

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Ischemic heart disease, including myocardial infarction (MI), is a significant contributor to morbidity and mortality worldwide.1) An MI occurs when blood flow to the heart is blocked, resulting in a weakened heart muscle and impaired cardiac function, which increases the risk of heart failure (HF), arrhythmias, or cardiac arrest.2)3) Despite the development of revascularization techniques that have dramatically improved post-MI outcomes and reduced in-hospital mortality,4) they do not address the underlying damage caused to cardiomyocytes (CMs) that may be irreversible. Consequently, patients who develop these complications after MI may still have a poor prognosis, highlighting the need for more efficient and effective therapies that can stimulate the regeneration of damaged heart tissue and enhance cardiac function.
Numerous innovative therapeutics are under investigation to regenerate damaged myocardial cells and improve cardiac function. One promising approach is stem cell therapy, which uses stem cells to regenerate damaged heart tissue.5) However, before it can become a standard treatment for ischemic heart disease, several challenges must be addressed. One challenge is the need to ensure efficacy of stem cell therapy in improving cardiac function and reducing mortality rates. Clinical trials have yielded mixed results, with some demonstrating improvements in cardiac function and others showing no significant differences from placebo or standard treatments.6) Another area of research involves the use of growth factors, such as neuregulin-1 (NRG1) and synthetic peptide PEGylated insulin-like growth factor-1 (IGF-1), to promote cardiac regeneration by stimulating the proliferation and differentiation of CMs and enhancing angiogenesis.7)8) Gene therapy, which involves the delivery of therapeutic genes to cells to promote tissue repair, is also a potential approach for cardiac regeneration. Preclinical studies using genes encoding vascular endothelial growth factor (VEGF) and calcium-handling protein sarcoplasmic/endoplasmic reticulum Ca2+ ATPase 2a) have shown promise in stimulating the growth of new blood vessels and cardiac muscle tissue.9)10) However, clinical trials for cardiac regeneration have yielded only modest efficacy results, despite of having shown their safety.
To make cardiac regeneration a practical therapy, various technical and safety challenges still need to be addressed. These challenges include the development of efficient and safe delivery systems, the possibility of off-target effects or immune reactions, and the requirement for careful monitoring of long-term safety and efficacy. Extensive research has been conducted on these issues and have been reviewed elsewhere.11)12) Therefore, in this review, we focus on the hostile microenvironments present in infarcted hearts that can hinder the process of cardiac regeneration by impeding the survival, proliferation, and differentiation of both endogenous and exogenous myocardial cells. We introduce innovative therapeutic strategies, including (1) reconstruction of cardiac niche microenvironment, (2) recruitment of functional CMs by promoting their proliferation or differentiation, and (3) organizing 3-dimensional (3D) tissue construct beyond the CMs, that can overcome these hostile environments. Additionally, we examine the recent discoveries regarding the mechanisms that underlie these strategies to enhance heart regeneration and highlight the current obstacles in translating these methods to clinical practice.
STRATEGY I: RECONSTRUCTION OF CARDIAC NICHE MICROENVIRONMENTS
Cardiac niches refer to specialized microenvironments present within the heart, which are essential for supporting the survival and function of cardiac cells.13) These niches play a critical role in the development, maintenance, and repair of the heart. The niche constitutes a dynamic entity in which the control of cell function depends on the complex interaction of intrinsic and extrinsic factors. With aging, there are several changes that can occur in the cardiac niche, including an increase in inflammation, oxidative stress, and fibrosis, which can lead to a decline in cardiac function. Alterations of niche function in various cardiac diseases such as HF, MI, and arrhythmia can include impaired angiogenesis, chronic inflammation, and the changes in the extracellular matrix (ECM). These changes can have significant effects on cardiac function and contribute to the progression of disease (Figure 1).
Figure 1
Angiogenesis and immune modulation.
CPC = cardiac progenitor cell; ECM = extracellular matrix; EPC = endothelial progenitor cell; FGF = fibroblast growth factor; G-CSF = granulocyte-colony stimulating factor; HGF = hepatocyte growth factor; HPC = hematopoietic progenitor cell; MSC = mesenchymal stem cell; MΦ = macrophage; VEFG-A = vascular endothelial growth factor-A.
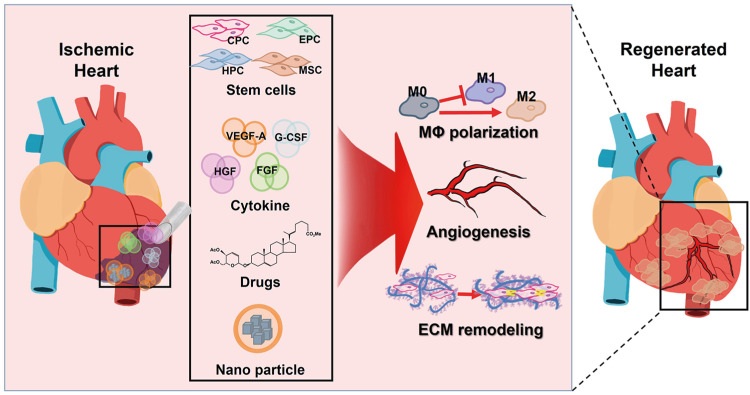
Angiogenesis
The coronary microvasculature is composed of arterioles, capillaries and venules, with specialized endothelial cells (ECs) that serve various functions such as acting as a selective barrier and regulating vasomotor, hemostatic, and inflammatory reponses.14)15) However, when the myocardium is exposed to prolonged ischemia, the essential functions of ECs can be lost, leading to reduced myocardial perfusion. The formation of new blood vessels, particularly from the ischemic border zone into the infarcted tissue, is vital in restoring oxygen and nutrient supply to the damaged tissue, and is considered a key factor in adequate post-ischemic repair.16) This process, known as angiogenesis, involves the sprouting new blood vessels from pre-existing vessels characterized by EC proliferation and migration, as well as the incorporation of circulating mononuclear cells.17) Angiogenesis plays a crucial role in the development and maintenance of cardiac niches by delivering nutrients and oxygen to the cells.
One approach to promote angiogenesis in the heart is by using growth factors such as VEGF-A, fibroblast growth factor (FGF), and hepatocyte growth factor (HGF), which stimulate the growth and proliferation of ECs and facilitate the formation of new blood vessels.18) However, endogenous growth factors may not be sufficient on their own to fully repair the damage, making it necessary to provide exogenous growth factors to boost cardiac repair and promote angiogenesis. Previous studies have shown that these growth factors can enhance the angiogenic response and improve cardiac function in various animal models of myocardial ischemia and reperfusion (I/R) injury. Saeed et al.19) utilized an adeno-associated viral vector containing the VEGF gene to treat a porcine model of myocardial I/R injury. They injected the vector into the myocardium 1 hour after reperfusion, resulting in increased infarct resorption and perfusion, better preservation of left ventricular function, and increased capillary and arterial density in the infarct and peri-infarct regions 8 weeks after treatment, as confirmed by contrast-enhanced cardiovascular magnetic resonance and histopathologic studies. Battler et al.20) observed an increased angiogenic response with higher microvessel counts in a porcine model of myocardial no-reflow phenomenon treated with FGF-2, while HGF gene transfer increased capillary density and preserved left ventricular function in a rabbit model of myocardial I/R injury.21) Similarly, Saeed et al.22) injected an adeno-associated viral vector containing HGF (VM202) into the myocardium of pigs after reperfusion, resulting in increased regional perfusion, and increased vascular densities after 50 days.
Stem cells, a type of undifferentiated cells with the ability to transform into various cell types, can also aid in promoting angiogenesis. Within bone marrow mononuclear cells (BMMCs), subsets such as endothelial progenitor cells (EPCs), hematopoietic progenitor cells (HPCs), and mesenchymal stem cells (MSCs), are believed to play a role in repairing the microvasculature through paracrine effects or by differentiating into ECs at sites of neovascularization.23) In patients with MI, HPCs and EPCs experience a peak in numbers during the early stages and the decrease after 7 days,24) indicating that their spontaneous migration from the bone marrow to the infarcted region may be a vital mechanism for myocardial repair. In a mouse model of myocardial I/R injury, Templin et al.25) injected HPCs into the artery root 24 hours after reperfusion and observed an increased capillary density in the treatment group after 5 weeks, despite the HPC myocardial engraftment being short-lived. Augmented angiogenic response was also observed in a rat model as well as a porcine model of I/R injury following MSC infusion in multiple studies.26)27)28) In an effort to promote angiogenesis after I/R injury, several studies have examined the potential of cytokine-induced mobilization of BMMCs. Granulocyte-colony stimulating factor (G-CSF), a substance that plays a significant role in mobilizing BMMCs from the bone marrow into the blood, were used in combination with stem cell factor by Kanellakis et al.29) in a murine I/R injury model and were found to enhance the number of blood vessels and improve left ventricular function. Additionally, in a porcine I/R injury model, Sato et al.30) showed an accelerated angiogenic response in treated animals through subcutaneous injection of G-CSF, as indicated by immunohistochemical analysis.
Based on this preclinical evidence, several clinical trials have been conducted to induce angiogenesis in patients with chronic coronary artery disease. While the efficacy of growth factor therapy for improving left ventricular function after MI is still uncertain and may only have a modest effect, modified MSCs that produce growth factors have shown potential for promoting angiogenesis and cardiac repair in infarcted heart tissue. Guo et al.31) showed that the preservation of cardiac function was improved with intramyocardial injection of MSCs that were transfected with the HGF gene, compared to the use of MSCs alone. Lu et al.32) conducted a study using a porcine model of I/R injury to compare the effectiveness of MSCs alone or genetically modified with VEGF or HGF for promoting myocardial repair and restoring cardiac function. The researchers found that implanting MSCs genetically modified with either VEGF or HGF genes into the infarcted myocardium through the retrograde coronary vein route prevented ventricular dysfunction after an MI.32) Both the MSC+HGF and the MSC+VEGF groups exhibited higher blood vessel densities in the infarct and peri-infarct regions than the MSC group, but there was no significant difference in vessel densities between the MSC+HGF and MSC+VEGF groups.32) The authors developed 2 novel strategies for enhancing vascular regeneration in infarcted hearts. The first, termed in vivo priming, involves epicardial implantation of a 3D cardiac patch containing genetically engineered MSCs that express HGF directly into the infarcted area of the heart.33) In this study, primed MSCs were shown to have improved vasculogenic potential and cell viability when exposed to HGF. This enhancement ultimately led to increased vascular regeneration and restored cardiac function in infarcted hearts. Further histological analyses confirmed that the primed MSCs survived longer within the cardiac patch and provided cardioprotection, as evidenced by a higher number of viable CMs in infarcted hearts.33) The second strategy involved using 2 different types of stem cells, ECs derived from induced pluripotent stem cells (iPSC-ECs) and MSCs that continuously secrete stromal derived factor-1α (SDF-eMSCs), to simultaneously promote post-natal vasculogenesis and angiogenesis, 2 core mechanisms in neovascularization.34) In order to promote more extensive vascular regeneration, we utilized iPSC-ECs to induce new vessel formation through vasculogenesis via intramyocardial injection. A 3D cardiac patch containing SDF-eMSCs (SDF-eMSC-PA) was also epicardially implanted to promote angiogenesis through sustained release of paracrine factors such as SDF-1α. The study demonstrated that iPSC-ECs contributed to the formation of new vessels in ischemic hearts, leading to improved cardiac function. Furthermore, co-implantation of SDF1α-eMSC-PAs increased the survival, retention, and vasculogenic potential of iPSC-ECs, resulting in more comprehensive neovascularization in hearts affected by MI.34) It is important to mention that the vessels formed through the dual stem cell approach were significantly bigger and more functional than the ones formed by iPSC-ECs alone.34) The results of these studies suggest that our strategy could be effective for treating ischemic heart disease.
Modulation of inflammatory and immune response
In the early stages following myocardial damage, inflammation plays a vital role by clearing debris and promoting angiogenesis.35) This process involves the recruitment of inflammatory cells, such as macrophages, neutrophils, and lymphocytes, to the site of injury, where they release cytokines and other inflammatory mediators that initiate the inflammatory response.36)37) However, excessive inflammation can be detrimental and impede the regenerative process, and maintaining the balance between pro-inflammatory and anti-inflammatory signals is critical for maintaining cardiac niche’s health. Therefore, regulating the inflammatory and immune response is essential for effective cardiac regeneration.
Ischemic injury mobilizes a diverse repertoire of innate and adaptive immune cells. Following ischemia, damage-associated molecular patterns released by dying cells,38) cytosolic DNA sensing pathway activation,39) and NOD-like receptor pyrin domain-containing protein 3 (NLRP3) inflammasome activation,40) are responsible for initiating the pro-inflammatory environment. Many immune cells interact with each other and release cytokines and chemokines to polarize existing macrophages and recruit additional monocytes. Infiltrating CCR2+ monocytes mature into proinflammatory Ly6chigh macrophages (M1), producing a potent cocktail of cytokines including interleukin (IL)-1β, IL-6, and tumor necrosis factor α (TNFα), while M2 macrophages promote heart repair and improve function following injury.41)42) The timing and ratio between M1 and M2 phenotypes of macrophages is critical, and tipping the balance toward M2-like macrophages is sufficient to stimulate heart repair and improve function following injury in animal models.42)43) In a prior study, the authors developed nanovesicles (NVs) derived from iron oxide nanoparticles-incorporated MSCs (IONP-MSCs) that mimic exosomes, termed IONP-NVs.44) These NVs showed improved retention within infarcted heart, which was augmented by magnetic guidance. Injection of IONP-NVs with magnetic guidance induced an early shift from an inflammation phase to a reparative phase in the infarcted myocardium, resulting in reduced apoptosis and fibrosis, and enhanced angiogenesis and cardiac function recovery.44) This dual approach may improve the therapeutic efficacy of MSC-derived NV therapy.
It has also been observed that the inflammatory environment of the injured heart attracts significant infiltrates of neutrophils alongside monocyte recruitment. Neutrophils participate in various aspects of the myocardial injury response, including amplifying pro-inflammatory signals, secreting proteolytic enzymes to remodel the ECM, and producing reactive oxygen species. In a previous study, the authors showed that the use of CU06-1004, a vascular leakage blocker, improved cardiac function by inhibiting EC hyperpermeability and subsequently reducing the plugging and infiltration of neutrophil in infarcted hearts.45) The polarization of macrophages towards a reparative M2 phenotype by efferocytosis may be an another way in which neutrophils promote cardiac recovery.46) Targeting subsets of pro-inflammatory neutrophils may, therefore, offer therapeutic opportunities.
The adaptive immune system is integral in harboring a pro-inflammatory response in the injured heart. Post-MI, cross-priming dendritic cells activate many classes of pro-inflammatory T cells, which infiltrate the injured myocardium in response to chemotactic signals, resulting in sustained myocardial injury and decreased left ventricular function.47)48) Pro-inflammatory CD8+ T cells produce potent cytokines, such as IL-17, interferon-γ, and TNFα.49) As shown in a mice model, adoptive transfer of splenic T cells from HF mice was sufficient to induce HF in naive recipient mice.50) However, some evidence suggests that injection of dendritic cells primed in injured hearts may help coordinate a beneficial, pro-reparative myocardium through the coordination of regulatory T cells (Treg) and shifting macrophage polarization to the M2-like phenotype.51) Furthermore, Foxp3+ Treg cells have been shown to stimulate recovery of injured heart muscle by driving pro-reparative phenotypes.52)53) Although CD4+ and CD8+ T cells’ rapid pro-inflammatory response may be beneficial in the acute phase of myocardial injury by enhancing scar formation, it may be maladaptive in the long term.
Therapies aimed at reducing inflammation and promoting tissue repair have been found to improve cardiac function after injury. Nonsteroidal anti-inflammatory drugs (NSAIDs) like aspirin54) and ibuprofen55) have demonstrated efficacy in reducing inflammation and improving cardiac function in preclinical models of MI, but prolonged use of NSAIDs can also have adverse effects such as gastrointestinal bleeding and renal impairment.56) Immunosuppressants such as cyclosporine and tacrolimus are also being used as a way to suppress the immune response in the cardiac niche. These drugs can impede the activation of fibroblasts and the excessive production of collagen, thereby preventing the development of myocardial fibrosis and scar formation in the injured heart. Targeting specific components of the inflammatory and immune response is another approach. For example, inhibiting the NLRP3 inflammasome has shown potential to improve cardiac function and reduce inflammation in preclinical models.57) Modulation of T cell responses, particularly by increasing anti-inflammatory Tregs, has also been investigated as a potential strategy for cardiac regeneration.58)
Extracellular matrix remodeling
The ECM is a complex network of proteins and carbohydrates that provides structural support to cells and tissues, including the heart. Recent evidence suggests that the cardiac ECM plays an active role in regulating the behavior of cardiac cells, orchestrating their proliferation, differentiation, and migration. For instance, hyaluronan and its binding protein Hapln1 are important for promoting CM expansion during heart regeneration in zebrafish, and their genetic ablation or inactivation leads to decreased CM proliferation and impaired regeneration.59) Agrin, a heparan sulfate proteoglycan abundant in regenerating hearts, has been shown to promotes CM proliferation in vitro and reactivate CM proliferation in adult mice and pigs after an MI.60)61) Similarly, periostin, an ECM protein expressed in the developing and adult post-MI mammal hearts,62) has been found to induce adult CMs to re-enter the cell cycle after MI via the PI3K/Akt pathway in rats and pigs.63)64)
As previously mentioned, angiogenesis is crucial for cardiac revascularization after MI and is mainly responsible for the formation of new blood vessels in adult organisms.65) The ECM plays a pivotal role in this process by supporting the growth of sprouting vessels and stabilizing their structure, as well as regulating EC function and providing a reservoir of cytokines and growth factors that promote angiogenesis.66)67) Several studies have shown that ECM proteins can enhance the revascularization of the infarcted area, leading to a reduction in CM apoptosis and fibrosis.68)69)70) This process is closely linked to CM migration to the injury site, particularly during neonatal regeneration in mammals, and is highly dependent on the ECM.71) Additionally, the matricellular factor cellular communication network factor 2a is essential for the attachment of CMs to newly formed blood vessels and subsequent myocardial regeneration.72)
The mechanical properties of the ECM, in addition to its composition, also have a significant impact on the proliferation and regeneration of cardiac cells. During the development period, there is a transition from high CM proliferative potential to low potential, which coincides increased ECM stiffness just after birth.73) Following myocardial injury like MI, the ECM undergoes extensive remodeling, resulting in biochemical and mechanical changes.74) This fibrotic response leads to increased stiffness of the ECM, affecting nearby cells. Previous studies suggest that a matrix with high stiffness can induce immature-like CM phenotypes,75) whereas CMs grown on softer surfaces may have better potential to de-differentiate and proliferate.76) Moreover, the rigidity of the substrate primarily affects CM cytokinesis rather that karyokinesis, and chemical-induced sarcomere disorganization may trigger the CM cell cycle re-entry.76)
The molecular and structural cues that regulate CM proliferation and angiogenesis suggest that ECM could be a novel therapy for inducing cardiac regeneration after MI. The whole ECM can be applied by sewing or gluing it onto the epicardial surface or injecting it into the infarcted region. Combining the ECM with human cardiac progenitor cells (CPCs) to increase its retention rate has also been explored, but subsequent studies showed that the therapeutic effect was mainly due to presence of the ECM itself, promoting tissue remodeling and preserving cardiac function in rat MI models.69)70) These positive results led to a recent clinical trial VentriGel in post-MI patients with left ventricular dysfunction.77) VentriGel is an ECM hydrogel derived from decellularized porcine myocardium and was directly injected into the endocardium of post-MI patients. Although the study was not designed to evaluate efficacy, there were indications of improvement based on increased 6-min walk test distance and decreased New York Heart Association functional class across the entire patient cohort.77)
STRATEGY II: RECRUITMENT OF FUNCTIONAL CARDIOMYOCYTES BY PROMOTING THEIR PROLIFERATION OR DIFFERENTIATION
The limited regenerative capacity of adult CMs highlights the importance of recruiting functional CMs for cardiac regeneration, as they make up the majority of the heart tissue and are responsible for its pumping action. Although CMs may divide and proliferate under certain circumstances, such as during development or after injury,78)79) this regenerative capacity decreases with age and may not be enough to fully restore cardiac function after myocardial injury. Currently, the approach after restoration of blood flow in MI patients involves close monitoring and limiting adverse remodeling, but not regeneration. However, there is an opportunity to activate endogenous CM recruitment by stimulating CM proliferation or delivering exogenous cells to replace lost CMs during this critical time window (Figure 2).
Figure 2
Recruitment of functional cardiomyocytes.
CM = cardiomyocyte; HAND2 = Heart And Neural Crest Derivatives Expressed 2; hESC = human embryonic stem cell; hiPSC = human induced pluripotent stem cell; hPSC-CM = human pluripotent stem cell-derived cardiomyocyte; NRG1 = neuregulin 1.
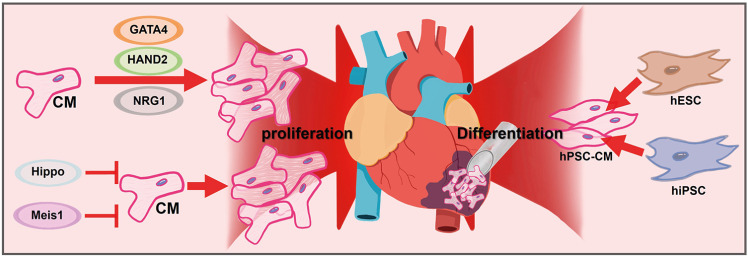
Endogenous cardiomyocyte recruitment by stimulating cardiomyocyte proliferation
It has been a long-held belief that mammalian heart tissue cannot regenerate and that cardiac cells are incapable of cell division.80) It has been discovered that the primary source of new cardiac cells after injury is pre-existing CMs,81) and several intrinsic signaling pathways can regulate their re-entry into the cell cycle and proliferation.79)80) Studies have also identified transcriptional regulatory networks that control cardiac muscle cell proliferation and regeneration, including GATA4,82) Hand2,83) and MEIS1,84) with the latter being able to induce cell-cycle re-entry in adult CMs when deleted.85)
The Hippo signaling pathway is a pathway that regulates tissue and organ size by controlling cellular proliferation, apoptosis, and cell fate decisions.86) When the Hippo pathway is inactive, YAP/TAZ proteins are dephosphorylated, translocate to the nucleus, and promote gene expression that promotes cell proliferation.87) Studies have shown that Hippo signaling inhibits adult cardiac regeneration.88) In rodents, deletion of Hippo signaling components has resulted in improved cardiac repair and reversal of HF after MI.89) In this study, direct myocardial delivery of AAV-Salvador-shRNA during, or even weeks after injury, improved heart function.89) Other studies have demonstrated that cardiac-specific YAP activation following MI promoted CM proliferation and reduced infarct size, while preserving cardiac function in a murine model.90) Recent reports have also revealed that the Hippo effector YAP interacts with the dystrophin-glycoprotein complex and the extracellular protein agrin at the plasma membrane to regulate CM proliferation.91) Recent profiling of human HF patients also suggest that inhibiting Hippo signaling may be a viable strategy to treat pump failure.89) Altogether, the inhibition of the Hippo pathway may lead to the mobilization of endogenous cardiac muscle cells, which could potentially be used as a direct treatment for HF in human patients.
There are several types of ligand/cell surface receptor complexes that can exogenously stimulate cell cycle re-entry of CMs and promote regeneration. One such complex is NRG1, a member of the epidermal growth factor family that plays a critical role in cardiac development and homeostasis.92) NRG1 is essential for ventricular wall trabeculation and expansion of the compact zone.93) Treatment of neonatal rat CMs with NRG1 in vitro results in increased F-actin organization, proliferation, protein production, and hypertrophy.94)95) The mitogenic effects of NRG1 on CMs in vitro have led investigations into its potential role in cardiac regeneration. In zebrafish, injured hearts express NRG1 protein in the outer layer of the heart wall,96) and exogenous NRG1 expression in uninjured zebrafish hearts promotes CM proliferation and dedifferentiation.97) In adult mice, NRG1-mediated activation of the ERBB2 receptor heterodimer stimulates CM proliferation,98) and transient induction of a constitutively active ERBB2 receptor is enough to reawaken CM proliferation in adults.99) Recently, Aharonov et al.100) showed that transient overexpression of activated ERBB2 in adult CMs promotes cardiac regeneration in a HF model. This restoration of ERBB2 signaling led to an epithelial-mesenchymal transition (EMT)-like response within resident CMs characterized by cytoskeletal remodeling, junction dissolution, and ECM turnover. YAP is involved in this process as it interacts with nuclear-envelope and cytoskeletal components, which reflects an altered mechanical state elicited by ERBB2.100) This study suggests that reactivation of ERBB2-mediated YAP signaling in adult CMs involves the developmental EMT-like behavior, resulting in robust heart regeneration.
Exogenous transplantation of cardiomyocytes derived from pluripotent stem cells
Human pluripotent stem cells (hPSCs) have the ability to differentiate into various cell types including CMs (hPSC-CMs), which offer a promising avenue for cardiac repair.101) In recent years, significant progress has been made in generating functional hPSC-CMs, both embryonic stem cells (hESCs) and induced pluripotent stem cells (hiPSCs). These cells can be generated in large numbers and can be used for cell replacement therapies, tissue engineering, drug screening, and disease modeling.101) Several studies have shown that transplantation of hPSC-CMs can improve cardiac function in MI animal models including mouse, rat and pig.102) In our previous study, we also demonstrated that the iPSC-CM aggregates exhibited higher survival and engraftment potentials compared to single CMs, resulting in improved cardiac function in a rat MI model.103) Furthermore, we also exploited the advantages of dual approach with the transplantation of hPSC-CMs and human MSC-loaded patch to amplify cardiac repair in a rat MI model.34) The epicardial implantation of hMSC-PA provided a complimentary microenvironment that improved vascular regeneration by secreting paracrine factors, and it significantly increased the retention and engraftment of intramyocardially injected hPSC-CMs, resulting in restoration of cardiac function.34) The majority of injected hPSC-CMs exhibited an adult CM-like morphology, suggesting that the secretomic milieu of hMSC-PA had pleiotropic effects in vivo.34) A study conducted on macaque monkeys was the first clinical-scale trial of hESC-CMs. In this study, the injection of hESC-CMs into the myocardium of infarcted hearts resulted in successful remuscularization after 2 weeks of I/R injury. The grafts showed excellent survival rates for over 3 months without any teratoma formation or off-target effects.104) Another study conducted by Liu et al.105) on macaque monkeys demonstrated that the transplantation cryopreserved hESC-CMs improved left ventricular ejection fraction after ischemic injury, with durable and robust improvement in cardiac function without arrhythmias.
However, there are still many challenges that need to be overcome before this approach can be used clinically. These include improving the efficiency and safety of CM generation, ensuring proper engraftment and integration of transplanted cells into the heart tissue, and avoiding immune rejection. Several clinical trials are currently underway to test the safety and efficacy of iPSC-CM therapy for cardiac regeneration in humans (NCT04696328, NCT04396899, NCT03763136. These trials are aimed at determining the optimal cell type, delivery method, and dosing regimen for the therapy. While there are still challenges to be overcome, iPSC-CM based regeneration therapy holds great promise for the treatment of ischemic heart disease by improving heart function.
STRATEGIES III: ORGANIZING 3-DIMENSIONAL TISSUE CONSTRUCT BEYOND THE CARDIOMYOCYTES
While CMs are the most studied cell type in this field, there is growing evidence that non-CMs play a crucial role in cardiac regeneration by providing structural and functional support to the injured heart. Indeed, co-culturing CMs with ECs and CFs in the 3D construct can mimic the complex microenvironment of native tissues and play important roles in regulating the function of CMs. In addition, 3D engineered tissue constructs using biomaterials can be used to create a supportive environment for multiple cardiac cells to grow and interact (Figure 3).
Figure 3
Exploring 3D structures beyond cardiomyocytes.
hESC = human embryonic stem cell; hiPSC = human induced pluripotent stem cell, hPSC-CM = human pluripotent stem cell-derived cardiomyocyte; hPSC-EC = human pluripotent stem cell-derived endothelial cell; hPSC-CF = human pluripotent stem cell-derived cardiac fibroblast; PLGA = polylactic-co-glycolic acid.
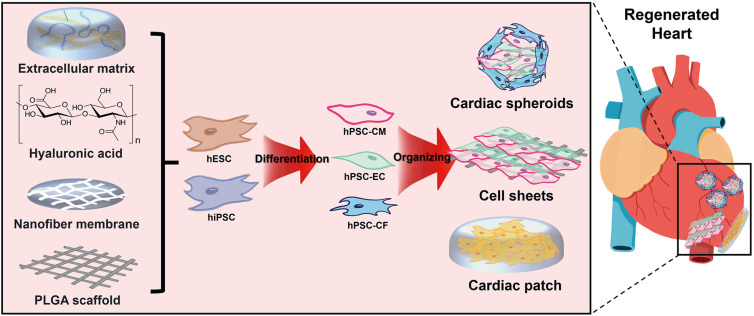
Cardiac organoids and spheroids
Organoids are 3D structures that can be generated from stem cells or tissue-derived cells in vitro, and they can recapitulate some of the structural and functional characteristics of the corresponding organs.106)107) In the context of cardiac regeneration, organoids have been proposed as a potential approach for generating functional heart tissue for transplantation. Cardiac organoids can be generated by differentiating hPSCs into various cardiac cell types including CMs, ECs and cardiac fibroblasts (CFs), and assembling them into 3D structures that resemble the architecture of the heart.108)109) Increasing interest has been directed towards non-CM cell types in driving myocardial renewal. CFs produce native ECM proteins and show bidirectional signaling crosstalk with CMs. Complex heterocellular coupling between CMs and CFs through gap junctions and intercellular calcium signaling influences contractile function and electrical conduction in the heart. In a recent study, Wu et al.110) investigated how post-natal CFs and ECM can influence cytokinesis in CMs. The researchers discovered that CF-mediated modulation of ECM can affect CM division and induce their binucleation, providing evidence for the role of CFs and ECM in regulating CM proliferation. The study also identified SLIT2 and Nephronectin as CF-derived ECM components that promote CM proliferation, highlighting potential targets for further investigation into the mechanisms underlying CM cytokinesis from embryonic to adult stages.110) However, these interactions can vary depending on factors such as heart development to pathology, leading to changes in cardiac structure and function. Li et al.111) explored the direct structural and functional effects of fetal and adult CFs on CMs using a tissue-engineered 3D co-culture system. They found that tissues with adult murine CFs displayed slower conduction velocity, higher stiffness, and decreased calcium transient amplitude compared to those with murine fetal CFs. Mastikhina et al.112) also showed that CFs played a role in pathologic fibrotic remodeling of the heart with the addition of transforming growth factor β as a trigger for fibrosis.
ECs also influence cardiac performance and remodeling through intercellular communication; small molecules and peptides secreted by ECs play a critical role in CM contractility. When co-cultured with CMs, ECs are also capable of forming capillary-like structures in engineered cardiac tissues (ECTs), which might enhance CM organization and support long-term survival in ECTs. Sekine et al.113) reported the formation of greater number of capillaries in CM sheets co-cultured with ECs compared to neonatal rat CM-only. In the presence of preformed EC vascular networks, CM sheets had better contractile properties with high expression of CM structural and gap junction proteins.113) After transplantation onto infarcted rat heart, capillaries from the CM sheets spread into host heart and connected with host capillaries, which was accompanied by recovery of cardiac function and reduced fibrosis in the surrounding host myocardium.113) In our previous study, we also demonstrated that in-vivo priming human MSC sheets by stacking HUVEC sheets could lead a significant improvement in the cardiac function, including intrinsic contractibility and remodeling.114)
ECTs from CM-EC and CM-CF co-cultures have better structural and functional properties with greater resemblance to native myocardium than those from pure CMs. Compared to ECTs with neither ECs nor CFs, CM-EC-CF tri-culture tissues have enhanced contractile function. According to Giacomelli et al.115) the tri-culture tissues displayed longer contraction duration and higher contraction amplitude compared to hiPSC-derived CM-EC and CM-CF co-cultures. In addition to electrophysiological maturation, tri-culture tissues also showed sarcomeric maturation. Lesman et al.116) found that transplanting CM-EC-CF seeding porous scaffolds onto naive rat hearts resulted in improved CM survival and larger graft formation. Furthermore, the vessels in tri-culture tissues could function post-transplantation and were perfused by host circulation. However, despite the evidence that ECTs co-cultured with CM-EC-CF provided increased contractile force, enhancing alignment and sarcomeric banding, and promoting structural and electrical maturation, their therapeutic potentials for cardiac regeneration have not yet been determined. One of the major limitations is the difficulty of generating large, vascularized organoids that can integrate with the host tissue and support long-term survival and function. Another challenge is the heterogeneity and variability of organoids, which can affect their reproducibility and reliability as models for drug screening or disease modeling. Overall, while organoids hold great promise for cardiac regeneration and disease modeling, further research is needed to optimize their generation, maturation, and integration with the host tissue, and to establish their safety and efficacy in preclinical and clinical settings.
Cell sheets and cardiac patches
Tissue engineering is another promising approach for cardiac regeneration. Tissue engineering involves using a combination of cells, biomaterials, and biochemical factors to create a functional tissue that can replace damaged or diseased tissue. One approach is to create a 3D scaffold that mimics the structure and function of the heart tissue, and then seed cardiac cells into the scaffold. The scaffold can be made from a variety of biomaterials, including synthetic polymers, natural ECM components, and decellularized heart tissues. The seeded cells can then grow and differentiate on the scaffold, forming a functional cardiac tissue. Ishigami et al.117) were able to show in a porcine model of MI that the infarcted myocardium could be functionally restored using large cardiac tissue sheets. The sheets consisted of various hPSC-derived cardiac cells, including CMs, ECs, and vascular mural cells.117) In our previous study, we also introduced a novel method for a comprehensive cardiac repair by sutureless transplantation of a highly integrable BM-MSC sheets stacked by HUVEC sheets based on the utilization of a highly aligned thermoresponsive nanofiber membrane.114) A prolonged secretion of multiple angiogenic cytokines, such as VEGF, angiopoietin-1, and IGF-1 seemed to lead a significant improvement in the cardiac function, including intrinsic contractibility and remodeling.114) Recently, Huang et al.118) developed an off-the-shelf therapeutic cardiac patch composed of a decellularized porcine myocardial ECM scaffold and biodegradable polylactic-co-glycolic acid microcarriers delivering human cardiac stromal cell-secreted factors. This fully acellular artificial cardiac patch maintained its potency after long-term cryopreservation, and provided cardioprotective, pro-angiogenic and proliferative potential in rodent and porcine models of MI.118) By combining a decellularized cardiac matrix with exosomes derived from MSCs, a new cell-free graft was developed with regenerative properties that were preserved even after cryopreservation.118) This promising approach offers a clinically compliant and ready-to-use strategy for therapeutic applications.
Another approach is to use bioprinting technology to create a 3D structure of cardiac tissue using a bioink that contains cardiac cells and biomaterials. Bioprinting allows for precise control over the spatial arrangement of the cells, which can improve its functional status. Gaetani et al.119)120) utilized extrusion-based 3D bioprinting to create an in vitro cardiac patch using RGD-modified alginate and hyaluronic acid/gelatin, which was loaded with human CPCs. Their work demonstrated the potential of bioprinting and showed improved cell viability, cardiac commitment, and in vivo engraftment over time, as well as the precise control over the microstructure of the patch. In a previous study, the authors manufactured ECM patches including human CPCs and MSCs by 3D bioprinting technology and demonstrated that the printed structure composed of spatial patterning of dual stem cells improved cell-to-cell interactions and differentiation capability and promoted functionality for cardiac regeneration. Kupfer et al.108) developed a 3D bioprinting technique using an ECM-derived bioink formulation that supported the viability, proliferation, and maturation of hPSCs into CMs. The resulting cardiac organoid had a complex geometry and robust electromechanical function, representing a significant advancement in hPSC differentiation.108) Ong et al.121) developed a biomaterial-free method to deliver stem cells using 3D bioprinted cardiac patches. They aggregated hPSC-derived cardiac cells with FBs and ECs to form mixed cell spheroids, which were then used to create cardiac patches that beat spontaneously and exhibited uniform electrical conduction throughout the patch. Immunohistochemistry revealed rudimentary blood vessel formation and the presence of cardiac gap junction protein, Cx43, localized to cell-cell borders. In vivo implantation into rat myocardium suggested vascularization and engraftment of the 3D bioprinted cardiac patche.121) While there are still challenges to be overcome, such as achieving optimal cell survival and integration into the host tissue, tissue engineering holds great promise for the treatment of heart disease by promoting cardiac regeneration and improving heart.
CONCLUSION
While stem cell therapy has shown potential for cardiac repair, there are several reasons why it is not yet a widely used treatment for ischemic heart disease, including limited effectiveness, safety concerns, technical challenges and lack of standardization. In this review article, we focused on the limited effectiveness of stem cell therapy and asked ourselves “Are there hopeful therapeutic strategies to regenerate infarcted hearts?” We already know that stem cells play a critical role in our bodies, and their growth and differentiation require a very delicate environment. However, we have not yet fully understood how to regulate it, especially in infarcted hearts. We acknowledge that to efficiently regenerate the heart, we require the activation of endogenous CPCs or the use of PSC-derived CMs with superior regenerative abilities. However, we should take their hostile microenvironment into account which may limit the effectiveness of therapies, and develop the innovative therapeutic strategies to overcome this hurdle, including (1) reconstruction of cardiac niche microenvironment, (2) recruitment of functional CMs by promoting their proliferation or differentiation, and (3) organizing 3D tissue construct beyond the CMs. The issue is not about which technology is more important or not, but each one will play an important role in exploring the effectiveness of regenerative therapy. Therefore, collaboration between researchers from diverse fields is necessary to make progress in this area.
 XML Download
XML Download