

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Preeclampsia (PE) is a leading cause of both maternal and infant morbidity, and mortality.1 It is characterized by the onset of maternal syndromes such as hypertension and proteinuria after 20 weeks of pregnancy. PE was diagnosed as the presence of hypertension (systolic blood pressure ≥ 140 mmHg and/or diastolic blood pressure ≥ 90 mmHg) and proteinuria (protein > 300 mg) or the absence of proteinuria according to the new guideline presented in American College of Obstetricians and Gynecologists (ACOG) in 2013.23 World Health Organization (WHO) statistics reveal nearly 10% of pregnant women suffer from PE, and PE contributes to one-seventh of the deaths in pregnant women.45
Early- and late-onset PE exhibit important clinical differences both regarding associations to fetal growth restriction as well as short- and long-term health consequences for the mother and offspring. There is no definite consensus, but clinical practice dichotomizing into preterm or term delivery (before or from gestational week 37) or into a very premature delivery or not (before or from gestational week 34) is common. The early-onset type is linked to poor placentation and fetal growth restriction, whereas maternal factors were suggested to cause the late-onset disease, without a placental impact.6789
The pathophysiology of PE is still not completely clear. However, the primary etiology includes abnormal development and function of placental cells called trophoblasts. There is poor understanding of early human placental development due to limited availability of human placenta samples. To overcome this limitation, previous studies have presented on the differentiation of human pluripotent stem cells including induced pluripotent stem cells (iPSCs) into trophoblast lineage cells of early pregnancy.7101112 Therefore, this study used trophoblast stem-like cells (TSLCs) from iPSCs as a model of an early pregnancy with evident characteristics of early-onset PE.1 The placenta is a highly vascularized organ and serves as a maternal blood store.13 Thus, maternal gene expression changes can provide information of biomarkers for early detection or management of the disease.131415 Therefore, our study used maternal peripheral blood sample from 35 weeks pregnant women diagnosed with PE and normal pregnant woman as models of the late term pregnancy, respectively.
Oxidative stress is related in the pathogenesis of PE, with the huge production of reactive oxygen species (ROS) and hypoxia-induced placental oxygen tension oscillations being the hallmark to ROS production.161718 Mitochondria are involved in the regulation of cellular metabolism and the production of ROS. Mitochondrial matrix-related NME/NM23 nuceloside diphosphate kinase 4 (NME4) gene is known to have the ability to supply nucleotide triphosphate (NTP) and deoxynucleotide triphosphate (dNTP) for replication and transcription of mitochondria. In addition, previous study reported changes in mitochondrial structure and function in trophoblasts of PE, resulting in ROS overproduction, oxidative stress and cell death.19
Cell death plays a crucial role in the regulation of trophoblast life span. Exaggerated cell death shows high correlation to the increased generation of trophoblast debris, which sheds into the maternal circulation and may trigger the clinical manifestations of PE.182021 Therefore, mitochondrial dysfunction-induced cell death might be a promising therapeutic target for PE management.
Our study aimed to investigate candidate genes associated with the mechanism of PE using patient-specific TSLC in a model of early pregnancy and to confirm the expression of candidate genes in maternal peripheral blood mononuclear cells (PBMNCs) in the model of late preterm pregnancy. Since differential of gene expression appear earlier than the onset of the disease, we attempted to discover biomarkers for early diagnosis of preeclampsia using transcriptome analysis.
METHODS
Generation of iPSCs from fetal-origin amniotic epithelial cells
For patient-derived iPSCs, human placentas were attained from three normal pregnant women and three pregnant women with preeclampsia. PE was diagnosed as according to the presence of hypertension (systolic blood pressure ≥ 140 mmHg and/or diastolic blood pressure ≥ 90 mmHg) and proteinuria (protein > 300 mg) or absence of proteinuria according to the new guideline of ACOG in 2013.12322 Briefly, the samples used as iPSCs in our study have clinical properties of early-onset PE, such as hypertension (170/100, 150/90, and 160/100 mmHg) and proteinuria. In addition, these samples have genes associated with very premature delivery (32,34,35 weeks), low birth weight (1,400, 1,710, and 2,300 g), and extracellular matrix (ECM) receptor interactions, focal adhesion pathways-related genes that are different from normal.1 The process of fetal-origin amniotic epithelial cells (AECs) from human placentas has been previously reported.1
Vector mixtures (total 3 μg; Thermo Fisher Scientific, Waltham, MA, USA) encoding the four Yamanaka factors of Oct3/4, Sox2, Klf4, c-Myc, were electroporated using a microporator system (Neon; Invitrogen, Carlsbad, CA, USA) in accordance to the manufacturer’s instructions. After three pulses with a voltage of 1,650 for 10 ms, the cells were grown in Dulbecco’s modified Eagle medium containing 10% fetal bovine serum (FBS). Seven days after transfection, the cells were transferred onto ES-Matrigel (Corning Inc., Corning, NY, USA) coated well. Approximately one month after transfer onto ES-Matrigel, the iPSC colonies similar to human embryonic stem cells (ESCs) were manually selected and further cultured for characterization. Human iPSCs (Normal #1–#3, and patient #1–#3) generated in this study were cultured in Essential 8 (E8) medium (Life Technologies, Carlsbad, CA, USA). Cells were passaged every 5–6 days using dispase (1U/mL) (Stem Cell Technologies, Vancouver, Canada). Cells were grown in a humidified incubator at 37°C with 5% CO2.
Characterization of iPSCs
To characterize iPSCs, we performed alkaline phosphatase (ALP) activity using ALP staining kit (Sigma-Aldrich, St. Louis, MO, USA) and immunostaining. To confirm the pluripotent stem cell characteristics by immunostaining, the cells were fixed in 4% paraformaldehyde solution, permeabilized with 0.2% Triton X-100, and stained with OCT4 (1:100; Santa Cruz Biotechnology, Dallas, TX, USA) and SSEA-4 (1:100; Abcam, Cambridge, UK) antibodies. Embryoid bodies (EBs) formed by partial dissociation of iPSCs using collagenase type IV (Invitrogen) were transferred to ultralow attachment plates (Corning Inc.) and cultured in DMEM/F12 medium (Thermo Fisher Scientific) supplemented with 20% knockout serum (Invitrogen), 4.5 g/L L-glutamine, 1% nonessential amino acids, 0.1 mM 2-mercaptoethanol, and 5% FBS. After a week of cultivation under these conditions, EBs were attached onto Matrigel-coated culture dishes and further cultured for 10 days. Spontaneous differentiation of EBs into cells representing the three germ layer lineages was detected by immunostaining with markers for ectoderm (Nestin [1:100; Abcam] and Pax6 [1:400; Abcam], mesoderm [Brachyury, 1:100; R&D Systems, Minneapolis, MN, USA]), and endoderm (α-fetoprotein; AFP [1:100; Invitrogen]) antibodies.
Differentiation and characterization of TSLCs from iPSCs
To induce TSLCs, iPSCs were directly transferred onto Matrigel-coated dishes, and cultured with bone morphogenic protein 4 (BMP4) (Proteintech, Chicago, IL, USA) for 8 days. The BMP4/E8 medium was changed every alternate day. To confirm the characterization of the TSLCs, the cells were plated onto the Matrigel-coated coverslips and differentiated for 8 days with BMP4 (10 ng/mL) before fixation in 4% paraformaldehyde. Coverslips were placed in 5% bovine serum albumin + 2.5% v/v donkey serum for 1 hour to block non-specific staining and then incubated overnight at 4°C with cytokeratin-7 antibody (1:400; MilliporeSigma, Burlington, MA, USA). The most commonly used trophoblast markers reported in the literature are cytokeratin 7 (CK7), human leukocyte antigen-G, and human chorionic gonadotrophin. Especially, CK7 was proposed as a marker because trophoblast is the only epithelial cell in the placenta. Given that the model of TSLCs was derived from human AECs in our study, CK7 was used to identify and confirm TSLC character. After incubation with secondary antibody (Alexa Fluor® 488 goat anti-rat, 1:400; Invitrogen) for 1 hour at room temperature as around 20–22°C, coverslips were mounted with Vectashield® 4,6-diamidino-2-phenylin-dole (DAPI).
PBMNCs isolation
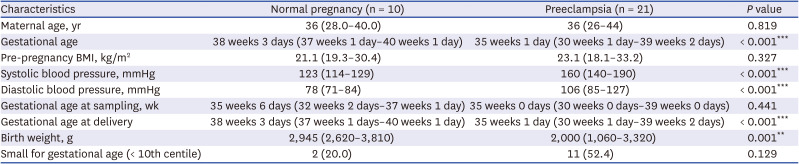
Human peripheral blood samples were obtained from women with normal (n = 10) or PE pregnancies (n = 21) via the approval of the IRB of Severance Hospital. Yonsei University Health System approved this consent procedure (No. 4-2017-0348). Inclusion criteria for pregnancy with PE in the Introduction section of this article.12322 The inclusion criteria for normal pregnancy without PE are featured with normal pregnancy process, normal blood pressure (under 140/90 mmHg). All participants are natural and singleton pregnancy. Blood samples were drawn from pregnant women diagnosed with PE and normal pregnant woman respectively, followed immediately by isolation of PBMNCs (Table 1).
Table 1
Clinical features of enrolled subjects for preparation of PBMNCs
PBMNCs were prepared using Ficoll-Paque following the manufacturer’s protocol (Amersham Biosciences Corporation, Piscataway, NJ, USA) within 2–3 hours of collection. 5–10 mL of the blood sample was overlaid on top of a 5 mL Ficoll gradient and centrifuged at 400 × g for 30 minutes. The top white interface containing PBMNCs was collected and diluted at 1:10 with PBS and centrifuged again for 10 minutes at 100 × g. PBMNCs pellets were saved in aliquots for further characterization.13
RNA extraction
Total RNA was extracted as previously described.1 It was extracted from TSLCs s and PBMNCs using TRIzol reagent (Invitrogen) according to the manufacturer’s protocol and stored at −80°C until further use. For quality control, RNA purity and integrity were estimated by optical density 260/280 ratio was confirmed with a Nanodrop spectrophotometer (Thermo Fisher Scientific). Purified RNA (1 µg) was used as a template to generate cDNA using the ReverTra Ace qPCR RT master mix with gDNA remover (TOYOBO, Osaka, Japan).
Transcriptome analysis of TSLCs from iPSCs
We performed transcriptome analysis by RNA sequencing (Macrogen Inc., Seoul, Korea) to identify genes that were differentially expressed in patients with PE compared to normal individuals. Functional annotation clustering was found to be significantly up- or down-regulated in the TSLCs s of patients with PE using Database for Annotation, Visualization, and Integrated Discovery (DAVID) software (http://david.abcc.ncifcrf.gov/). These analyses were checked with an enrichment score of > 1.7 for the data set analyzed. We performed real time-polymerase chain reaction (qRT-PCR) to validate candidate gene using models of the early and late term pregnancy of PE.
The qRT-PCR was performed on the StepOnePlus™ Real Time PCR System (Thermo Fisher Scientific) using the qPCRBIO SyGreen mix Hi-Rox (PCR Biosystems, London, UK), according to the manufacturer’s instructions. A total volume of 20 μL master mix prepared with 1 μL of cDNA was used in the qRT-PCR reaction, which was performed in triplicate on a LightCycler 480 using the LightCycler 480 SYBR Green master mix (Roche Applied Science, Mannheim, Germany). Thermal cycling conditions were 95°C for 10 minutes followed by 40 cycles of 95°C for 15 seconds and 60°C for 1 minute. Primers used for qRT-PCR were as follow: NME4 primer was purchased (HP103265, Sino Biological, Beijing, China) and used. p53 were 5’-TGC GTG TGG AGT ATT TGG ATG-3’ and 5’-TGG TAC AGT CAG AGC CAA CCT C-3’. Glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as an internal control. GAPDH were 5’-AAG GGT CAT CAT CTC TGC CC -3’ and 5’-GTG AGT GCA TGG ACT GTG GT-3’. The expression of each gene of interest was determined using the 2−ΔΔCt method. All results are expressed as mean ± standard error of the mean (SEM) from at least three independent experiments.
Western blotting
Proteins were extracted from iPSCs and TSLCs were by radioimmunoprecipitation assay buffer (Invitrogen) containing protease inhibitor and phosphatase inhibitors (both from Invitrogen). Thirty micrograms of total protein were separated by sodium dodecyl sulfate polyacrylamide gel electrophoresis gel and transferred to 0.45 µm nitrocellulose membranes (Millipore). Blots were blocked with 5% skimmed milk in Tris-buffered saline for 1 hour, incubated with primary antibodies at 4°C overnight, and then incubated with horseradish peroxidase-coupled secondary antibodies at 37°C for 1 hour. The primary antibodies used as follow: NME4 (1:1,000; Novus Biological, Centennial, CO, USA), p53 (1:1,000; Santa Cruz Biotechnology), thioredoxin (TRX) (1:250; Abcam) and β-actin (1:1,000; Santa Cruz Biotechnology).
Terminal deoxynucleotidyl transferase (TdT)-mediated deoxyuridine triphosphate (dUTP) nick end labelling (TUNEL) assay
The levels of cell death in PE were evaluated using a clinical recombinant TUNEL kit (G3250; Promega, Madison, WI, USA). TSLCs from iPSCs were fixed in fresh 4% paraformaldehyde, permeabilized with 0.2% Triton X-100, and incubated with recombinant TdT buffer for 60 minutes at 37°C. The reaction was terminated by washing with 2× SCC solution. The TSLCs from iPSCs were counterstained with DAPI and visualized using a fluorescence microscopy.
Statistical analysis
All experiments were performed with at least three replicates in each group, and each experiment was repeated three times. Data are presented as mean ± SEM. Statistical analyses were conducted using the Statistical Package for Social Sciences (SPSS) version 25.0 (IBM Corporation, Armonk, NY, USA). An independent t-test was performed to compare the means between the two groups. Statistical significance was set at P < 0.05.
Ethics statement
Approval for the research protocol was granted by the Ethics Committees of Severance Hospital, Yonsei University Health System (No. 4-2013-0344, 4-2017-0348 and 4-2017-0450). Written informed consent was provided for participants in this study, and the Institutional Review Board approved this consent process. All participants provided informed consent for placental tissue collection (No. 4-2013-0344 and 4-2017-0450) and maternal peripheral blood (No. 4-2017-0348) collection.
RESULTS
Characterization of patient-derived iPSCs and TSLCs from iPSCs
We constructed iPSCs from human AECs (hAECs) of normal and PE patients. Specifically, PE patients who provided hAECs gave birth at the 32–35 weeks of pregnancy, and the baby was underweight. Our previous study mentioned detail information about characteristics of enrolled subjects for iPSCs from hAECs.1 In addition, previous our study1 reported that genes related to ECM receptor interactions and focal adhesion pathways in the PE environment are different from normal and samples. Taken together, the TSLCs of PE in our study have the clinical properties of early-onset PE.
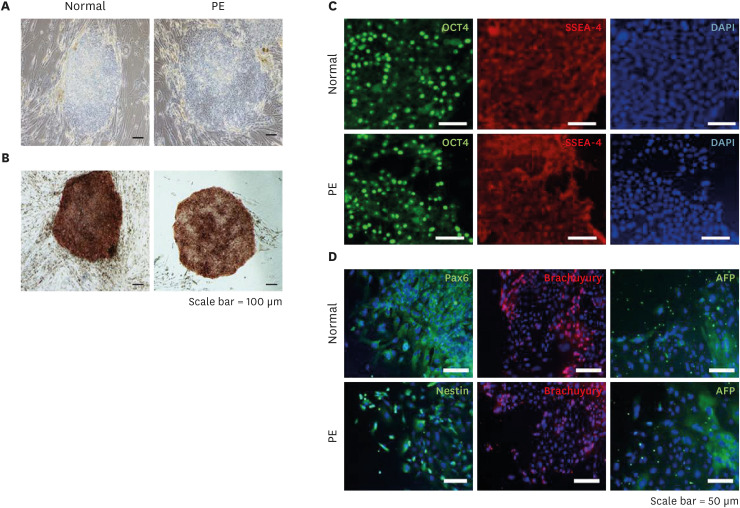
To confirm the characterization of iPSCs, we showed colonies exhibiting morphology and stained ALP activity (Fig. 1A and B). These iPSC colonies expressed typical ESC markers such as OCT4 and, SSEA-4 (Fig. 1C). Next, we confirmed the pluripotency of iPSCs via three-germ layer staining used EBs. As expected, markers for ectoderm (Nestin and Pax6), mesoderm (Brachyury), and endoderm (AFP) lineages were expressed in the differentiated cells from EBs (Fig. 1D). These data indicate that the iPSC lines derived from hAECs have pluripotency similar to human ESCs.
Fig. 1
Generation and characterization of iPSCs derived hAECs. (A) The morphology of iPSCs derived from hAECs of normal and PE was confirmed. (B) ALP staining was used to characterize iPSCs from normal and preeclampsia patients (scale bar = 100 μm). (C) To confirm the characteristics of iPSCs, the expression of OCT4 and SSEA-4, which are human ESC-specific markers, were confirmed by immunocytochemistry. (D) In order to confirm the pluripotency of iPSCs, the expression of markers representing ectoderm (Pax6 or Nestin), mesoderm (Brachuyury), and endoderm (AFP) were confirmed by immunocytochemistry. Each marker was merged with DAPI (scale bar = 50 μm).
iPSC = induced pluripotent stem cell, hAEC = human amniotic epithelial cell, PE = preeclampsia, ALP = alkaline phosphatase, ESC = embryonic stem cell, AFP = α-fetoprotein, DAPI = 4,6-diamidino-2-phenylin-dole.
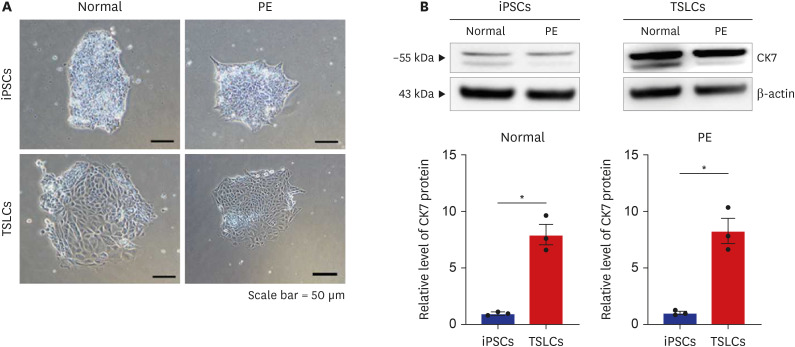
To investigate whether iPSCs could be differentiated into TSLCs by BMP4 treatment (10 ng/mL), our study confirmed changes in morphology from iPSCs to induced TSLCs via microscope (Fig. 2A). Trophoblast marker CK7 expression in TSLCs from Normal (P = 0.002) and PE (P = 0.003) significantly increased compared to iPSCs via western blot analysis (Fig. 2B). These results demonstrate that human iPSCs can differentiate into TSLCs following BMP4 treatment.
Fig. 2
Differentiation and characterization of TSLCs derived iPSCs. (A) The change of morphology in TSLCs s through BMP4 treatment was confirmed (scale bar = 50 μm), (B) western blotting confirmed that the expression of CK7, human trophoblast marker, was significantly increased in both the normal and PE groups after BMP4 treatment. Results are expressed as means ± SEM from three independent experiments.
TSLC = trophoblast stem-like cell, iPSC = induced pluripotent stem cell, BMP4 = bone morphogenic protein 4, CK7 = cytokeratin 7, PE = preeclampsia, SEM = standard error of the mean.
*P < 0.05 versus normal group.
Characteristics of enrolled subjects for preparation of PBMNCs
A total of 21 patients with preeclampsia and 10 healthy controls were included in this study for PBMNCs experiment. The basic clinical information was described in Table 1. There were no significant differences between the two groups in terms of maternal age, pregnancy body mass index, gestational age at delivery (weeks). Both average of the systolic (160 mmHg) and diastolic blood pressures (106 mmHg) in PE were significantly higher than those in normal pregnancy (systolic blood pressure, 123 mmHg and diastolic blood pressure, 78 mmHg) (P < 0.001). The average of birth weight (g) in the PE (2,000 g) was significantly lower than that in normal pregnancy group (2,945g) (P = 0.001). There were 2 low-birth weight babies, ranging from 10% centile in the normal group to Korean standards,23 a constitutionally small fetus with no pathological problems. In addition, weight of 2,945 g at 35 weeks of birth is equivalent to 50% centile at 38 weeks of birth, which is not a low birth weight when viewed by the Korean single-fetal infant standard.23
Upregulation of p53 and thioredoxin in PE
p53 plays a crucial role in regulating cell death and cell cycle arrest by activating or repressing various downstream target genes.2425 Cell death plays a crucial role in the regulation of trophoblast life span. In this study, protein and mRNA levels of p53 were determined in TSLCs and PBMNCs using qRT-PCR and western blotting. In patients with PE, expression of p53 was significantly elevated in TSLCs (P = 0.002) (Fig. 3A) and PBMNCs (P = 0.011) (Fig. 3B). TUNEL assay also confirmed that apoptotic cells were significantly increased in the PE group compared to the normal group (P = 0.024) (Fig. 3C).
Fig. 3
Cell death-related expression of p53 upregulated in PE. (A) Western blotting result showed that the expression of p53 was significantly increased in TSLCs (B) qRT-PCR result showed that the expression of p53 was significantly increased in PBMNCs of PE (C) Clinical TUNEL staining was employed to evaluate the level of cell death (green, TUNEL, DAPI nuclear staining) in TSLCs. TSLCs of PE group exhibited increased number of TUNEL positive rate which was normalized to the normal group (scale bar = 20 μm) (D) Western blotting result showed that the expression of TRX tend to increase in TSLCs of PE. Results are expressed as means ± SEM from three independent experiments.
PE = preeclampsia, TSLC = trophoblast stem-like cell, qRT-PCR = quantitative reverse transcription-polymerase chain reaction, PBMNC = peripheral blood mononuclear cell, TUNEL = deoxynucleotidyl transferase (TdT)-mediated deoxyuridine triphosphate (dUTP) nick end labelling, DAPI = 4,6-diamidino-2-phenylin-dole, TRX = thioredoxin, SEM = standard error of the mean.
*P < 0.05 versus normal group.
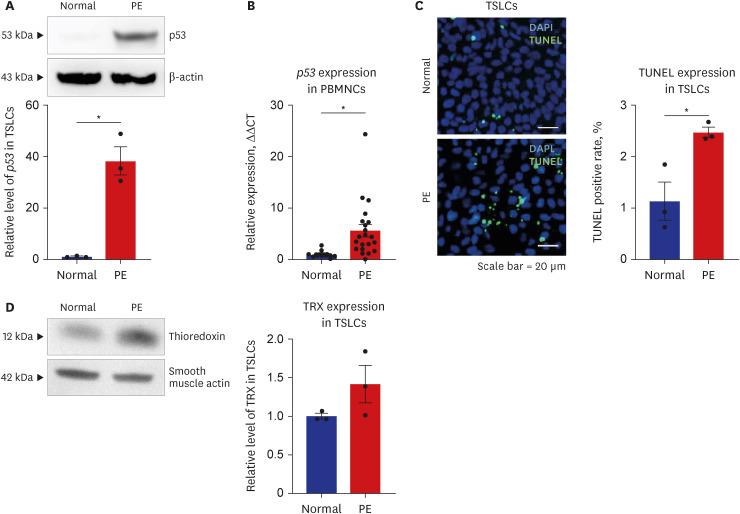
TRX protects cells from oxidative stress and regulates cell activity under various oxygen conditions. Previous studies have reported that TRX is rich in the human placenta, suggesting its protective role in trophoblasts.262728 Our results also showed that the oxidation-reduction process-related protein TRX tended to be upregulated in TSLCs (Fig. 3D) in PE. Similar to our results, a recent study showed that TRX levels increased under hypoxic conditions.28
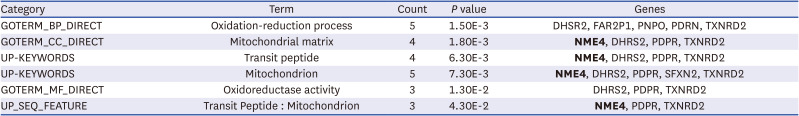
Expression of mitochondrial matrix-related gene, NME4, in PE
Transcriptome analysis was performed to identify genes differentially expressed TSLCs from preeclamptic patients and controls. Total 7,515 differentially expressed transcripts were obtained in approximate. Functional annotation clustering was analyzed via DAVID analysis program. Functional annotation clustering analysis confirmed that mitochondrial matrix-related genes, such as NME4, were downregulated in PE. We performed qRT-PCR to validate of NME4 in TSLCs and PBMNCs of normal pregnancy and PE. In particular, the oxidation-reduction process and mitochondrial matrix-related genes were downregulated in PE (Table 2). Our study verified whether these down-regulated genes might play a role in the pathophysiology of PE. Previous studies have reported that the syncytiotrophoblasts in PE show altered mitochondrial structure and function resulting in ROS overproduction, oxidative stress and cell death.19 Therefore, our study focused on elucidating the relationship between the mitochondrial matrix genes, NME4 and PE.
Table 2
Functional annotation clustering in the TSLCs from preeclamptic patients using DAVID software
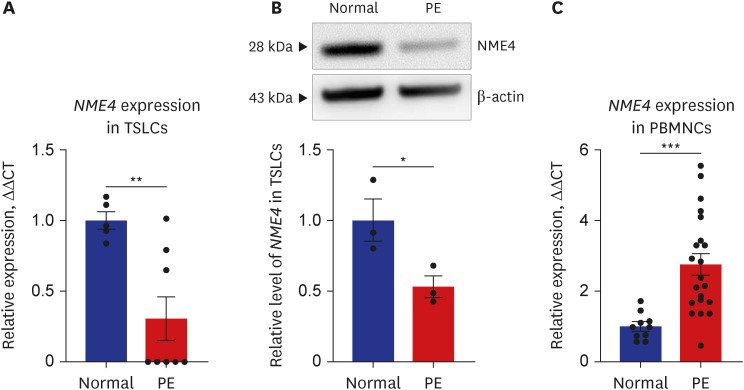
We confirmed that the mitochondrial matrix-related gene NME4 was also downregulated at the RNA (P = 0.006) and protein level in TSLCs of PE (P = 0.046) (Fig. 4A and B). However, the expression of NME4 in PBMNCs of PE significantly increased at the late term pregnancy in contrast to the early pregnancy (P < 0.001) (Fig. 4C). In addition, we analyzed subgroup in the case of no proteinuria in the diagnosis inclusion criteria. Three PE patients were classified into no proteinuria group. Total 21 PE patients, three were no proteinuria and 18 were classified into the proteinuria groups. There were no significant differences between the two groups in expression level of NME4.
Fig. 4
NME4 expression decreased in a model of early pregnancy and increased in a model of late preterm pregnancy of PE. (A) qRT-PCR results showed that the expression of NME4 was significantly decreased in TSLCs. (B) western blotting result showed that the expression of NME4 was significantly decreased in TSLCs. (C) qRT-PCR result showed that the expression of NME4 was significantly increased in PBMNCs of PE. Results are expressed as means ± SEM from three independent experiments.
NME4 = NME/NM23 nuceloside diphosphate kinase 4, PE = preeclampsia, qRT-PCR = quantitative reverse transcription-polymerase chain reaction, TSLC = trophoblast stem-like cell, PBMNC = peripheral blood mononuclear cell, SEM = standard error of the mean.
*P < 0.05, **P < 0.01, ***P < 0.001 versus normal group.
The collaborative results from our study displayed downregulation of the mitochondrial matrix-related gene NME4 in early-onset PE and upregulation of p53 and TRX in hypoxic environments following abnormal placenta development.
DISCUSSION
PE-related cellular and/or molecular abnormalities occur between 8 and 18 weeks of gestation. The inability to obtain placental tissues from early pregnancy stages of gestation and the delayed clinical manifestations of PE represent the problems in the study of this disease.29 Therefore, most studies of PE provide information about the clinical PE stage, as most depend on the placentas obtained after delivery. To overcome these problems, recent studies have optimized a system to induce differentiation into trophoblast lineage cells and have optimized conditions for efficient conversion.1130 Therefore, in this study, we used a trophoblast lineage differentiated from iPSCs through BMP4 treatment. Our study confirmed the characteristics of trophoblast lineage cells using CK7 since the model of early pregnancy was derived from human AECs. However, our study has a limitation to confirm the characteristics of trophoblast lineage cells only with CK7. Therefore, a further study is needed to identify with another trophoblast lineage cell markers. Our previous study1 also reported that human AECs from PE patients have clinical characteristics of early-onset PE such as hypertension and proteinuria. In addition, these samples have very premature delivery (before or from gestational week 34), low birth weight (under 2,500 g) and ECM receptor interactions, focal adhesion pathways-related genes that are different from normal. Taken together, TSLCs from human AECs samples in this study have clinical characteristics of early-onset PE.
Placental electron microscopic ultrastructural studies have reported abnormal mitochondrial morphology in preeclamptic placentas.31323334 Mitochondrial dysfunction begins in the early stages of pathogenic conditions with oxidative stress in PE. Changes in mitochondrial structure and function in trophoblasts of PE, resulting in ROS overproduction, oxidative stress, and cell death.19 In previous study, cytotrophoblast cells of the placenta from PE displayed mitochondria vacuolization under the electron microscope, which is a sign of mitochondrial swelling and cell injury.33 Mitochondrial swelling is the result of increased oxidative stress damage to lipids in the mitochondrial membrane.353637
Therefore, we suggest that mitochondrial membrane damage by oxidative stress in PE induced downregulation of mitochondrial matrix-related gene, NME4, in pregnancy period.
In general, mitochondria usually originate from the maternal line. During development, sperm do not contribute to mitochondria. Previous studies reported that increases of mitochondrial DNA (mtDNA) was noted from blood samples obtained from preeclampsia patients.3839 In addition, mtDNA copy number are highly sensitive to oxidative stress have been suggested as sensitive indicator of cellular oxidative stress, mitochondrial dysfunction.4041 Oxidative stress can increase mtDNA copy number synthesis as a compensatory mechanism to ensure cell survival. Mild oxidative stress can first respond to increased energy demands by increasing mtDNA copy number.414243 However, continued exposure to oxidative stress can cause mtDNA damage, reducing or limiting the replication of mtDNA.4244
Our results showed that the mitochondrial matrix-related gene, NME4, significantly decreased in TSLCs as a model of early pregnancy in PE. Although few studies about function of NME4 has been reported, previous study mentioned that NME4 plays a major role in the synthesis of nucleoside triphosphates other than ATP45 and acts as a mitochondria-specific nucleoside-diphosphate kinase.46
In our study, TSLCs and PBMNCs were used as models mimic PE at early and late term pregnancy. There is poor understanding of early human placental development due to limited availability of human placenta samples. To overcome this limitation, previous studies have presented on the differentiation of human pluripotent stem cells including iPSCs into trophoblast lineage cells of early pregnancy.7101112 Therefore, this study used TSLCs from iPSCs as a model of an early pregnancy with evident characteristics of early-onset PE.1 The hypoxia condition in placenta from PE may stimulate the increase of apoptotic signal and the release of placental factors, including inflammatory cytokines and lipid peroxides, ROS, into maternal circulation.474849 Analysis of blood cells in experiments including humans appears to be a new research field with potential predictive biomarkers in several conditions.5051525354555657 Especially, maternal gene expression changes can provide information of biomarkers for early detection or management of the disease.131415 Therefore, our study used maternal peripheral blood sample from 35 weeks pregnant women diagnosed with PE and normal pregnant woman as models of the late term pregnancy, respectively.
To confirm whether NME4 can be generalized as a predictive marker of preeclampsia, maternal blood samples were drawn from 35 weeks pregnant women diagnosed with PE and normal pregnant woman respectively, followed immediately by isolation of PBMNCs (Table 1). To investigate that down-regulated NME4 at early pregnancy was maintained until the late preterm pregnancy of PE.
Interestingly, our results showed that the expression of NME4 at the late preterm pregnancy of PE was significantly increased in PBMNCs. Reduced utero-placental flood flow has been observed in cases of PE with hypertension.5859 In particular, histological changes in various physiological circumstances including narrowing of capillaries have been demonstrated.3358 Previous study showed that umbilical cord blood mtDNA copy number of PE patients were much lower than in normal controls, in contrast to the results of maternal blood. This study described that the fetus was a very vulnerable period to oxidative stress, mitochondria exceeded the coverage range in cord blood of PE patients, resulting in a decrease in the number of replicates.41
These results were similar to our findings, where the amount of mitochondrial matrix-related gene, NME4, in PE maternal blood and fetal side derived TSLCs were opposite. However, in this study, mitochondrial function by downregulation of NME4 level in TSLCs were not thoroughly investigated. Therefore, a further study to elucidate this association is needed.
As a key regulator in multicellular organisms, p53 plays a crucial role in regulating cell death and cell cycle arrest by activating or repressing various downstream target genes.2425 In this study, the expression of p53 significantly increased in the model of early and late preterm pregnancy of PE. TUNEL assay results also showed that apoptotic cells significantly increased in the PE group compared to the normal group. Based on the results of the study, it was hypothesized that that cell death occurs both in the early and late preterm pregnancy due to downregulation of mitochondrial matrix gene expression. However, it has not been thoroughly investigated for NME4 mediated cell death. Therefore, further study will need to determine whether normalization of NME4 expression leads to decreased p53 expression.
Many studies mentioned that dysfunction of the placenta in PE induced prolonged hypoxic stress.6061 Our results showed that oxidation-reduction process related protein TRX tend to up-regulated in model of early pregnancy of PE. Previous studies have reported that TRX is rich in the human placenta, suggesting its protective role in trophoblasts.262728 In addition, TRX levels increased under hypoxic conditions.29 Based on these results, we hypothesized that the increase in TRX expression would be a compensatory response of hypoxia by abnormal placenta in early-onset PE.
This study elucidated an early pregnancy model of PE and confirmed that NME4 expression, a mitochondrial-associated gene, was reduced in that model. In addition, it was confirmed that cell death increased from the model of early pregnancy and the oxidation-reduction process related protein increased. However, whether NME4 can actually act as a key predictive marker requires further studies. This includes the relationship between NME4 expression and mitochondrial function, and NME4-mediated cell death while also obtaining NME4 expression patterns throughout the entire gestation period.
This study confirmed that the expression of the mitochondrial matrix-related gene NME4 differs between early pregnancy and late preterm pregnancy of PE, and suggests that this expression pattern may be a potential biomarker for early diagnosis of PE. However, whether NME4 can actually act as a key predictive marker requires further studies.
 XML Download
XML Download