

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Pain is a predominant problem in modern healthcare [1]. In addition to diminishing patient quality of life, pain also exerts immense financial costs [2,3]. For example, nearly 50–100 million Americans live in constant pain, with an estimated annual cost of $635 billion [4]. Opioid analgesics such as morphine remain the major treatment modality for moderate to severe pain; however, most chronic pain patients develop opioid tolerance after long-term opioid use [5]. Although many studies have focused on opioid analgesic tolerance, the underlying mechanisms of the initiation and development in opioid tolerance remain unclear because of their complexity.
Synaptic plasticity participates in the transmission and regulation of pain signals [6,7]. Opioids largely influence the synaptic plasticity of learning and memory within the neural system that are crucial to analgesic tolerance development [8]. Calcium/calmodulin-dependent protein kinase II (CaMK II) plays a fundamental role in neural plasticity, long-term potentiation, learning, and memory [9,10]. Acute or chronic morphine exposure enhances CaMK II expression in the prefrontal cortex or spinal cord [11,12]. In rodent models of morphine tolerance, administration of CaMK II inhibitors into the spinal cord, hippocampus, or prefrontal cortex not only prevented, but also reversed morphine tolerance. This occurs either directly or through learning/memory pathways [10,11,13,14]. The above results imply that CaMK II is essential to the development and retention of morphine tolerance.
The nucleus accumbens (NAc) is a limbic structure that is associated with reward, addiction, and pleasure. Recent pain studies have shown that the NAc also plays a major role in pain modulation and antinociceptive tolerance and acts as a source of analgesia [1,15,16]. The authors’ earlier report showed that the microinjection of the CaMK II inhibitor autocamtide-2-related inhibitory peptide (AIP) or KN93 into the NAc alleviates pain that is related to behaviors in the peripheral neuropathic pain model [17,18]. However, the role of CaMK II in the NAc in naïve and morphine-tolerant rats has yet to be the subject of in-depth examination. Therefore, the current research was conducted to determine whether CaMK II participates in the transmission of nociception in the NAc of naïve rats and to examine the role of CaMK II in rats exhibiting morphine tolerance.
Go to : 
MATERIALS AND METHODS
1. Chemicals and drugs
The solutions for NAc microinjection were dissolved in 0.9% sterilized saline, and the drug concentration was 4, 8 or 12 µg of AIP (a selective inhibitor of CaMK II; GL Biochem) per milliliter. The experimental group was injected with 1 µL of 0.9% sterilized saline.
2. Animal preparation
In this study, adult male Sprague–Dawley rats with weights within the range of 200–250 g (Experiment Animal Center of Kunming Medical University) were used. These were housed in cages, provided with ad libitum access to food and water and under a 12-hr reversed light/dark cycle, and maintained in a 22°C–24°C room. The experimental protocols both complied with the Guidelines for the Care and Use of Laboratory Animals of the National Institutes of Health and fulfilled the requirements of the Institutional Animal Care and Use Committee of Kunming Medical University (no. Kmmu 20221212).
3. Chronic morphine tolerance model
Rats received intraperitoneal morphine injection (10 mg/kg, morphine hydrochloride injection, Shenyang First Pharmaceutical Factory) twice a day (09:00 and 21:00) for a total of seven days for induction of chronic morphine tolerance. To determine antinociceptive tolerance, the authors assessed the effect of the antinociception after 30 minutes of the first morphine injection every day, which was also compared with that of the first day. The increase in hindpaw withdrawal latencies (HWLs) in response to noxious thermal and mechanical stimulation due to morphine markedly decreased after repeated use of morphine. After seven days of repeated injection, the chronic morphine tolerance rat model was established.
4. Nociceptive tests
Before the experiment, the rats were subjected to behavioral test training using noxious thermal and mechanical stimuli for three to five days. The HWLs, in response to noxious thermal and mechanical stimuli, were examined as described elsewhere [18–20].
In the experiment, HWLs, in response to nociceptive thermal stimulation were assessed by a hot-plate (Type YSL-6B Intelligent Heat Panel Instrument) that was set to a temperature of 52 ± 0.2°C. Radiant heat stimulation was applied to the plantar surface of the hind paw of the rats, and latency was measured in seconds and then recorded as the HWL of each rat to thermal stimulation.
Rat HWLs to nociceptive mechanical stimulation were assessed using the Randall Selitto Test (Ugo Basile, Type 37215). The push rod was applied at a loading rate of 30 g/s onto the dorsal surface of its hind paw, then recorded the latency necessary to induce a withdrawal reaction as HWL in response to mechanical stimulation. Each rat was examined using thermal and mechanical stimuli. To prevent tissue damage due to thermal and mechanical stimulation, a cut-off limit of pain detection was set at 15 seconds.
Before microinjection into the NAc, the HWLs were detected thrice to obtain an average value that was designated as the basal HWLs. The HWLs examined in subsequent experiments were then recorded as the percentage change at the basal level (% changes of HWL) in every rat examined.
5. Surgical procedure and intra-NAc microinjection
A 60 mg/kg intraperitoneal injection of sodium pentobarbital was used to anesthetize the rats. Then, the rats were placed on a brain stereotaxic instrument. A stainless steel guide cannula with an external diameter of 0.8 mm was directed into the bilateral NAc (1.6 mm anterior to Bregma, 1.6 mm lateral to midline, and 7.0 mm ventral to the skull surface) following Paxinos and Watson [21], which was fixed using dental acrylic to the skull. After the surgery, the rats received daily injections of penicillin (80,000 units/one rat) for three days. Three days of recuperation were allowed before the following behavioral experiment was performed.
During the experiment, a syringe needle (0.4 mm external diameter) was inserted into the guide cannula, which was 1.5 mm longer than the cannula tip. Microinjected 0.5 µL of solution into each lateral NAc at a rate of 0.5 µL/min. The syringe needle was left in place for 1 minute after every microinjection.
At the end of the experiment, the rats’ brains were removed, and microinjection sites were confirmed according to the brain atlas. Only the results from rats whose injection sites were within the NAc were included in the statistical analysis.
6. Western blotting assay
CaMK II expression levels were assessed using western blotting, using glyceraldehyde-3-phosphate dehydrogenase (GAPDH) as a reference. Upon completion of the experiment, the rats were anesthetized using 4% isoflurane, and the brains were immediately collected. The NAc was dissected on ice and they were flash frozen at –80°C. The NAc tissue samples were then homogenized in RIPA lysis buffer (Beyotime Institute of Biotechnology) on ice. After centrifugation at 14,000 rpm for 10 minutes at 4°C, the supernatant was collected. The protein concentrations were then measured using a BCA Protein Assay Kit (Beyotime Institute of Biotechnology). An equal amount of 20 µg of total protein was loaded into each lane, which were then separated by 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis. Later, the proteins were transferred onto PVDF membranes (MilliporeSigma), then blocked with 5% nonfat dry milk in Tris-buffered saline Tween (TBST) for 1 hour at room temperature, followed by incubation in blocking solution containing primary antibody (rabbit anti-CaMK II antibody 1:5,000; Abcam, rabbit anti-CaMK II (phospho T286) antibody 1:1,000; Abcam); or horse radish peroxidase (HRP)-conjugated GAPDH (1:10,000; Kangcheng) overnight at 4°C. The membranes were later washed with TBST thrice, with 10 minutes for each wash and then incubated for 1 hour at room temperature with an HRP-conjugated anti-rabbit secondary antibody (1:5,000; ZhongShan GoldenBridge Technology) or TBST. After another batch of 3 × 10 minutes washes using TBST, the bands were visualized using enhanced chemiluminescence reagents (Beyotime Institute of Biotechnology). The relative band densities were evaluated using ImageJ. The ratios of the optical densities of total CaMK II (t-CaMK II) or phosphorylated CaMK II (p-CaMK II) relative to those of GAPDH were determined for each sample.
7. Statistical analysis
The data collected from the experiment were expressed as mean ± standard error of the mean and analyzed with GraphPad Prism 5. Differences between groups were evaluated by two-way ANOVA for repeated measures or the Student’s t-test (two-tailed) where applicable. Differences with P < 0.05 were deemed statistically significant.
Go to :
RESULTS
1. Antinociceptive effects caused by intra-NAc microinjection of AIP into naïve rats
To examine the role of CaMK II in pain regulation in the NAc, four groups of naïve rats were subjected to intra-NAc microinjection of 4 µg (n = 8), 8 µg (n = 9) or 12 µg (n = 7) of AIP, with 1 µL of 0.9% sterilized saline as a control (n = 10). Relative to the control group, HWLs in response to noxious thermal and mechanical stimuli in the rat group treated with AIP significantly increased followed intra-NAc administration of 4 µg of AIP (hot-plate test: Fleft/left = 194.60, P < 0.001; Fright/right = 188.50, P < 0.001; Randall Selitto test: Fleft/left = 264.40, P < 0.001; Fright/right = 550.70, P < 0.001), 8 µg of AIP (hot-plate test: Fleft/left = 194.70, P < 0.001; Fright/right = 271.10, P < 0.001; Randall Selitto test: Fleft/left = 296.60, P < 0.001; Fright/right = 775.20, P < 0.001), or 12 µg of AIP (hot-plate test: Fleft/left = 329.40, P < 0.001; Fright/right = 484.80, P < 0.001; Randall Selitto test: Fleft/left = 659.20, P < 0.001; Fright/right = 984.00, P < 0.001), as shown in Fig. 1. These results implied that intra-NAc microinjection with CaMK II inhibitor AIP imparted significant antinociceptive effects in naïve rats.
 | Fig. 1Antinociceptive effect of microinjection of 4, 8 or 12 µg of AIP into NAc in naïve rats. The left HWL are shown in A (noxious thermal stimulation) and C (noxious mechanical stimulation); the right HWLs are depicted in B (noxious thermal stimulation) and D (noxious mechanical stimulation). The data in the experiment are expressed as mean ± standard error of the mean. Two-way ANOVA was employed to analyze the statistical difference among experimental groups. ***P < 0.001 means relative to the control group. AIP: autocamtide-2-related inhibitory peptide, NAc: nucleus accumbens, HWL: hindpaw withdrawal latency.
|
2. Effect of AIP intra-NAc administration on naïve rat CaMK II expression
To study the effect of AIP on CaMK II expression, rats received an intra-NAc microinjection of 12 µg of AIP (n = 3) or 1 µL of 0.9% sterilized saline as a control (n = 3). CaMK II expression and activity were assessed based on the degree of autophosphorylation at Thr 286 using western blotting. P-CaMK II/t-CaMK II ratio (n = 3; t = 3.57; P = 0.023) significantly decreased after intra-NAc microinjection of AIP (Fig. 2). The results show that p-CaMK II is possibly involved in pain regulation in the NAc, inhibiting p-CaMK II by AIP-induced antinociception in naïve rats.
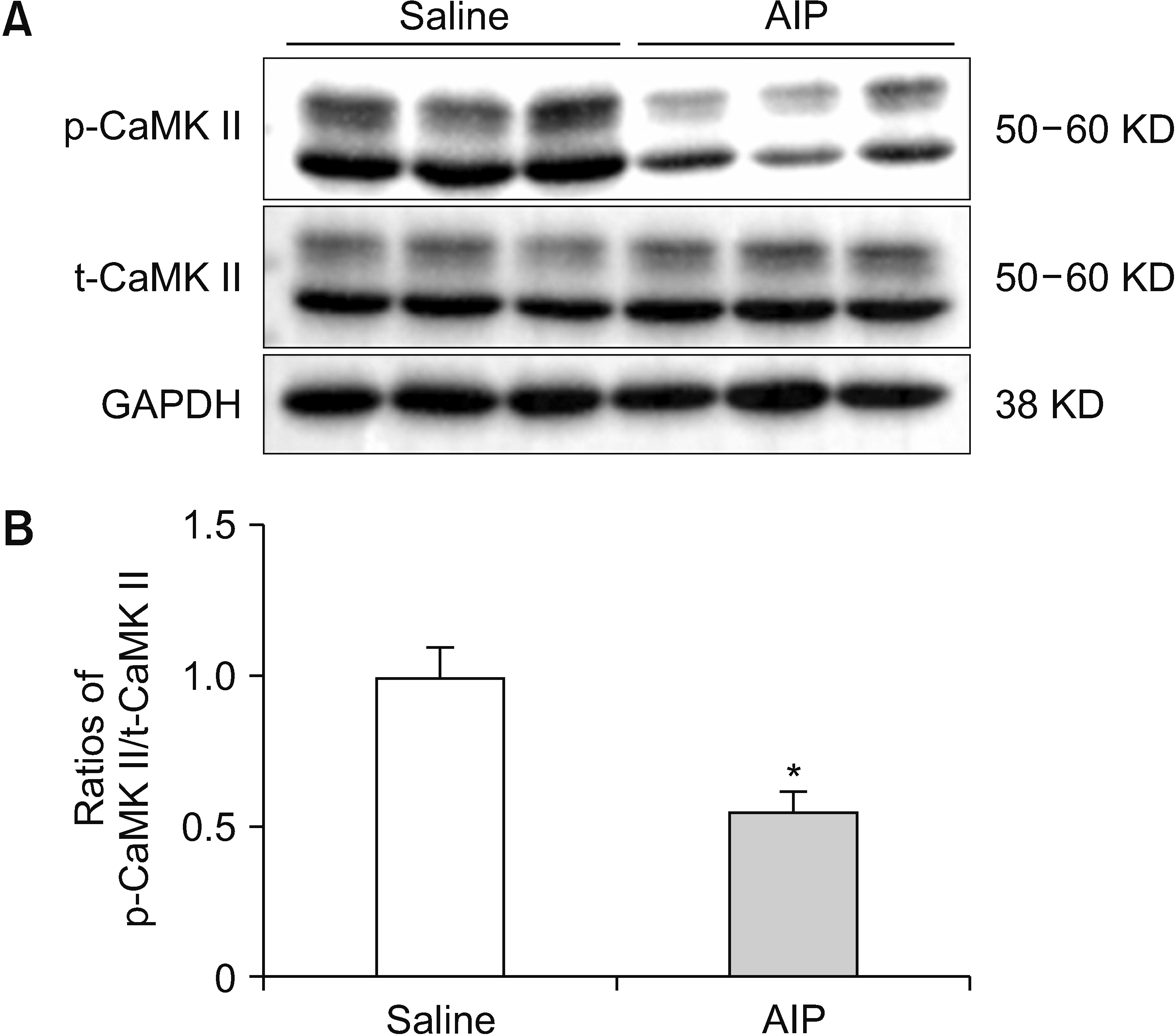 | Fig. 2Changes in CaMK II expression after AIP intra-NAc administration. (A) p-CaMK II/t-CaMK II and GAPDH western blotting results of the saline control and AIP groups. (B) p-CaMK II/t-CaMK II ratios are displayed using histograms. The data in the experiment are expressed as the mean ± standard error of the mean. Two-tailed Student’s t-test was employed to assess the statistical difference among groups. Differences with *P < 0.05 are deemed statistically significant. CaMK II: calcium/calmodulin-dependent protein kinase, AIP: autocamtide-2-related inhibitory peptide, NAc: nucleus accumbens, p-CaMK II: phosphorylated CaMK II, t-CaMK II: total CaMK II, GAPDH: glyceraldehyde-3-phosphate dehydrogenase.
|
3. Influence of morphine tolerance on CaMK II expression in the NAc of rats
To elicit chronic tolerance to morphine antinociception, the rats (n = 10) intraperitoneally received morphine (10 mg/kg) twice a day for a total of 7 days. As shown in Fig. 3, the antinociceptive effects caused by morphine at 30 minutes after the first injection significantly decreased, and the rats exhibited morphine tolerance on day 7, relative to that of the first day (hot-plate test: tleft = 7.03, P < 0.001; tright = 4.82, P < 0.001; Randall Selitto test: tleft = 11.93, P < 0.001; tright = 10.28, P < 0.001). The authors further explored the influence of chronic morphine tolerance on CaMK II expression. CaMK II expression and activity in the NAc were tested by analyzing the degree of autophosphorylation at Thr 286 using western blotting. Rats (n = 3) received morphine injections intraperitoneally twice a day for seven days for induction of chronic morphine tolerance. For the control group (n = 3), rats received the same volume of 0.9% sterilized saline. Fig. 4 shows that compared with the saline group, repeated administration of morphine resulted in a significant increase of p-CaMK II/t-CaMK II (n = 3; t = –5.69; P = 0.005) in the NAc. The results indicated that morphine tolerance increased the p-CaMK II/t-CaMK II ratio significantly in the rat NAc.
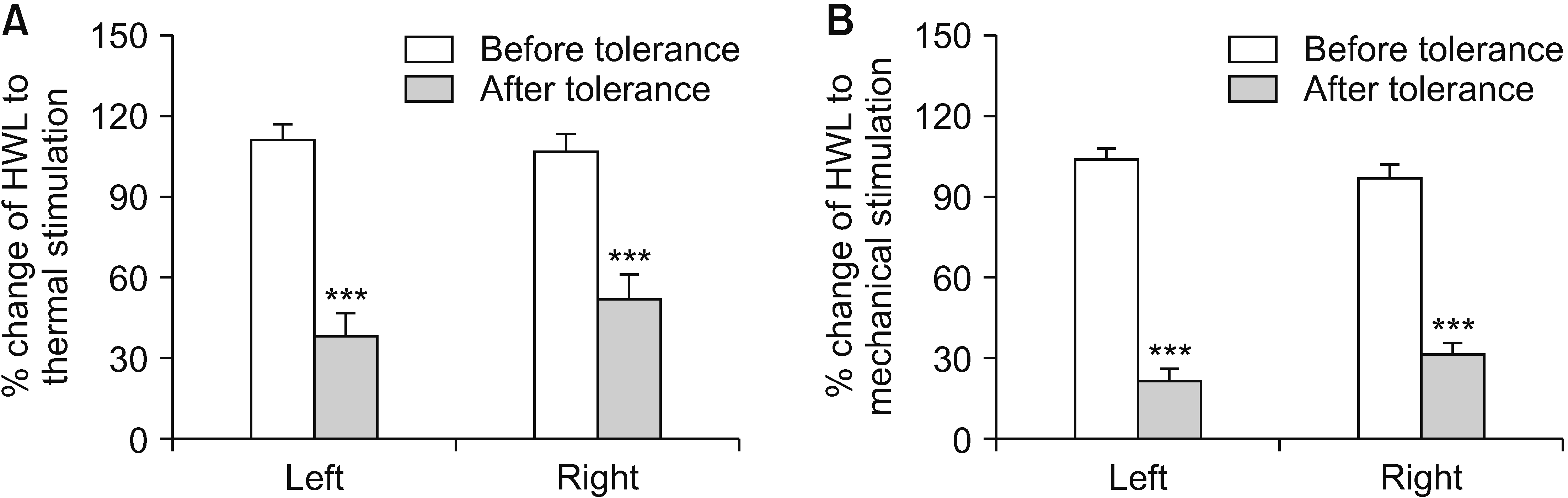 | Fig. 3Morphine tolerance induced by intraperitoneal morphine injection. A indicates HWL in response to noxious thermal stimulation; B represents HWLs to noxious mechanical stimulation. Rats have received intraperitoneal morphine injection for 7 days, and the antinociceptive effects are evaluate 30 minutes after the first morphine administration on day 1 and 7. The data of the experiment are expressed as mean ± standard error of the mean. Two-tailed Student’s t-test was employed to assess the statistical difference among groups. Differences with ***P < 0.001 are deemed statistically significant. HWL: hindpaw withdrawal latency.
|
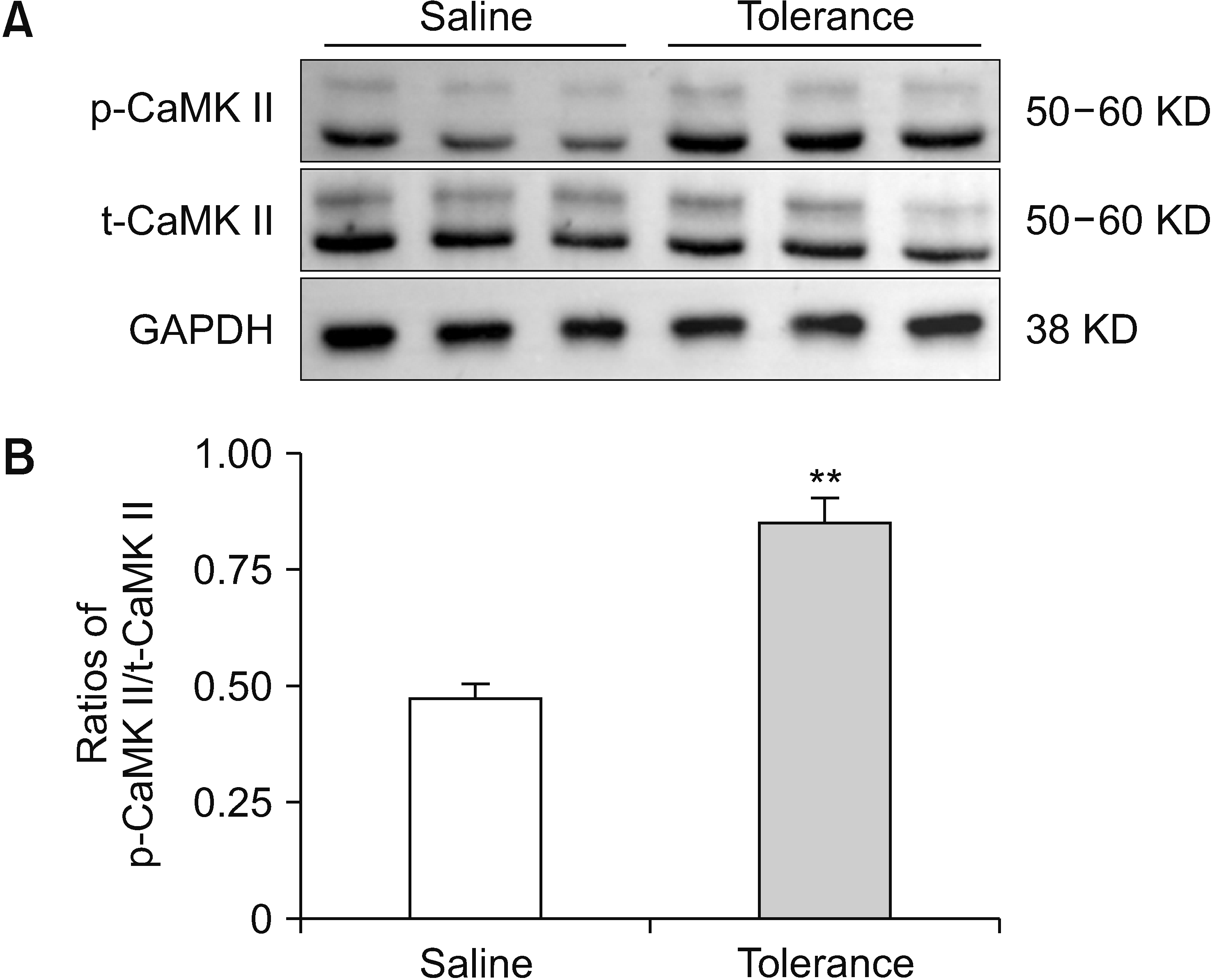 | Fig. 4Changes in CaMK II expression in morphine-tolerant rat NAc. (A) p-CaMK II/t-CaMK II and GAPDH western blotting results of the saline control and morphine tolerance groups. (B) p-CaMK II/t-CaMK II ratios are shown in histograms. The data of the experiment are expressed as mean ± standard error of the mean. Two-tailed Student’s t-test was employed to analyze statistical differences among groups. Differences with **P < 0.01 are deemed statistically significant. CaMK II: calcium/calmodulin-dependent protein kinase, NAc: nucleus accumbens, p-CaMK II: phosphorylated CaMK II, t-CaMK II: total CaMK II, GAPDH: glyceraldehyde-3-phosphate dehydrogenase.
|
4. Antinociceptive effects caused by AIP intra-NAc microinjection into morphine-tolerant rats
The above results revealed the involvement of CaMK II in rat NAc chronic morphine tolerance. Thus, experiments were conducted to correlate the biochemical changes with the behavioral effects. Four groups of rats exhibiting chronic morphine tolerance received intra-NAc microinjections, i.e., they received (1) 1 µL of 0.9% sterilized saline as a control (n = 10), (2) 4 µg of AIP (n = 9), (3) 8 µg of AIP (n = 10), or (4) 12 µg of AIP (n = 10). Ten minutes later, all of the rats were given an intraperitoneal injection of 10 mg/kg of morphine. Relative to the control (saline plus morphine), the HWLs in response to noxious thermal and mechanical stimuli of the group treated with AIP plus morphine significantly increased after intra-NAc administration of 4 µg of AIP (hot-plate test: Fleft/left = 21.74, P < 0.001; Fright/right = 77.98, P < 0.001; Randall Selitto test: Fleft/left = 23.37, P < 0.001; Fright/right = 43.57, P < 0.001), 8 µg of AIP (hot-plate test: Fleft/left = 89.75, P < 0.001; Fright/right = 75.96, P < 0.001; Randall Selitto test: Fleft/left = 39.24, P < 0.001; Fright/right = 61.15, P < 0.001), or 12 µg of AIP (hot-plate test: Fleft/left = 105.60, P < 0.001; Fright/right = 109.90, P < 0.001; Randall Selitto test: Fleft/left = 104.30, P < 0.001; Fright/right = 146.00, P < 0.001), as shown in Fig. 5. These findings show that intra-NAc administration of AIP imparted significant antinociceptive effects in morphine-tolerant rats.
 | Fig. 5Antinociceptive effect of microinjection of 4, 8 or 12 µg of AIP into NAc in rats showing morphine tolerance. The left HWL are depicted in A (noxious thermal stimulation) and C (noxious mechanical stimulation); the right HWL are shown in B (noxious thermal stimulation) and D (noxious mechanical stimulation). 4, 8 or 12 µg of AIP was injected into NAc 10 minutes prior to intraperitoneal injection of 10 mg/kg morphine. Time = 0 min, indicates the start of intraperitoneal morphine injection. The data of the experiment are expressed as mean ± standard error of the mean. Two-way ANOVA was employed to analyze statistical differences among groups. ***P < 0.001 means relative to the control group. AIP: autocamtide-2-related inhibitory peptide, HWL: hindpaw withdrawal latency, NAc: nucleus accumbens.
|
5. Comparison of the AIP induced antinociceptive effect in the NAc of naïve and morphine-tolerant rats
The above results showed that AIP induced antinociceptive effects both in naïve and morphine-tolerant rats. So, are there some changes in the antinociceptive effect of AIP in these two different states? In order to compare the changes of antinociceptive effect induced by AIP under normal and morphine tolerance conditions, Fig. 6 was used to depict the alteration in antinociception caused by AIP microinjection into the NAc of rats with morphine tolerance, relative to that in naïve rats. These results showed a significant increase in HWLs in response to noxious thermal stimulation post administration of 4 µg of AIP (hot-plate test: tleft/left = –1.60, P = 0.132; tright/right = –2.35, P = 0.033), 8 µg of AIP (hot-plate test: tleft/left = –2.96, P = 0.009; tright/right = –3.81, P = 0.001), but not using 12 µg of AIP (hot-plate test: tleft/left = –1.38, P = 0.187; tright/right = –0.05, P = 0.960) into the NAc of morphine-tolerant rats compared with naïve rats. In addition, no significant difference in HWL to noxious mechanical stimulation was observed after microinjection of 4 µg of AIP (Randall Selitto test: tleft/left = 0.02, P = 0.988; tright/right = 0.08, P = 0.940), 8 µg of AIP (Randall Selitto test: tleft/left = 0.66, P = 0.521; tright/right = –0.55, P = 0.591) or 12 µg of AIP (Randall Selitto test: tleft/left = 0.34, P = 0.738; tright/right = –0.60, P = 0.560) into the NAc. These results indicate that the antinociceptive effects of AIP were not attenuated and may induce a stronger thermal antinociception after establishment of chronic morphine tolerance.
 | Fig. 6Comparison of the antinociceptive effects imparted by intra-NAc microinjection of 4, 8 or 12 µg of AIP on the HWLs of naïve rats and rats showing morphine tolerance. A and B depict HWLs to noxious thermal stimulation; C and D represent HWLs to noxious mechanical stimulation. Left HWLs are presented in A and C; right HWLs are depicted in B and D. The antinociceptive effects are evaluate 30 minutes after drug injection. The data of the experiment are expressed as the mean ± standard error of the mean. Two-tailed student’s t-test was used to assess statistical differences among experimental groups. *P < 0.05, **P < 0.01 indicate compared with the control group. 4 A, 8 A or 12 A represent intra-NAc microinjection of 4, 8 or 12 µg of AIP to naïve rats; 4 A, 8 A or 12 A + morphine represent intra-NAc microinjection of 4, 8 or 12 µg of AIP to morphine-tolerant rats. AIP: autocamtide-2-related inhibitory peptide, HWL: hindpaw withdrawal latency, NAc: nucleus accumbens.
|
Go to :
DISCUSSION
The results of this study showed that the naïve rat HWLs, in response to both thermal and mechanical stimuli, significantly increased after intra-NAc microinjection of AIP. These findings imply that AIP imparts an antinociceptive effect within the NAc. To further detect the antinociceptive effect of AIP in chronic morphine tolerance, the authors also observed that microinjection of AIP into the NAc resulted in prolonged HWLs in rats with chronic morphine tolerance. Interestingly, the thermal antinociceptive effect caused by AIP was stronger in rats exhibiting morphine tolerance relative to naïve rats using the same dose in the NAc. These results suggest that CaMK II participates in the modulation of nociception in naïve rats as well as in chronic morphine-induced antinociceptive tolerance in the NAc. Therefore, targeting CaMK II may not only help in elucidating the mechanism underlying pain modulation, but also provide a potential target of pharmacological intervention for morphine tolerance management.
With the increase in the incidence rate of pain, the widespread use of opioid therapy has proliferated in the past decades [22]. Opioid tolerance is the main reason for decreased pain management and increased doses [23]. In the past 15 years, the death rates caused by opioid overdose in the United States have increased four-fold [24]. For example, over 130 Americans die every day due to opioid overdose in the United States [4]. Therefore, there is an urgent need to develop a new treatment strategy that can both enhance opioid analgesia, as well as alleviate tolerance to improve patient safety [23].
A lot of studies have discussed the involvment of CaMK II in morphine tolerance at the spinal cord level [10,25–28]. Inhibition of spinal CaMK II can disrupt morphine antinociceptive tolerance in rodent models. The acute intrathecally administration of CaMK II inhibitor KN93 attenuated chronic morphine tolerance in rats [25]. Further investigations have demonstrated that chronic morphine treatment results in a significant increase in CaMK II mRNA and protein expression in the spinal cord of mice. Confocal microscopy showed that the increase in expression of CaMK II was mainly localized to the superficial laminae of the dorsal horn [26]. Another study showed that intraperitoneal injection of haloperidol, a butyrophenone antipsychotic drug, inhibited CaMK II; it both prevented and reversed morphine tolerance in mice. Their western blotting experimental results showed that pretreatment or short-term application of haloperidol inhibited the increased CaMK II α activity in the spinal cord [10].
However, few studies have focused on the participation of CaMK II beyond the spinal levels in morphine tolerance [11,12,14]. Fan et al. [14] reported that hippocampal CaMK II may act through learning and memory to affect morphine tolerance and dependence. Intrahippocampal dentate gyrus administration of CaMK II inhibitor in rats prevented the development of morphine tolerance. CaMK II may also affect morphine tolerance independent of learning and memory. Tang et al. [12] reported that intracerebroventricular KN93 injection dose-dependently reversed the established antinociceptive tolerance to morphine. Pretreating mice using KN93 could also prevent the generation of morphine tolerance. Further studies by Hu et al. [11] showed that chronic morphine treatment significantly increased CaMK II expression and activity in the cortices or prefrontal cortex in mice. The present study showed that chronic morphine tolerance induced a significant increase of p-CaMK II in the NAc, and inhibiting CaMK II via intra-NAc administration of AIP in rats resulted in antinociception of established antinociceptive morphine tolerance. These results are concordant to previous findings that have implicated CaMK II in the establishment and maintenance of morphine tolerance.
The µ-opioid receptor (µOR) is considered the most relevant factor influencing opioid tolerance. The desensitization of µOR may play an important role in the development of morphine tolerance [10]. Biochemical evidence supports a direct interaction effect between CaMK II and the µOR [5]. The µOR contains consensus sites for phosphorylation by CaMK II [29]. Constitutively active CaMK II promotes agonist-induced desensitization of the µOR expressed in cellular models, and this effect is abolished by mutation of the putative CaMK II phosphorylation site [5,29,30]. This was further supported by the observation that µOR and CaMK II colocalize to anatomic areas responsible for the transmission and processing of nociceptive signals, including the dorsal root ganglia and the superficial laminae of the spinal cord dorsal horn [31]. However, intracellular Ca2+, CaM, and CaMK II can each be regulated by morphine [5]. Indeed, the expression and activity of CaMK II both increase after morphine exposure [10,11,25,26].
Furthermore, CaMK II may also interact with N-methyl-D-aspartic acid (NMDA) receptors, leading to morphine tolerance [32]. CaMK II and the NMDA receptor may interact in a feedforward manner during the initiation stage of morphine tolerance. Activated CaMK II was phosphorylated, which in turn activated the NMDA receptor, leading to Ca2+ influx through the channels. In addition, increased Ca2+ influx through the activated NMDA receptor results in CaMK II autophosphorylation at position Thr 286 that then activates the kinase [5,11,12].
In conclusion, the results of this study indicate that administration of AIP into the NAc induced antinociceptive effects in naïve rats, and p-CaMK II expression was decreased by microinjection of AIP into the NAc. Moreover, increased p-CaMK II expression in the NAc was observed after chronic morphine tolerance. In addition, the administration of AIP into the NAc imparted antinociceptive effects in morphine-tolerant rats. These results reveal that CaMK II in the NAc participates in the nociception transmission and modulation in naïve and morphine-tolerant rats.
Go to :
 XML Download
XML Download