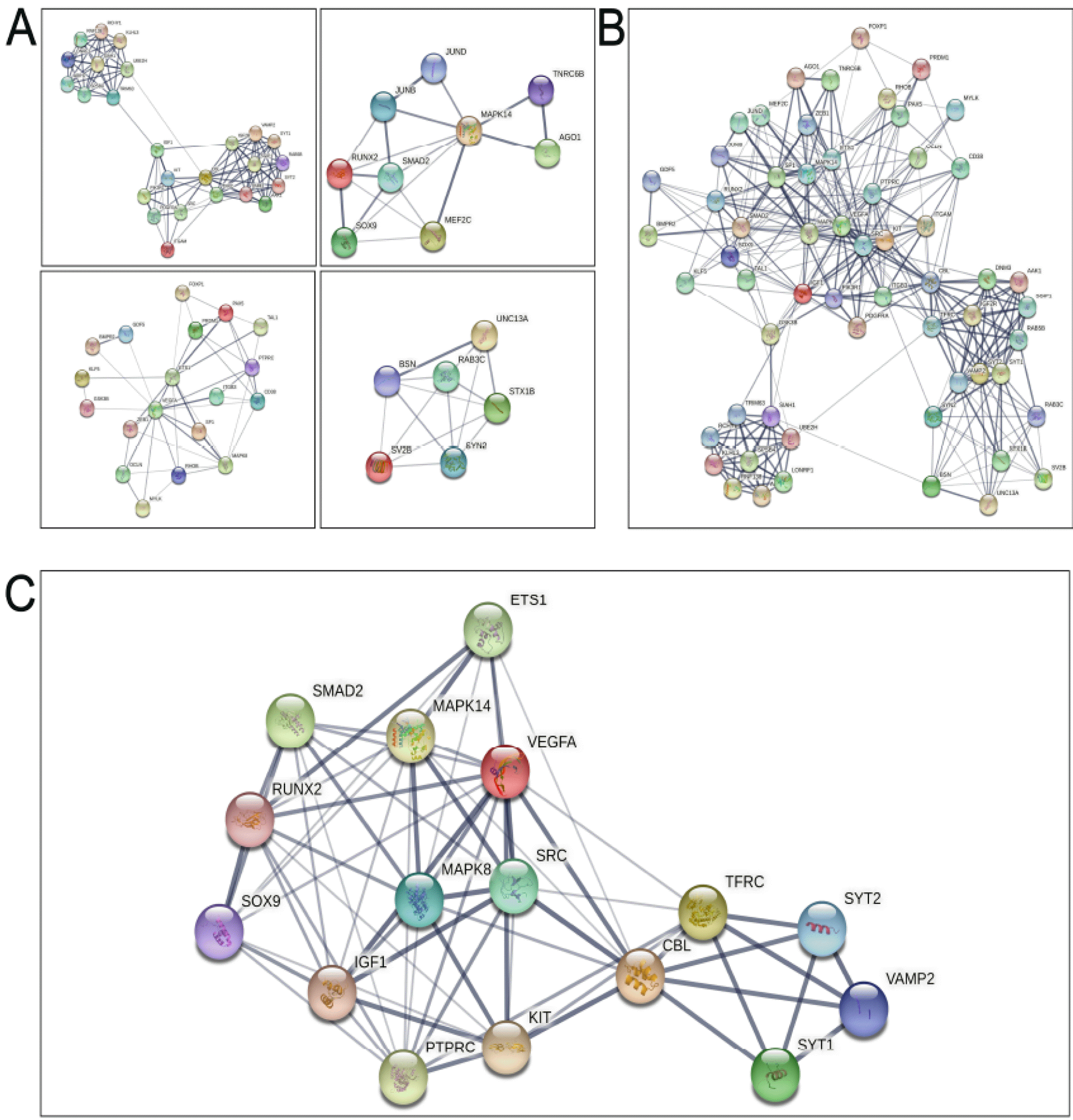
The 10 miRNAs were input into miRWalk3.0 and miRDB. In total, 922 upregulated and 138 downregulated genes (1,060 total) were found (Fig. 7A). Interactions between the 1,060 genes were analysed in STRING and visualized in Cytoscape. 526 genes were found 1,699 interactions to each other in total. These genes and interactions were visualized as nodes and lines, which formed a large interaction network (Fig. 7B). MCODE was used to detect the highest degree of clustering and found the top four significant modules, which comprising a total of 59 proteins (Fig. 8A). The 59 proteins also have close interactions as shown in Fig. 8B. The interactions were further counted and ranked by STRING. Finally, the top 20% of proteins (16 proteins) were selected and considered the hub proteins. These hub proteins and their interactions were visualized in STRING (Fig. 8C).
GO function and KEGG pathway enrichment analysis of hub proteins
To understand the functions of the sixteen-protein network, the CluoGO network analysis plug-in was used for GO and KEGG annotation. KEGG and GO functional analysis revealed that these proteins were enriched in the PI3K signalling pathway and some others such as endocytosis and exocytosis (Fig. 9). There are five of the hub genes (CBL, IGF1, KIT, SOX9 and SRC) were found related to the positive regulation of PI3K signalling pathway.
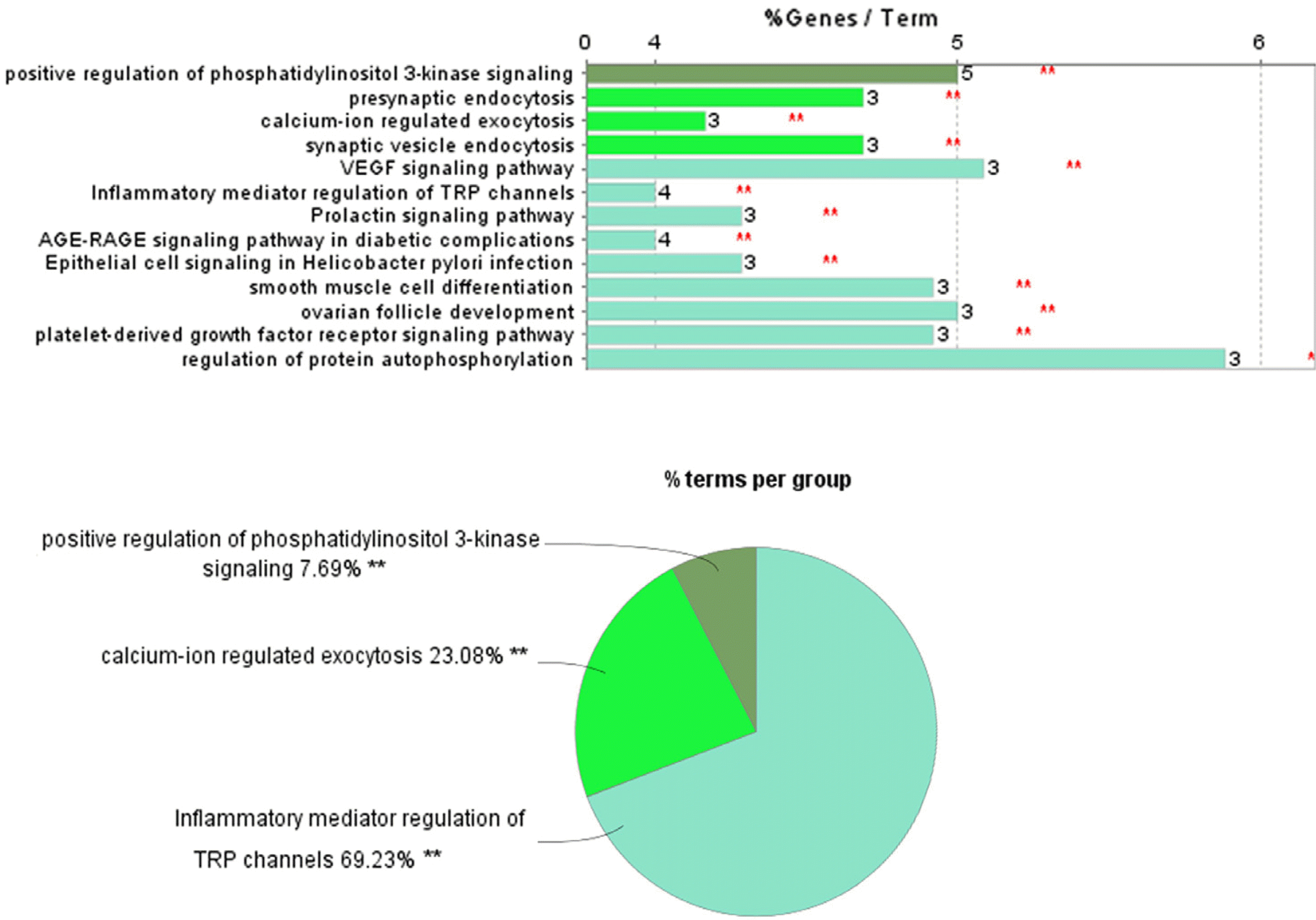
Discussion
In this study, we isolated and identified PDLC-derived exosomes. We found that exosomes but not MVs of cyclic tension stretch-applied PDLCs increased the migration ability of BMSCs. We also discovered the alternation of miRNA profiles in CTS-PDLCs and unstretched PDLC-derived exosomes. With bioinformatics analysis, we identified hub genes of the differentially expressed exosomal miRNAs and predicted their functions in migration and vesicle transmission. We suggested key signal pathways that the CTS-PDLC-derived exosomal miRNAs may target when affecting BMSCs.
PDLC-derived exosomes were characterized by obser-vation of the typical morphology, particle size and the marker proteins. The western blot results showed that PDLC-derived exosomes expressed exosomal surface markers (CD63, CD9, CD81, Alix and TSG101) while vimentin and the nonexosomal marker (Calnexin) were not expressed. There seems no signal in the MVs lane. In a recent study, CD63 was not found in the cardiac cells derived large EVs, similar to our results. They found Alix and calnexin much less expressed in large EVs than cells and exosomes. It may due to the exosome presence in their large EVs which have a range particles size from 50 nm to 300 nm (30). In our study, MVs showed negative expression of exosomes’ markers CD63, CD9, CD81, Alix and TSG101, which helped to demonstrate the accuracy of exosome isolation and higher purification. As to there were no western blot signal of calnexin and vimentin in MVs, it may due to the positive signal of PDLCs and exosomes were very strong, the weak or no signal in MVs was hardly captured by a computer-controlled imaging instrument.
Extracellular materials from PDLCs can accelerate the migration of BMSCs. Secretory proteins, microvesicles and exosomes are relevant to bone marrow mesenchymal stem cells behaviour (31). In the present study, we found MVs depletion from the medium did not alleviate BMSC mig-ration. However, further removal of the exosomes weakened the effect. Thus, BMSC migration is affected by exosomes but not MVs derived from CTS-applied PDLCs.
Exosomes are small vesicles released by cells whose surface antigens are characterized by cell origin. These vesicles may play a key role in cellular communication by transmitting RNA, proteins and bioactive lipids (32). There are hundreds of articles about BMSC-derived exosomes but few research reports concerning the exosomes from other types cells treatment of BMSCs. In the present study, despite membrane proteins’ expression differences, both Exos_CTS and Exos_US can enter BMSCs smoothly in a short time. Transwell results showed that exosomes derived from stretched PDLCs enhanced the migration activity of BMSCs compared to the exosomes derived from unstretched PDLCs.
The contents of exosomes, such as nucleic acids (DNA, mRNA, miRNA, lncRNA, etc.), are of importance for the biological effects mediated by exosomes (22). miRNAs are present in extracellular fluids such as serum, plasma, saliva and urine. Moreover, the generation and transport mode of RNA are special. Unlike proteins, which can be directly secreted, most RNAs are transported by various vesicles. miRNAs are highly enriched in extracellular vesicles (21). In this study, we found the miRNA content in exosomes from PDLCs accounted for 60% of small RNA content (RNA quality assessment results as appendix materials). This indicates the important role of miRNA in exosomes. Our results also demonstrated that when siRNA-Drosha (20 nM) was used to deplete Drosha (a key protein in miRNA production) from CTS-PDLCs, the enhancement of CTS-PDLC-exosomes to BMSC migration was attenu-ated. Thus, miRNAs within the Exos_CTS may be responsible for the observed enhancement of BMSC migration. Our previous work had detected the difference of miRNA in stretched and unstretched PDLCs which regulate the process of OTM. miRNAs in exosomes may also vary in different conditions. Thus, we tested miRNA profiles by microarray and found the differential expression of 10 exosomal miRNAs (four upregulated and six downregu-lated) derived from CTS-applied PDLCs. The 10 miRNAs in PDLC-derived exosomes may regulate the BMSC migration after internalization. Among them, miR-30-5p and miR-3180-5p were found in exosomes from human Plasma samples (33, 34). miR-8485 was found in chondrocyte-de-rived exosomes and regulated the differentiation of BMSCs (35). miR-8485 was also reported in oral mucosal stem cell derived exosomes (36). The seven other miRNAs are rarely reported as exosomal miRNAs. There are some evidences that miR-30c-5p regulates the migration of tumour cells (37, 38).
The differentially expressed miRNAs may play a network-like regulation role on targets by acting on many mRNAs. Bioinformatics analysis can be used as an accurate and efficient way to analysis the high-throughput experiment produced data, shall benefit the identifying of key biological mechanisms and understanding of life process at the molecular level. Thus, we studied these 10 miRNAs in the miRNA-mRNA regulation network throu-gh a series of bioinformatics analyses. We found 16 hub proteins in the miRNA-mRNA regulation network, which probably are the potential exosomal miRNAs’ targets in BMSCs and affect BMSC behaviour.
Further GO and KEGG analysis showed that these potential hub genes were closely related to the regulation of vesicle transmission such as endocytosis and exocytosis. This revealed that the 10 miRNAs may be actively selected for loading into exosomes, at least partly, for the acceleration of exosome transfer.
Exosomes could enhance the proliferation, migration, and osteogenic differentiation of BMSCs by activating PI3K/Akt signalling (39). PTEN is a negative regulator of the PI3K/Akt signalling pathway. Inhibition of PTEN can promote the expression of CXCR4, thus enhancing the migration ability of cells by inducing the activation of the PI3K/Akt/mTOR signalling pathway (40). In this study, hub genes were found most related to regulation of the PI3K signalling pathway. Among the five genes related to PI3K pathway, CBL, SOX9 and SRC are predicted targets of miR-8485. IGF1 and KIT are predicted targets of miR-4633-5p and 4672. Besides, eleven of the sixteen hub genes (CBL, ETS1, MAPK14, PTPRC, SMAD2, SOX9, SRC, SYT1, SYT2, VAMP2 and VEGFA) are predicted targets of miR-8485. miR-8485 may be one of the keys to this biological regulation mechanism. Thus, further investigation of role of the PI3K pathway and exosomal miRNAs such as miR-8485 in BMSC migration is necessary. The lack of this investigation is a limitation of the present study.
 XML Download
XML Download