

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Gastric cancer (GC), a commonly diagnosed malignant cancer, accounted for 7.1% of all cancer-associated deaths in 2020 [1]. GC is associated with complex genetic and environmental interactions and is therefore considered to be a heterogeneous disease [2]. Despite the development of interventions such as gastrectomy and adjuvant chemotherapy, patients with GC have a poor prognosis because of the frequency of lymphatic or distant metastases [3]. Epigenetic changes are closely associated with GC invasion, metastasis, and chemoresistance [4,5]. Therefore, identifying the epigenetic regulatory mechanisms and risk factors involved in GC initiation and progression may provide additional therapeutic targets.
Long noncoding RNAs (lncRNAs) are associated with several epigenetic alterations and play epigenetic regulatory roles [6]. In addition, lncRNAs can influence post-transcriptional regulation by interacting with microRNA (miRNA)-mediated pathways, regulating multiple cellular processes [6]. High-throughput sequencing studies have identified a majority of lncRNAs with notable dysregulation in tumor samples or cells [7]. Alterations in lncRNA expression and mutations in lncRNAs can promote or restrain tumorigenesis and tumor metastasis, providing a new paradigm for the study of tumor biology [8]. LncRNAs are widely known to show cell-, tissue-, and developmental stage-specific expression patterns, and their dysregulation affects cancer cell growth, energy metabolism, chemoresistance, clinicopathology, and survival [9,10]. Accordingly, numerous lncRNAs with enormous regulatory potency in GC have been proposed as diagnostic or therapeutic biomarkers [11]. Currently, many of the lncRNAs identified to be differentially expressed in tumor samples in public databases, such as GEO and GEPIA, have not been functionally characterized. By analyzing data from GEPIA, we obtained the lncRNA LINC00562, whose expression is significantly upregulated in stomach adenocarcinoma, suggesting the cancer-promoting potential of LINC00562 in GC. However, the role of LINC00562 in GC has not been studied.
LncRNA/miRNA/mRNA networks have been widely used to explain tumor progression [12]. With the advances in computational bioinformatics [13,14], complementary sequences between lncRNAs and targeted miRNAs, as well as between miRNAs and targeted mRNAs, are easily obtained. These data provide a basis for the construction of lncRNA/miRNA/mRNA networks. LncBase and TargetScan were used in this study to predict that LINC00562, miR-4636, and adaptor protein complex 1 sigma 3 (AP1S3) may constitute a lncRNA/miRNA/mRNA network. MiR-4636 has been found to be downregulated in human colorectal cancer [15], GC [16] and cervical cancer [17], in which miR-4636 functions as a cancer suppressor. AP1S3 belongs to the adaptor protein (AP) family and is related to the classification of transmembrane proteins [18]. To date, there have been few reports on the role of AP1S3 in human cancers. Toda et al. [19] demonstrated that high AP1S3 expression level was significantly associated with poor prognosis in patients with breast cancer. In addition, AP1S3, overexpressed in glioma, promotes glioma progression [20]. However, the role of LINC00562–miR-4636–AP1S3 axis in glioma development remains unclear.
In the current study, we focused on the GC-associated lncRNA LINC00562 and explored its role in GC development. We also explored the regulatory mechanisms of GC by studying the targeted miRNAs and downstream functional genes. The miR-4636–AP1S3 network governed by LINC00562 was confirmed using rescue experiments. Our study is the first to characterize the role of LINC00562 in GC as a candidate cancer-associated lncRNA.
METHODS
Tissue specimens
Thirty-eight GC patients recruited at Wuchang Hospital Affiliated to Wuhan University of Science and Technology were enrolled in this study. The enrolled patients were diagnosed with GC for the first time, did not receive any therapies, fully realized the experimental procedures and provided written informed consent. Tumor specimens and matched normal specimens were excised during the surgical operation, frozen, and stored at –80°C. The present study was approved by the Wuchang Hospital Affiliated to Wuhan University of Science and Technology (approval number: 2021023).
Cell culture and transfections
Non-cancerous control cells (GES-1), normal gastric mucosa cell line (RGM-1), and GC cell lines [(HGC-27 (derived from metastatic site: lymph node), AGS (derived from gastric adenocarcinoma), and MKN45 (derived from undifferentiated carcinomas)] were purchased from Procell. GES-1, HGC-27, and MKN45 cells were cultured in RMPI1640 (Procell) supplemented with 10% fetal bovine serum (FBS, Procell), while AGS cells were cultured in Ham’s F-12k (Procell) supplemented with 10% FBS.
Small interfering RNA against LINC00562 (si-lnc) or AP1S3 (si-AP1S3) and their corresponding negative control (si-NC) were provided by Sangon Biotech. MiR-4636 mimic (miR-4636), miRNA mimic NC (miR-NC), miR-4636 inhibitor (inhibitor), and miRNA inhibitor NC (inhibitor-NC) were directly obtained from Ribobio. The experimental cells were subjected to various transfections using Lipofectamine 2000 (Invitrogen) and then incubated at 37°C with 5% CO2 for 24 h. Next, the cells were collected, and real-time quantitative PCR (RT-qPCR) or Western blotting was conducted to examine transfection efficiency. The sequences of the vectors are summarized in Supplementary Table 1.
RT-qPCR
For RNA isolation, small pieces of tissue or cell material were lysed using TRIzol reagent (Cwbio). The isolated RNA was identified using a NanoDrop spectrophotometer (Thermo Fisher Scientific). The assembly of cDNA was carried out using PrimeScript 1st strand cDNA Synthesis Kit (Takara) or the qScriptmicroRNA cDNA Synthesis Kit (Quantabio) for miRNA, followed by cDNA amplification for qPCR detection using SYBR Green Buffer (Takara) through a QuantStudio 6 PCR System (Applied Biosystems). The primers used for each factor are shown in Table 1. The relative expression level was calculated using the 2−△△Ct method [21], employing GAPDH or U6 for normalization.
Table 1
Real-time PCR primer synthesis list
![]()
Subcellular location
A commercial PARIS kit (Invitrogen) was used to extract RNA samples from the cytoplasm or nucleus of GC cells. Cytoplasmic or nuclear RNA was used for RT-qPCR to determine the abundance of LINC00562 in each part. GAPDH and U6 were used as internal references for the cytoplasm and nucleus, respectively.
Cell Counting Kit-8 (CCK-8) assay
Treated cells were seeded in 96-well plates (5 × 103 cells/well). Cells cultured for 24, 48, or 72 h were treated with the CCK-8 reagent (Cwbio; 10 μl per well). After 2 h of incubation, the absorbance at OD450 nm in each well was measured using a microplate reader (Thermo Fisher Scientific).
Western blotting
The primary antibodies used in this study included anti-Bax (Product# ab32503; Abcam; 1/2,000), anti-Bcl-2 (Product# ab32124; Abcam; 1/1,000), anti-AP1S3 (Product# PA5-23894; Invitrogen; 1/1,000), and anti-GAPDH (Product# ab9485; Abcam; 1/2,500), and goat anti-rabbit IgG (Product# ab205718; Abcam; 1/5,000) as the secondary antibody. Western blotting was performed as previously described [22]. Total proteins isolated using RIPA lysis buffer (Beyotime) were quantified using a BCA Kit (Beyotime). After separation by 10% SDS-PAGE, the separated blots were transferred to PVDF membranes and blocked with 5% nonfat milk for 2 h at room temperature. Membranes were then incubated with primary and secondary antibodies, and finally exposed to the ECL kit (Beyotime) to monitor protein density.
Colony formation
Cells transfected with si-lnc, miR-4636 inhibitor, si-AP1S3, si-NC, or inhibitor-NC were seeded onto 6-well plates (300 cells/well), recovered with 0.6% agar, and subsequently maintained at 37°C in incubators with 5% CO2 for 14 days. Cell transfection was performed every two days, and the conditions of colony growth were observed every two days until the colonies were visible. Cell colonies (> 50 cells) were fixed (methanol) and stained (0.1% crystal violet (Beyotime), counted and photographed.
Wound-healing assay
Treated cells were seeded into 24-well plates (5 × 104 cells/well) and cultured until they reached 90% confluence. A cell surface was created with a simulated wound using a sterile pipette tip. Images recording the distance of the wound at 0 h and 24 h were captured by light microscopy (Nikon), with a magnification of 40×.
Animal models
BALB/c nude mice (female; 6–8-week-old; six mice) were purchased from the Vital River. We injected HGC-27 cells with LINC00562 depletion or NC into nude mice to construct xenograft models. Briefly, short hairpin RNA (shRNA) against LINC00562 (sh-lnc) and sh-NC were synthesized and packaged using lentivirus by Sangon Biotech. HGC-27 cells were infected with sh-lnc or sh-NC and implanted into mice (2 × 106 cells/mouse) to induce tumorigenesis. All mice were housed regularly and tumor growth conditions were observed every day. The tumor volumes (length × width2/2) were measured weekly. Tumors were allowed to grow for five weeks, and then all mice were euthanized to excise tumor tissues. The animal protocol was authorized by the Wuchang Hospital Affiliated with Wuhan University of Science and Technology (approval number: WNH2021007).
Dual-luciferase assay
The public database LncBase was used to analyze the binding sites between LINC00562 and miR-4636. The public database TargetScan was used to analyze the binding sites between miR-4636 and AP1S3. A sequence fragment of LINC00562 (Forward: 5’- GCTAGCTTTTATCTTGGTAGC-3’ and Reverse: 5’-CTCGAGTTCGAGGTGCAATTG-3’) or AP1S3 (Forward: 5’-GCTAGCATTGTGGGAAGCAGC-3’ and Reverse: 5’-CTCGAGTTCCTCTATTAAAACA-3’) wild-type (WT) containing the miR-4636 binding site was synthesized and inserted into the pmirGLO vector (Promega). LINC00562 or AP1S3 mutant-type (MUT) mutated binding sites of miR-4636 were generated using the QuickMutation Site-Directed Mutagenesis Kit (Beyotime). Subsequently, WT and MUT reporter vectors of LINC00562 or AP1S3 were obtained. The WT or MUT constructs of LINC00562 or AP1S3 were transfected with the miR-4636 mimic (miR-NC as a control) into the experimental cells, and the cells were incubated for 48 h. Luciferase activity in the transfected cells was ascertained using the Dual-Luciferase Reporter Assay System (Promega).
RNA Binding Protein Immunoprecipitation (RIP) assay
Ago2 protein is a crucial element in miRNA-induced RNA‐induced silencing complex (RISC) an Ago2-mediated RIP assay was performed to explore the binding of miR-4636 to LINC00562 or AP1S3 using the RIP Assay Kit (Millipore). Briefly, magnetic beads were pre-treated with anti-Ago2 or anti-IgG (control). Lysates of GC cells were exposed to magnetic beads for RNA precipitation. RNA complexes on the beads were analyzed by RT-qPCR.
Statistical analysis
Three independent experiments were performed and the experimental data were processed using GraphPad Prism 8 (GraphPad). Data are expressed as means ± standard deviation. Student’s t-test was used for comparison between two groups, and one-way ANOVA followed by Tukey’s post-hoc test was used for comparison among multiple groups. The correlation between the two sets of data was analyzed using Pearson’s analysis. p-value < 0.05 was considered to be statistically significant difference.
RESULTS
LINC00562 expression levels were elevated in tumor samples and GC cells
First, we identified the expression pattern of LINC00562 in GC cells. High expression levels of LINC00562 were observed in not only tumor specimens but also in GC cell lines (HGC-27, AGS, and MKN45), in contrast to those in normal tissue specimens and GES-1 cells (Fig. 1A, B). In addition, LINC00562 expression level in normal gastric mucosa cells RGM-1 was not significantly different from that in GES-1 cells (Fig. 1B). HGC-27 and AGS cells with relatively high LINC00562 expression levels were used for further experiments. We determined that LINC00562 was expressed in more than 60% of the cytoplasm and less than 40% of the nucleus (Fig. 1C). In summary, we observed aberrant upregulation of LINC00562 in GC.
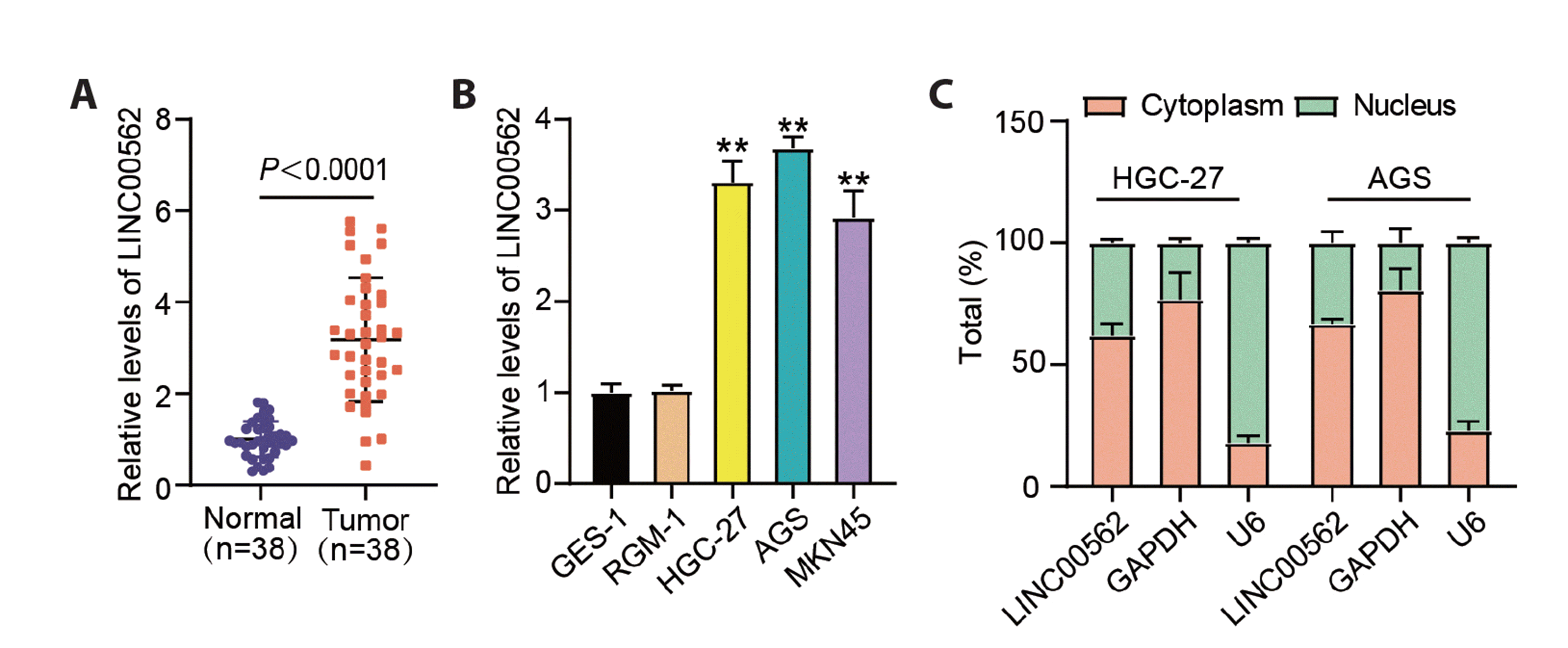
Fig. 1
LINC00562 expression was enhanced in GC.
(A) LINC00562 expression levels in tumor and normal specimens. (B) LINC00562 expression levels in GSE-1, RGM-1, HGC-27, AGS and MKN45 cells, **p < 0.01 in contrast to GES-1. (C) Distribution of LINC00562 in cytoplasm and nucleus of AGS and HGC-27 cells. Values are presented as mean ± SD. GC, gastric cancer.
![]()
Silencing LINC00562 restrained GC cell proliferation and migration but provoked apoptosis
The endogenous level of LINC00562 was knocked down in GC cells using si-lnc, to explore LINC00562 function. LINC00562 expression level was effectively reduced in HGC-27 and AGS cells after si-lnc transfection compared to si-NC (Fig. 2A). The biological function of GC cells was evaluated by LINC00562 downregulation. CCK-8 results showed that GC cells with LINC00562 expression depletion had OD450 values reduced by approximately 40% at 72 h post-incubation (Fig. 2B). In addition, Bax is an apoptotic protein, and Bcl-2 is an anti-apoptotic protein. Therefore, we used western blotting to detect the protein expression of Bax and Bcl-2. We observed that Bax expression level increased, whereas Bcl-2 expression was weakened in GC cells after LINC00562 downregulation (Fig. 2C). The colony-formation assay showed that the number of GC cell-induced colonies was remarkably decreased by over 85% following LINC00562 knockdown (Fig. 2D). In addition, through a wound-healing assay, we observed that the migratory capacity of GC cells was notably impaired after LINC00562 depletion (Fig. 2E). The in vitro data highlighted that LINC00562 depletion blocked the malignant behavior of GC cells.

Fig. 2
Silencing LINC00562 restrained GC cell growth and migration but enhanced apoptosis.
(A) LINC00562 expression in HGC-27 and AGS cells transfected with si-lnc or si-NC. (B) Effect of LINC00562 silencing on cell proliferation was assessed by CCK-8 assay in HGC-27 and AGS cells. (C) Protein levels of Bax and Bcl-2 were affected by LINC00562 silencing in HGC-27 and AGS cells. (D) Effect of LINC00562 silencing in colony formation by HGC-27 and AGS cells. (E) Impact of LINC00562 silencing on cell migration was assessed using wound-healing assay in HGC-27 and AGS cells. Values are presented as mean ± SD. ×200. GC, gastric cancer. **p < 0.01 in contrast to si-NC.
![]()
LINC00562 absence hindered GC tumor cell growth in vivo
After determining the anti-cancer effects of LINC00562 absence in vitro, we constructed animal models to confirm the effects of LINC00562 absence in vivo. Representative tumor images are displayed in Fig. 3A, showing a smaller tumor size in the sh-lnc group than that in the sh-NC group. Notably, a poorer tumor volume was observed in animal models with LINC00562 absence (Fig. 3B). As expected, tumor tissues isolated from sh-lnc-administered animals had lower tumor weights (Fig. 3C). The in vivo data confirmed that LINC00562 absence repressed tumor growth.
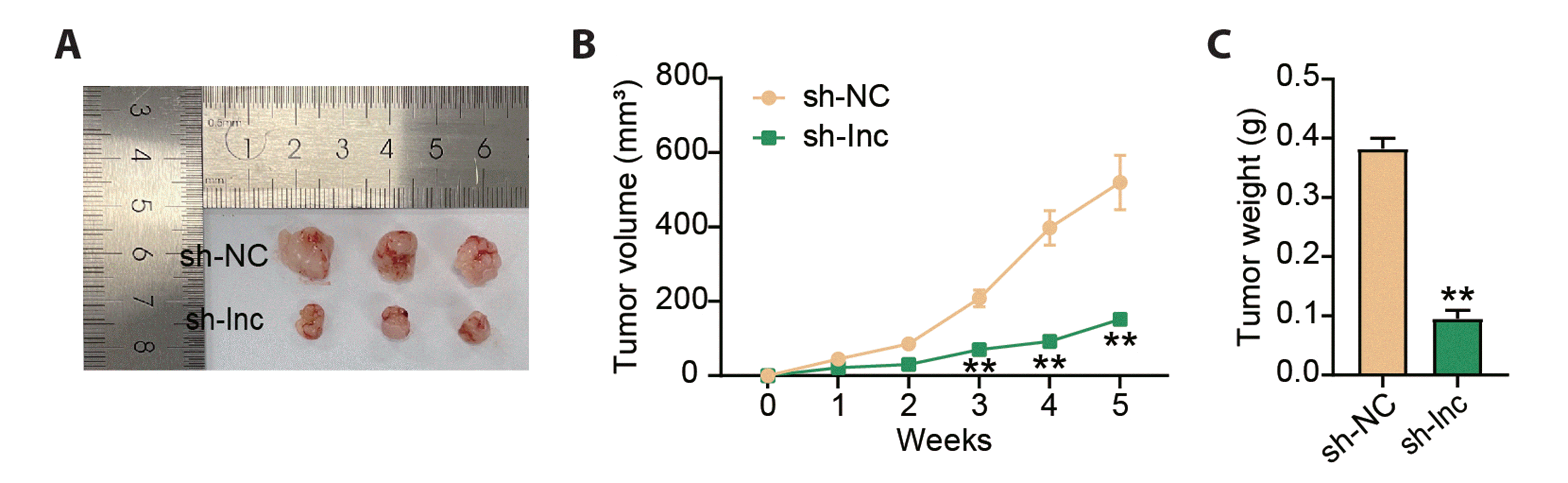
Fig. 3
Depletion of LINC00562 restrained GC tumor growth in vivo.
Nude mice (N = 6) were subcutaneously injected with HGC-27 cells that were infected with sh-lnc or sh-NC to induce tumorigenesis. (A) Representative images of tumor tissues. (B) Tumor volumes were measured once a week. (C) Tumor weight was measured after tumor growth for 5 weeks. Values are presented as mean ± SD. GC, gastric cancer. **p < 0.01 in contrast to sh-NC.
![]()
LINC00562 bound to miR-4636
Using a public database, LncBase, we analyzed miRNAs potentially targeted by LINC00562. As depicted in Fig. 4A, LINC00562 possesses binding sites for miR-4636. We validated the predicted binding sites according to the decreased luciferase activity in the experimental cells co-transfected with the miR-4636 mimic and the WT reporter vector of LINC00562 (Fig. 4B). In addition, the binding of miR-4636 to LINC00562 was confirmed by the RIP study, of Ago2-induced RISC of miR-4636 (Fig. 4C). MiR-4636 expression level considerably declined in tumor specimens of GC, relative to that in normal samples (Fig. 4D). Decreased expression level of miR-4636 was also detected in HGC-27 and AGS cells relative to that in GES-1 cells (Fig. 4E). Interestingly, miR-4636 expression was negatively correlated with LINC00562 expression in tumor samples (Fig. 4F). These data verified the targeting of LINC00562 to miR-4636.
Fig. 4
LINC00562 targeted miR-4636.
(A) LncBase (https://diana.e-ce.uth.gr/lncbasev3) predicted LINC00562–miR-4636 binding sites. (B) Putative binding sites were validated using dual-luciferase assays, **p < 0.01 in contrast to miR-NC. (C) Binding of LINC00562 to miR-4636 was confirmed by RIP assay, **p < 0.01 in contrast to Anti-IgG. (D) MiR-4636 expression levels in tumor and normal specimens. (E) MiR-4636 expression levels in GES-1, RGM-1, and GC cells (HGC-27 and AGS), **p < 0.01 in contrast to GES-1. (F) Correlation between miR-4636 expression and LINC00562 expression in tumor samples was determined using Pearson’s analysis. Values are presented as mean ± SD. GC, gastric cancer.
![]()
MiR-4636 repression rescued the inhibitory effects of LINC00562 silencing on GC cell malignant phenotypes
Next, we explored the interactional effects of LINC00562 and miR-4636 on GC cell development. MiR-4636 expression level was notably higher in si-lnc-transfected GC cells but notably lower in inhibitor-transfected cells, and miR-4636 expression was remarkably repressed by si-lnc+inhibitor co-transfection compared to si-lnc transfection alone (Fig. 5A). The function of miR-4636 was revealed; miR-4636 inhibition aggravated GC cell proliferative capacity, repressed Bax expression, enhanced Bcl-2 expression, promoted migratory capacity, and strengthened colony-formation ability (Fig. 5B–E). Notably, miR-4636 inhibition partly overturned the effects of LINC00562 absence, evidenced by the repressive effect of si-lnc+inhibitor on Bax expression and the restored effects of si-lnc+inhibitor on cell proliferation, Bcl-2 expression, cell migratory distance, and the number of colonies, in contrast to si-lnc (Fig. 5B–E). The data revealed the cancer-promoting effects of miR-4636 repression and the rescued effects of miR-4636 repression on LINC00562 absence-inhibited GC cell development.

Fig. 5
MiR-4636 inhibition reversed the inhibitory effects of LINC00562 silencing on GC cell survival.
HGC-27 and AGS cells were transfected with si-lnc, si-NC, inhibitor, inhibitor-NC, or si-lnc+inhibitor. (A) RT-qPCR was used to assess the expression level of miR-4636. (B) CCK-8 assay tested the proliferative capacity of cells. (C) Western blotting confirmed the expression of Bax and Bcl-2. (D) Wound-healing assay showed cell-migration ability. (E) Colony-formation assay was used for proliferation analysis. Values are presented as mean ± SD. ×200. GC, gastric cancer. *p < 0.05, **p < 0.01 in contrast to si-NC; #p < 0.05, ##p < 0.01 in contrast to inhibitor-NC; &p < 0.05, &&p < 0.01 in contrast to si-lnc+inhibitor.
![]()
MiR-4636 directly targeted AP1S3
Using the public database TargetScan, we analyzed mRNAs potentially targeted by miR-4636. As depicted in Fig. 6A, miR-4636 harbored a binding site in the AP1S3 3’UTR. In addition, decreased luciferase activity was identified in the experimental cells co-transfected with the miR-4636 mimic and WT reporter vector of AP1S3 (Fig. 6B). In addition, RIP analysis showed the binding of miR-4636 to AP1S3, showing upregulated expression levels of miR-4636 and AP1S3 on Ago2 (Fig. 6C). AP1S3 mRNA level was shown to be heightened in tumor specimens and cells (HGC-27 and AGS) of GC, compared to normal samples and GES-1 cell, respectively (Fig. 6D, E). Interestingly, miR-4636 expression was negatively linked to AP1S3 expression in tumor samples (Fig. 6F). These data indicated that miR-4636 binds to AP1S3 and downregulates AP1S3.
Fig. 6
MiR-4636 targeted AP1S3.
(A) TargetScan (http://www.targetscan.org/vert_72/) predicted miR-4636–AP1S3 binding site. (B) Predicted binding site was validated using dual-luciferase assays, **p < 0.01 in contrast to miR-NC. (C) Binding of AP1S3 to miR-4636 was confirmed using RIP assay, **p < 0.01 in contrast to Anti-IgG. (D) AP1S3 mRNA levels in tumor and normal specimens. (E) AP1S3 mRNA levels in GES-1, RGM-1, and GC cells (HGC-27 and AGS), **p < 0.01 in contrast to GES-1. (F) Correlation between miR-4636 expression and AP1S3 expression in tumor samples was determined using Pearson’s analysis. Values are presented as mean ± SD. AP1S3, adaptor protein complex 1 sigma 3; GC, gastric cancer.
![]()
MiR-4636 depletion strengthened AP1S3 expression and thus attenuated the anti-cancer effects of AP1S3 downregulation in GC cells
We further explored the interactive effects of miR-4636 and AP1S3 on GC cell development. The expression level of the AP1S3 protein was largely reduced in si-AP1S3-transfected GC cells but greatly increased in inhibitor-transfected cells, and its expression level was partly recovered by si-AP1S3+inhibitor relative to alone si-AP1S3 in GC cells (Fig. 7A). In terms of the function of AP1S3, we reported that AP1S3 knockdown enhanced Bax expression and restrained cell proliferation, Bcl-2 expression, cell migratory ability, and colony-formation potency in GC cells (Fig. 7B–E). Interestingly, the promotive effect of AP1S3 knockdown on Bax expression and the repressive effects of AP1S3 knockdown on cell proliferation, Bcl-2 expression, migration, and colony formation were partially attenuated by miR-4636 inhibition (Fig. 7B–E). We concluded that miR-4636 depletion aggravated the malignant phenotypes via upregulating AP1S3.

Fig. 7
MiR-4636 depletion inhibited the anti-cancer effects of AP1S3 knockdown in GC.
HGC-27 and AGS cells were transfected with si-AP1S3, si-NC, inhibitor, inhibitor-NC, or si-AP1S3+inhibitor. (A) Western blotting revealed the expression of AP1S3 protein. (B) CCK-8 assay was used to analyze the cell proliferative capacity. (C) Western blotting confirmed the expression of Bax and Bcl-2. (D) Wound-healing assay showed cell migration ability. (E) Colony-formation assay was used for proliferation analysis. Values are presented as mean ± SD. ×200. AP1S3, adaptor protein complex 1 sigma 3; GC, gastric cancer. *p < 0.05, **p < 0.01 in contrast to si-NC; #p < 0.05, ##p < 0.01 in contrast to inhibitor-NC; &p < 0.05, &&p < 0.01 in contrast to si-AP1S3+inhibitor.
![]()
DISCUSSION
Apart from functional genes, non-coding RNAs serve as key players in multiple cellular activities in the tumor microenvironment [23]. It is crucial to investigate oncogenic and tumor-suppressive lncRNAs in GC development. In this study, we identified an oncogenic lncRNA, LINC00562, which promotes cell growth, survival, and migration. MiR-4636, as a target of LINC00562, directly modulates the expression and function of downstream AP1S3. By governing the miR-4636–AP1S3 axis, LINC00562 exerts oncogenic effects in GC.
LncRNAs, which are crucial regulators that affect aggressive cell behaviors, have been extensively studied in GC. For instance, lncRNA GMAN is strongly expressed in GC tumor samples and positively linked to tumor metastasis, and knockdown of GMAN in GC cells can restrain cell invasion [24]. LINC00261 exhibits low expression levels in cancerous samples and GC cells, and its ectopic expression in GC cells represses GC cell migration and invasion, as well as in vivo metastasis [25]. The lncRNA UCA1 is overexpressed in GC tumors and cell samples, and GC cells overexpressing UCA1 are more likely to immune escape and less prone to apoptosis [26]. These views highlight that the function of lncRNAs in cancer progression largely depends on their expression pattern. In our study, the upregulation of LINC00562 was confirmed in tumor samples and GC cells. After transfecting si-LINC00562 into GC cells, we observed that LINC00562 absence restrained GC cell proliferation, migratory capacity, and colony formation but induced apoptosis. In vivo experiments, the repressive effects of LINC00562 absence on tumor growth were also observed. Our study is the first to confirm the carcinogenic effects of LINC00562 on GC development, implying that LINC00562 depletion might be a therapeutic strategy for GC. However, the association of LINC00562 expression with the clinical parameters of GC patients was not confirmed in our current study, which limited the clinical implications of LINC00562. However, this issue should be addressed in future studies.
In addition to its functional role, we explored the mechanism of action of LINC00562 in GC. We hypothesized that LINC00562 acts as a molecular sponge for miRNAs and thus investigated the miRNAs potentially targeted by LINC00562. By prediction using LncBase, miR-4636 attracted our interest because its expression level was greatly diminished in GC tumor samples and cells, which was opposite to the LINC00562 expression pattern in GC. MiR-4636 has been shown to be involved in cervical cancer, and its expression is linked to tumor volume and invasion depth [17]. Interestingly, miR-4636 expression is downregulated in GC, and miR-4636 expression restoration repressed the proliferative, migratory, and invasive capacities of GC cells [16]. Consistent with these results, we found that miR-4636 inhibition aggravated GC cell proliferation, migratory ability, colony-formation potency, and survival, indicating that miR-4636 is a tumor suppressor in GC. Overall, the role of miR-4636 in human cancers has been poorly investigated, and more studies should be conducted to verify its role in different cancers. Moreover, we observed that the repressive effects of LINC00562 downregulation on GC cell growth, migration, and survival were largely reversed by miR-4636 inhibition, confirming the interaction between LINC00562 and miR-4636.
AP1S3 is predicted by TargetScan to be a downstream target of miR-4636. Increased AP1S3 expression levels have been documented in breast cancer, glioma, and pancreatic ductal adenocarcinoma [19,20,27]. AP1S3 is targeted by miR-204-5p, and miR-204-5p represses breast cancer cell migration and invasion by depleting AP1S3 [19]. In addition, AP1S3 depletion also restrained glioma cell proliferation, migration, and invasion [20]. Following these results, we knocked down AP1S3 expression in GC cells and demonstrated that cell proliferation, colony formation, migration, and survival were repressed. These data also revealed the oncogenic effects of AP1S3 in GC. Furthermore, miR-4636 inhibition increased AP1S3 expression level and attenuated AP1S3 knockdown-induced GC growth inhibition.
In conclusion, we confirmed the aberrant upregulation of LINC00562 in GC and—for the first time—characterized the function of LINC00562 in GC, showing that LINC00562 absence restrained GC cell growth, migration, survival, and in vivo tumor growth. LINC00562 exerts carcinogenic effects in GC by targeting miR-4636 and releasing AP1S3. Functions of LINC00562 in GC should be explored further in detail.
 XML Download
XML Download