

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Many studies have indicated that caloric restriction (CR) is a good strategy to prolong life expectancy and improve life quality in most species [1,2]. CR is also thought to be beneficial for weight loss, blood glucose control, and reducing oxygen stress [3,4]. However, more and more studies have pointed out that CR should be adopted very cautiously due to its well-documented side effects on the skeletal system [5-9]. The mechanisms by which CR leads to bone loss are still not clear. Recently, many scholars have speculated that bone marrow adipose tissue (BMAT) may exert a pivotal effect on this [10]. Therefore, figuring out the effects of CR on bone metabolism and the role of BMAT in this can lead to a better understanding of CR-related bone diseases and the treatment of bone loss associated with it.
Go to : 
CALORIC RESTRICTION SUPPRESSES SKELETAL TURNOVER
In human research, there is evidence suggesting that weight loss can result in the reduced bone quantity and lower bone mineral density (BMD) in older adults [8,11,12]. In one study, 47 postmenopausal women underwent a 1-year CR trial and found weight loss caused bone loss at the ultradistal radius, 1/3 radius, and total hip [13]. Another study involving 1705 healthy and aging men followed up for 2 years showed that, compared to the people who gained weight during these 2 years, weight-loss participants lost 0.8% more of their hip BMD annually [14]. The effects of CR on younger individuals are controversial. A reduction in bone quantity has not been demonstrated in many studies in younger adults [15,16]. Nevertheless, a 2-year intervention in 218 healthy men and women whose ages were between 20 and 50 (i.e., Comprehensive Assessment of Long-term Effects of Reducing Intake of Energy [CALERIE]) concluded that the calorie restriction group had 2% lower BMD at the femoral neck, lumber spine, and total hip compared to those consuming a regular diet [9].
There is controversy regarding which skeletal areas are most affected by weight loss. Previous studies reported that the bone loss occurred mainly in the trabecular sites [13,17], while other research suggested that cortical bone was also strongly influenced in this process, especially in the weight-bearing sites [18,19]. Liu et al. [20] recently conducted a trial to examine how long-term (over 40 years) and recent (over 6 years) weight loss affected bone metabolism and discovered that both types of weight loss were related to decreased thickness and density of trabecular and cortical bone. Likewise, in rodents, food restriction can also result in reduced bone quantity and bone microstructure [21-23]. Devlin et al. [24] used young male mice (3 weeks of age) to test the effects of 30% calorie restriction on bone mass. The results demonstrated that CR mice were closely associated with lower skeletal parameters in both trabecular and cortical bone [24]. Conversely, another study using young female mice concluded that CR only suppressed cortical bone area, but increased the connectivity of trabeculae [25]. Our previous study showed that feeding female mice (8 weeks of age) a 30% CR diet for 4 and 8 weeks, respectively, showed CR only decreased cortical bone and didn’t influence trabecular bone mass [26]. These opposing results possibly reflect the different background strains of mice, age at onset of CR, sex, severity of the CR, and treatment duration used in various studies.
Except for bone quantity, bone quality is also an important factor in bone health, and several studies have pointed to a distinction between bone mass and bone quality [27,28]. Obese people with a higher weight, for example, have been shown to have compromised bone quality, yet BMD is normal or high [29-31]. In terms of CR, one study showed men who had accelerated bone loss were associated with a lower trabecular bone volume fraction (BV/TV), lower cortical thickness, and higher trabecular separation [32]. This finding was also confirmed in anorexia nervosa patients. One study on anorexia nervosa suggested that, compared with normal weight individuals, anorexia nervosa patients had decreased several related skeletal parameters both in trabecular bone and cortical bone, and this change was distinct from BMD [33,34]. However, another team’s trial concluded that moderate weight loss did not impair the bone quality of trabecular and cortical bone in obese men [35]. One possible explanation is that the subjects chosen for the studies were of varying ages or anatomical sites and responded differently to the CR. Bone strength is an important factor in the overall health of the skeleton, but we currently lack accurate methods for measuring it [4]. In animal experiments, biomechanical testing is a common method used to measure bone strength [24]. The results of three-point bending tests from one study concluded that 40% calorie restriction had little effect on bone stability and strength [36]. Another study in which male mice had caloric-restricted food for 10 weeks showed that diet restriction could reduce the ultimate force, stiffness, and second area moment of inertia of the femur [37]. Weight loss was also associated with a lower maximal force and yield point force in female rats, according to research [38]. Based on the evidence, we can conclude that different diet restriction strategies had different effects on the biomechanical properties of the bone. Moreover, even in the models that concluded food restriction caused lower bone strength, the skeleton still grew rather than stopping. In the future, more exploration and research will be needed on how to apply the bone strength-related measurements to our clinical practice.
Previously, the reduction in bone quantity and quality caused by CR was thought to be associated with weight loss. The skeleton system is a dynamic organ that can respond to various mechanical loads. Mechanical stress induced by body weight is a pivotal factor in maintaining bone homeostasis and increasing bone volume and strength [39,40]. Lower body weight has been demonstrated to have lower BMD and higher fracture incidence [41,42]. However, recent research indicates that bone mechanical stress is not the only factor affecting bone mass. Hyldstrup et al. [43] studied the effect of obesity on bone metabolism and concluded that BMD could also increase in non-weight-bearing regions. Similarly, Fogelholm et al. [44] found that BMD in non-weight-bearing regions was more closely related to weight loss than BMD in weight-bearing regions. Therefore, it is likely that other factors play an important role in the process of bone loss due to weight loss.
Go to :
CALORIC RESTRICTION INDUCES BONE MARROW ADIPOSITY
Unlike white adipose tissue (WAT) and brown adipose tissue, BMAT, another type of fat tissue in the bone marrow that accounts for more than 10% of total body fat, has not been well defined until recently [45,46]. Adipose tissue in the bone marrow is present from birth and was previously thought to be a passive filler in response to bone loss and the aging process. Recent research suggests that BMAT is important in responding to a variety of challenges, such as aging, diabetes, nutritional changes, and temperature differences [47-51]. In addition, BMAT is also a metabolic organ that can secrete endocrine cytokines to influence energy metabolism [45,52,53]. Furthermore, studies in animal models and humans have shown that there are two different types of BMAT, suggesting BMAT is a heterogeneous tissue. Regulated BMAT (rBMAT), which exists in the proximal regions surrounded by hematopoietic elements, is comprised of single, smaller adipocytes, while constitutive BMAT (cBMAT), which exists in the distal regions, features larger adipocytes but less marrow elements [54]. As noted, rBMAT has more active hematopoiesis and behaves more strongly under nutritional stress than cBMAT [54,55], but one study by Li et al. [56] suggested that 12 weeks of CR could also expand the cBMAT in the tibia, indicating that cBMAT may also have an important impact on skeleton metabolism.
It is well established that CR will decrease the mass and size of adipocytes in WAT, especially in visceral adipose tissue [57-61]. BMAT, on the other hand, accumulates in the presence of CR in both animal models and clinical diseases [24-26,55,62,63]. Anorexia nervosa is a psychiatric disease characterized by self-imposed starvation, which will cause extreme weight loss [19]. Paradoxically, clinical evidence suggests that anorexia nervosa reduces peripheral fat but increases fat tissues in the bone marrow, and when weight was regained, there was a corresponding decrease in BMAT [64,65]. Fazeli et al. [66] studied the response of BMAT to high fat and energy restriction in 23 volunteers and discovered that CR significantly increased vertebral BMAT.
Conclusions about the weight loss caused by surgery on BMAT are still controversial. Patients undergoing vertical sleeve gastrectomy showed that BMAT increases, while in people who received the Roux-en-Y gastric bypass, BMAT decreased after surgery [51,67,68]. In murine models, CR significantly increases the content of BMAT in adult and growing mice, as well as in mice of different sexes, accompanied by a decrease in serum leptin and insulin like growth factor 1 levels [24-26,55]. Our recent research also showed that 30% CR lasting for 4 or 8 weeks could induce bone marrow adiposity in both male and female mice.
To date, the mechanisms of how CR reduces bone integrity have not been fully elucidated. More research indicates BMAT may play an important role in CR-induced bone loss due to the inverse association between bone mass and marrow adipose tissue [24-26]. There are some theoretical foundations and experimental results to support this conclusion. Mesenchymal stem cells in bone marrow can give rise to various cell types like osteoblasts, adipocytes, and chondrocytes [9,69]. The accrual of adipose tissue under the condition of CR would blunt the ability of bone marrow stem cells (BMSCs) to differentiate into osteoblasts. Our research also showed that BMSCs from the CR group would form more lipid droplets but fewer calcium deposits after 9 days of adipogenic induction and 14 days of osteogenic induction. Moreover, the increased adipocytes in bone marrow could secrete some cytokines or adipokines to negatively influence the skeletal system. Peroxisome proliferator-receptor activated gamma (Pparγ), a major adipocyte transcriptional factor, increases during the CR process [55]. Wan et al. [70] found that heterozygous Pparγ +/– mice had more bone and less BMAT. In an in vitro study, Pparγ activation had an inhibitory effect on osteogenic differentiation [71-73]. Recently, adipsin, another adipokine secreted by fat tissue, was associated with bone loss [55]. Adipsin, also known as complement factor D, was secreted almost exclusively from marrow adipocytes and its expression significantly increases with CR, where the induction reached ninefold compared to the ad libitum group [55,74,75]. More importantly, when comparing the responses of wild type mice and adipsin knockout (KO) mice to CR, the researchers found that adipsin KO mice could successfully reverse the inhibitory impacts of CR on bone and decrease the adiposity of BMAT [55]. In addition to the above-mentioned reasons, many researchers have pointed out there are other related mechanisms in this process. Receptor activator of nuclear factors κB ligand (RANKL), an important cytokine for osteoclast differentiation, was previously shown to be produced primarily by osteoblasts and osteocytes [76,77], but bone marrow adipocytes (BMAds) have now been shown to be a critical source of RANKL production, affecting the balance between bone formation and bone resorption [78-80]. In humans, Attane et al. [81] showed that adipocytes in the bone marrow were devoid of lipolytic activity, which was different from classic white adipocytes. Similarly, research from Li et al. [82] concluded BMAds-induced lipolysis was essential to maintain bone mass under CR in a mouse model. Therefore, prolonged energy restriction may also contribute to the lack of bone mass by impairing the lipolysis of BMAds.
Go to :
POTENTIAL STRATEGIES TO MITIGATE THE SIDE EFFECTS OF CALORIC RESTRICTION ON BONE
Parathyroid hormone
Parathyroid hormone (PTH), a hormone from the parathyroid glands, has been convinced to have some beneficial effects on bone and is a potent therapy for osteoporosis [83-85]. Although the specific mechanism involved in PTH is not very clear, many studies have pointed out that PTH can target different cell types. For bone formation, PTH could increase osteoblast numbers and promote osteoblast activity [86-88]. PTH has also been shown to stimulate osteoclasts and bone resorption by increasing the expression of RANKL [89,90]. Besides osteoblasts and osteoclasts, PTH could affect the osteocytes to change the expressions of fibroblast growth factor-23 and sclerostin [91,92]. Furthermore, recent research has shown that PTH can recruit marrow stromal cells (MSCs) and change the balance between bone and fat by shifting the marrow lineage [93,94]. PTH treatment shifted MSCs’ commitment to the osteogenic lineage at the expense of the adipogenic lineage. Similar research suggested PTH related protein (PTHrP) could downregulate the differentiation of adipocytes, and overexpression of PTHrP in mice resulted in the accrual of osteoblasts [95,96]. Because CR caused MSCs to prefer differentiating toward adipocytes over osteoblasts, PTH treatment might mitigate the negative effects of CR on the skeletal system to some extent. Our previous study also showed that PTH administration to CR suppressed bone marrow adiposity and increased bone mass. Meanwhile, PTH promoted the lipolysis of adipocytes, and this fuel could transfer to osteoblasts during bone formation [26].
Vitamin D
Vitamin D, including vitamin D2 and vitamin D3, is hydroxylated in the liver to 25-hydoxyvitamin (25-OHD/calcidiol) and then converted in the kidney and bone cells to 1, 25-dihydroxyvitamin (1, 25(OH)D/calcitriol) [97]. Calcidiol, a major circulating form, is usually used for estimating vitamin D status, whereas calcitriol is the active metabolite for biological functions [97]. In order to function, vitamin D first needs to be combined with the vitamin D receptor (VDR), which can be expressed in various cells like BMSCs, osteoblasts, and osteoclast precursor lineages [98,99]. Vitamin D has been well recognized as having a close relationship with bone health. Vitamin D deficiency increased the risk of falling in older people, and supplementing with 700 to 800 IU of vitamin D daily can reduce this risk [100,101]. Animal evidence also suggested that VDR deficient mice have significantly impaired development and growth [102]. Although the specific mechanisms are still not very clear, it has been confirmed that calcitriol can promote the differentiation of BMSCs into osteoblasts [97,103,104]. Vitamin D also plays a role in osteoclastogenesis. Physiological doses of vitamin D could reduce bone resorption, while supraphysiological levels could increase the osteoclasts’ activity by enhancing RANKL expression [105]. Recently, studies have demonstrated calcitriol could retard the adipogenesis of bone marrow, which is consistent with the notion that bone and adipose tissue have an inverse relationship in the bone marrow [106-108]. One randomized double-blind trial studied the effect of vitamin D supplementation on the calcium level under the CR process and concluded that supplementing vitamin D during the CR could normalize serum calcium [109]. Taken together evidence suggests that vitamin D might be a potential treatment modality to decrease the inhibitory impacts of CR on the skeletal system.
Exercise
The effects of physical exercise on weight-stable individuals are controversial. Some evidence suggested that exercise-induced weight loss was harmful to bone health, while other evidence indicated that exercise had a positive impact on the skeletal system [6,110-112]. The different results may be due to the varying exercise prescriptions, the duration of the trial, and the different ages and genders chosen for the test. However, when it came to CR-induced bone loss, most studies showed that exercise, particularly resistance training, could mitigate this effect [113-115]. Emerging cellular evidence suggested exercise played a role in bone remodeling through different cell types. Exercise has been proven to enhance the proliferation and osteoblastic differentiation potential of BMSCs [116]. In this process, several signaling pathways are involved. The PTH signaling pathway could be activated by exercise, which changes the commitment of MSCs to osteoblasts at the expense of adipocytes [117]. Another pivotal mediator for bone remodeling, the Wnt signaling pathway, has also been reported to play a positive role. Exercise treatment caused β-catenin, a key component of the canonical Wnt pathway, to move to the nucleus while decreasing PPARg expression [118]. In addition to BMSCs, osteocytes are also important targets for exercise. Exercise was demonstrated to inhibit the apoptosis of osteocytes and promote the release of some cytokines like irisin that can influence bone turnover [119,120]. Irisin, a cleaved product from fibronectin type III domain-containing protein 5 (FNDC5), could increase the survival ability of osteocytes and sclerostin production through binding to its receptor, αV integrin, which suggested irisin generated by exercise might be a potential treatment target for CR-induced bone loss. Finally, there are several studies suggesting exercise can suppress bone marrow adiposity [121,122].
Go to :
CONCLUSIONS
Although CR is beneficial for human beings in regards to some tissues, its inhibitory effects on the skeletal system should be addressed. It may not only cause bone loss but also a significant decrease in bone quality, increasing the risk of fracture. As osteoporosis is a major public health problem worldwide and the cost of treating osteoporosis-related fractures has increased over the years, which makes it a heavy burden for both government and individuals, the possibility of osteoporosis due to CR is an issue that must be considered before considering treatment options.
Recently, BMAT has garnered more interest due to its increase in various pathological conditions, including CR, and has been suggested as a possible cause of bone loss. Evidence such as the fact that osteoblasts and adipocytes are derived from the same mesenchymal cells and that some cytokines secreted by adipocytes suppress osteoblast differentiation is suggesting that BMAT may become an important target for the future treatment of diet-induced bone loss (Fig. 1).
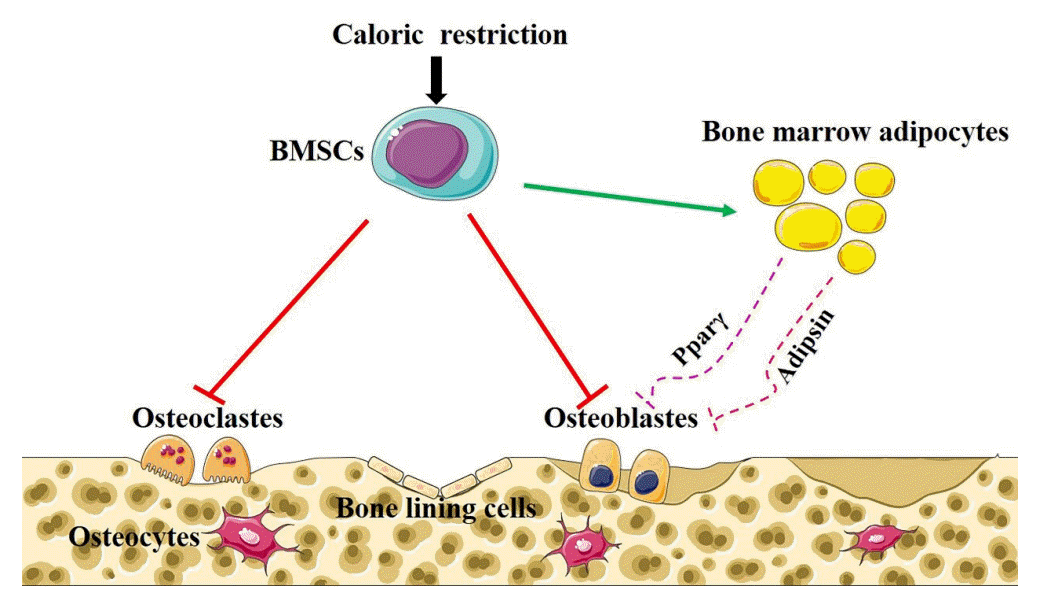 | Fig. 1.Caloric restriction induces bone loss and bone marrow adiposity. Caloric restriction induces bone marrow stem cells (BMSCs) to differentiate into adipocytes at the expense of osteoblasts, in which some adipokines such as peroxisome proliferator-receptor activated gamma (Pparγ) and adipsin play a role.
|
Go to :
 XML Download
XML Download