

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Insulin resistance is the common underlying feature of obesity-related metabolic diseases. A number of mechanisms linking obesity to insulin resistance have been suggested and comprehensively reviewed elsewhere [1-5], and the main cause of insulin resistance is now considered to be chronic low-grade inflammation in obese adipose tissue (AT) [6]. Studies have demonstrated that individuals who are obese and insulin-resistant exhibit elevated levels of proinflammatory cytokines such as tumor necrosis factor β (TNFβ), interleukin 1β (IL-1β), and IL-6 [7]. Proinflammatory cytokines can interfere with insulin signaling via multiple mechanisms, such as serine phosphorylation of insulin receptor substrate 1 (IRS-1) [8], reduction of cytokine signaling 3 (SOCS-3) expression [9], or decrease in transcriptional activity of peroxisome proliferator-activated receptor γ (PPARγ) [10]. However, it is not clear whether AT inflammation is a cause or consequence of insulin resistance [11]. Using an adipocyte-specific insulin-resistant mouse model (mTORC2 knockout mice), Shimobayashi et al. [12] found that insulin resistance in adipocytes precedes and causes AT inflammation by recruiting monocytes and proinflammatory macrophages.
In obese AT, adipocytes undergo hypertrophy and secrete a number of proinflammatory cytokines and chemokines, which activate and attract immune cells. The composition of immune cells that constitute the stromal vascular cell (SVC) fraction of ATs dramatically changes as adiposity increases [13]. In lean AT, eosinophils, type 2 innate lymphoid cells, T helper 2 (Th2) cells, and regulatory T cells are the predominant types of cells [14]. As obesity progresses, the loss of the aforementioned cells is coupled with the infiltration of macrophages, neutrophils, mast cells, T helper 1 cells, and natural killer (NK) cells into AT [14,15]. Adipose tissue macrophages (ATMs) are the most common immune cells in AT. ATMs are conventionally classified as CD11b+ CD11c+ M1 (classically activated) or CD11b+ CD11c− M2 (alternatively activated) macrophages. Classically activated macrophages express proinflammatory genes (e.g., TNFα and inducible nitric oxide synthase) and alternatively activated macrophages express anti-inflammatory cytokines (e.g., IL-10 and transforming growth factor β) [16]. The number and subtypes of ATMs greatly differ depending on the degree of obesity. For example, ATMs comprise less than 10% of all AT cells in lean mice; however, this percentage increases to 50% in extremely obese mice [17]. M2-type macrophages are found in abundance in lean AT while M1-type macrophages are predominant in obese AT. It is now generally accepted that ATMs shift from the M2-to-M1 state during the development of obesity, thereby contributing to chronic low-grade inflammation and insulin resistance [18]. Diverse types of ATMs are also observed between visceral adipose tissue (VAT) and subcutaneous adipose tissue (SAT) or between brown adipose tissue (BAT) and white adipose tissue (WAT) [17,18].
Aging has a similar pathophysiology to obesity, and is also associated with the development of chronic low-grade inflammation within AT [19]. There is strong clinical evidence that the risk of obesity increases with age [20,21]. Thus, obesity is linked not only to aging-associated metabolic diseases but also to aging itself [22,23]. However, the molecular mechanism by which obesity accelerates the aging process, or vice versa, is not well understood. In this review, we discuss the specific effects of sirtuins in the maintenance of AT homeostasis in the context of AT inflammation. Given the emerging therapeutic potential of sirtuin 6 (Sirt6) in metabolic diseases and the advances in the development of small molecule Sirt6 modulators (i.e., activators and inhibitors), we particularly paid attention to Sirt6. The current review presents the suppressive effects of Sirt6 on AT inflammation and its clinical significance.
Go to : 
PLEIOTROPIC NATURE OF SIRTUIN ON OBESITY AND INSULIN RESISTANCE
Sirtuin is a nicotinamide adenine dinucleotide (NAD)+-dependent protein/histone deacetylase that has been highly conserved during evolution from bacteria to mammals [24]. Mammals have seven sirtuins (from Sirt1 to Sirt7) which all possess highly conserved NAD+-binding and catalytic domains, but differ in their physiological functions and subcellular localization [25]. Depending on their isotype, sirtuins can act as a protein/histone deacetylase, a mono-adenosine diphosphate (ADP)-ribosyltransferase, a fatty deacylase, or as a combination of these three (Table 1) [26-41]. Through enzyme dependent- or independent-actions, sirtuins can epigenetically modulate gene transcription and modify protein function via posttranslational regulation. Thus, sirtuins alter the expression level and activity of proteins; mostly enzymes and transcription factors. Since all members of the sirtuin family are ubiquitously expressed in various tissues and play roles in regulating energy metabolism, albeit some more directly than others, we examine their function in detail with a focus on WAT.
Table 1.
Subcellular localization and functions of the mammalian sirtuins
| Sirtuin | Localization | Enzymatic activity | Biological effects | Reference |
|---|---|---|---|---|
| Sirt1 | Nucleus, cytoplasm | Deacetylase | Intracellular energy sensor, adipogenesis, adipose tissue inflammation | [26-29] |
| Sirt2 | Cytoplasm, nucleus | Deacetylase | Adipogenesis | [30] |
| Sirt3 | Mitochondria | Deacetylase | Adaptive thermogenesis | [31] |
| Sirt4 | Mitochondria | ADP-ribosyltransferase | Adipogenesis, lipogenesis | [32,33] |
| Sirt5 | Mitochondria | Deacetylase, demalonlylase, desuccinylase, deglutarylase | Adaptive thermogenesis | [34] |
| Sirt6 | Nucleus | Deacetylase, long chain fatty acyl deacylase, ADP-ribosyltransferase | Lipolysis, adipose tissue inflammation, adaptive thermogenesis | [35-39] |
| Sirt7 | Nucleus | Deacetylase | Adipogenesis, lipogenesis | [40,41] |
![]()
Sirt1
Sirt1, the most extensively studied sirtuin family member, is known for its ability to extend lifespan and for its role in intracellular energy sensor [27]. When mice are starved, Sirt1 in WAT is induced to deacetylate and inactivate PPARγ, thereby repressing the expression of adipogenesis markers [28]. In differentiated adipocytes, Sirt1 promotes the binding of forkhead box protein O1 (FoxO1) and CCAAT enhancer-binding protein alpha (C/EBPα) to the adiponectin gene promoter, and increases the expression of adiponectin, an insulin sensitizing hormone [42]. Transgenic overexpression of Sirt1 or treatment with resveratrol, a polyphenol that activates Sirt1, has been shown to reduce weight gain and metabolic derangements in high-fat diet (HFD)-fed mice [43-46]. Conversely, genetic ablation of Sirt1 in adipocytes leads to increased adiposity and insulin resistance [47]. These metabolic effects of Sirt1 in adipose tissue may be related to its anti-inflammatory properties. Adipocyte-specific Sirt1 ablation recruits macrophages, specifically M1-type macrophages, into adipose tissues in HFD-fed mice [26,48]. Mechanistically, Sirt1 in adipocytes deacetylates nuclear factor of activated T cells 1 (NFATc1) and enhances the binding of NFATc1 to IL-4 promoter, which facilitates M2-type macrophage polarization [48]. In the same study, the authors found that Sirt1 suppresses C-C motif chemokine ligand 2 (CCL2, also known as MCP-1) production in adipocytes, which in turn inhibits the recruitment of macrophages into adipose tissues.
To determine the role of Sirt1 in myeloid cells, Schug et al. [29] generated myeloid cell-specific Sirt1 knockout mice, provided them with a HFD, and observed a high degree of infiltration of macrophages into WAT and systemic insulin resistance. In a previous study, we also detected adipose tissue inflammation and metabolic derangement in myeloid cell-specific Sirt1 knockout mice [49]. When an HFD is consumed, Sirt1 ablation in myeloid cells enhances hyperacetylation of focal adhesion kinase, resulting in increased migration of macrophages into adipose tissue.
Sirt2–5 and Sirt7
Sirt2 is primarily found in the cytoplasm, but it can migrate to the nucleus in a cell cycle-dependent manner [50]. A number of proteins such as α-tubulin [50], FoxO1 [30,51], FoxO3a [52], peroxisome proliferator-activated receptor gamma coactivator 1α (PGC-1α) [53], Par-3 [54], p300 [55], p65 [56], and mitogen activated protein kinase phosphatase-1 [57] have been identified as deacetylation substrates of Sirt2. Studies have shown that Sirt2 expression is elevated in WAT of calorierestricted mice, where it deacetylates FoxO3a and reduces the amount of cellular reactive oxygen species [52]. Sirt2 deacetylates FoxO1 to repress the transcriptional activity of PPARγ, thereby suppressing adipogenesis [30]. Sirt2 also promotes oxidative catabolism in adipocytes by deacetylating PGC-1α, leading to an inverse correlation between adipose Sirt2 expression and obesity in human subjects [53].
Sirt3, Sirt4, and Sirt5 are mitochondrial sirtuins, and their role in obesity and AT inflammation has yet to be elucidated. Sirt3 is highly expressed in BAT and regulates adaptive thermogenesis [31]. Caloric restriction increases Sirt3 expression in both WAT and BAT, where it acts as a critical regulator of lipid homeostasis processes such as fatty acid β-oxidation and electron transport in mitochondria [58]. Global Sirt3 knockout mice display insulin resistance and obesity following suppression of mitochondrial fatty acid oxidation [59]. However, conflicting findings have been reported. In a recent study conducted by Porter et al. [60], adipocyte-specific Sirt3 knockout mice fed an HFD exhibited adipose tissue with normal mitochondrial function and showed no significant changes in whole-body metabolism. This study suggests that Sirt3 in adipocytes does not influence mitochondrial function or cause obesity-induced systemic metabolic complications. Sirt4 has an important role in regulating the proliferation and differentiation of preadipocytes [33]. In contrast to Sirt3, Sirt4 inhibits fatty acid β-oxidation and promotes de novo lipogenesis [32]. Sirt5 is expressed at higher levels in BAT than WAT, and is required for brown adipocyte differentiation. Sirt5 knockout mice exhibit reduced browning capacity in SAT and show cold intolerance [34].
Go to :
REGULATION OF ADIPOSE TISSUE INFLAMMATION BY SIRTUIN 6
Sirt6 is localized in the nucleus and was first identified as an ADP-ribosyltranferase enzyme [61]. Subsequent studies have shown that Sirt6 deacetylates histone H3 at lysine 9 [62], lysine 18 [63], and lysine 56 [64] residues, resulting in a compact chromatin structure. Sirt6 expression is induced in adipose tissue by a calorie-restricted diet or weight loss [35,65], whereas its expression is suppressed in diet-induced obese c57BL/6 mice [35,38], genetically obese db/db mice [39], and overweight humans [66]. Sirt6-deficient mice exhibit severe hypoglycemia, loss of subcutaneous fat, and abnormalities associated with aging, which eventually cause premature death at approximately 4 weeks of age [67]. In contrast, transgenic overexpression of Sirt6 results in less accumulation of triglyceride (TG) in visceral fat, as Sirt6 suppresses the expression of angiopoietin-like 4, which is known to inhibit diglyceride acyltransferase activity [68]. For further information about the role of Sirt6 in adipocytes and adipose-infiltrated immune cells, we refer to recent publications by our and other groups.
Adipocyte Sirt6
Sirt6’s main function in adipocytes is to regulate lipid metabolism and prevent the development of inflammation. When a HFD is consumed, adiponectin (Adipoq)-Cre-mediated Sirt6 deficiency accelerates weight gain due to impaired lipolytic activity [35]. On a molecular level, Sirt6 deficiency increases FoxO1 acetylation, facilitates the nuclear export of FoxO1 to cytoplasm, and reduces its effect on adipose TG lipase induction. In a study that used fatty acid binding protein 4 (Fabp4)-Cre-mediated Sirt6 deficient mice, similar phenotypes were observed. Significant increases in body weight, fat mass, adipose tissue inflammation, and insulin resistance were observed in Sirt6 knockout mice as compared to wild-type control mice [38]. Although the Fabp4-Cre mouse line may be predisposed to a target gene recombination in certain endothelial cells and macrophages, the two studies mentioned above clearly indicate that Sirt6 inhibits maintenance of white adipocytes. Brown adipocytes are also affected by Sirt6. Sirt6 increases the thermogenic function of BAT and the browning of SAT by recruiting phospho-activating transcription factor 2 to the promoter of PGC-1α gene [39]. Our studies using Adipoq-Cre-mediated Sirt6 knockout mice highlight that the pathogenesis of adipose tissue inflammation is affected by Sirt6. These mice displayed increased macrophage infiltration in VAT, mainly of M1-type macrophages, and obvious increases of body weight, fat mass, and systemic insulin resistance, even after being fed a normal chow diet [36]. In the same mouse model, Sirt6 induced the expression of Th2 cytokine IL-4 in adipocytes, which in turn promoted M2-type macrophage polarization (Fig. 1). Combined with our recent study showing that adipocytes require Sirt6 for metabolic adaptation to intermittent fasting [37], these studies demonstrate that Sirt6 stimulates lipolysis, enhances adipose tissue browning, ameliorates adipose tissue inflammation, and thereby improves insulin action in peripheral tissues.
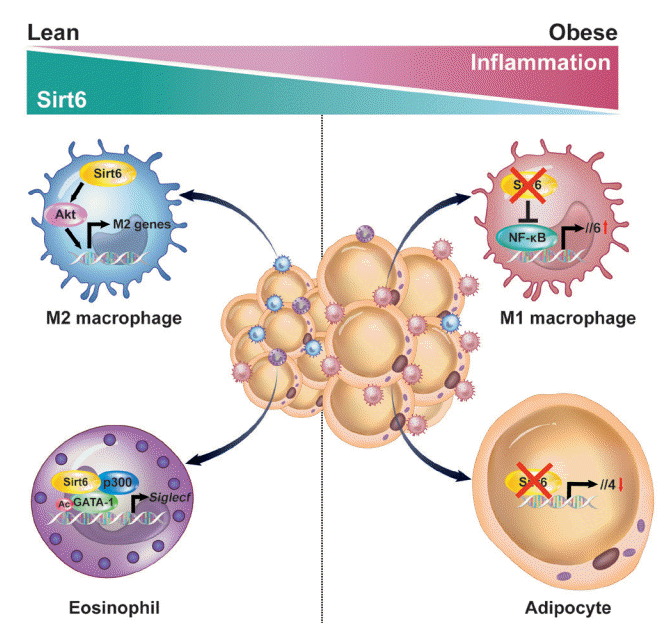 | Fig. 1.Inhibition of adipose tissue inflammation by sirtuin 6 (Sirt6). In adipocytes, Sirt6 increases the production of the type 2 cytokine interleukin 4 (IL-4) to promote M2-type macrophage polarization in a paracrine manner. In eosinophils, Sirt6 forms a ternary complex with GATA binding protein 1 (GATA-1) and p300 acetyltransferase to positively regulate GTAT-1 transcriptional activity. In macrophages, Sirt6 inhibits the nuclear factor-κB (NF-κB)–IL-6–signal transducer and activator of transcription 3 (STAT3) signaling axis and suppresses M1-type macrophage polarization in an autocrine manner. Conversely, Sirt6 activates the phosphoinositide 3-kinases (PI3K)-Akt pathway and promotes M2-type macrophage polarization.
|
Macrophage Sirt6
Hematopoietic stem cells give rise to two different types of blood cells; myeloid lineage cells and lymphoid lineage cells. The former cells include erythrocytes, monocytes, macrophages, neutrophils, eosinophils, basophils, and platelets. The latter cells include lymphocytes and NK cells. Lymphocyte Sirt6 interacts with and deacetylates GATA binding protein 3 (GATA-3), inhibits Th2 immune responses, and suppresses allergic airway inflammation induced by ovalbumin or house dust mice [69]. At the time of writing, no information regarding the role of lymphocyte Sirt6 in adipose tissue inflammation has been reported. However, increased infiltration of lymphocytes has been detected in obese adipose tissues [14,15], warranting future research on the potential role of lymphocyte Sirt6 in adipose tissue inflammation and systemic insulin resistance.
We have investigated the influence of Sirt6 on various inflammatory diseases, including adipose tissue inflammation, with a focus on macrophage polarization. Sirt6 levels are noticeably reduced in VAT after HFD feeding [70]. Myeloid Sirt6 knockout mice exhibit increased proinflammatory M1-type macrophage infiltration in VAT and a concomitant reduction of the M2-type macrophages [70,71]. Mechanistically, a Sirt6 deficiency activates nuclear factor-κB (NF-κB), produces IL-6, and activates the signal transducer and activator of transcription 3 (STAT3), which ultimately polarizes bone marrow cells toward M1-type macrophages (Fig. 1). Similarly, in an experimental rheumatoid arthritis model, myeloid Sirt6 deficiency increased the infiltration of M1-type macrophages in joint tissues [72]. Sirt6 also affects M2-type macrophage polarization in myeloid cells. Under IL-4-stimulated M2 polarization conditions, Sirt6 activates the phosphoinositide 3-kinases (PI3K)-Akt pathway and increases the expression of M2 marker genes [73]. In a full-thickness excisional lesion model, M2 type macrophage infiltration was markedly suppressed in myeloid Sirt6 knockout mice, resulting in prolonged inflammation in the wound site and delayed wound closure.
Eosinophil Sirt6
Eosinophils are also important cells in adipose tissue inflammation. Eosinophils differentiate from CD34+ progenitor cells in bone marrow under the influence of a variety of cytokines. Of these cytokines, IL-5 secreted by type 2 innate lymphoid cell (ILC2) cells is essential to support the growth, activation, and survival of eosinophils [74]. GATA transcription factors are crucial in eosinophil lineage commitment and differentiation [75]. We recently reported that Sirt6 is an essential factor in eosinophil differentiation via cooperation with GATA-1, which, to our knowledge, is the only report describing the role of eosinophil Sirt6 in the biology of adipose tissue [76]. Importantly, as Sirt6 simply acts as a scaffolding protein that recruits p300 acetyltransferase and GATA-1 in eosinophils, its deacetylase activity is not required for the expression of GATA-1 target genes (Fig. 1). Indeed, mice with a myeloid Sirt6 deficiency display reduced M2-type macrophage content in SAT and impairment of SAT beiging after exposure to cold temperatures. These results are consistent with previous studies which found that adaptive thermogenesis requires production of Th2 cytokines, including IL-4 by eosinophils, which leads to M2 macrophage polarization and browning of white adipocytes [77-79].
Go to :
CONCLUSIONS AND FUTURE PERSPECTIVES
Once largely recognized as an anti-aging molecule with the ability to repair DNA, the results of a number of research studies support the notion that Sirt6 prevents adipose tissue inflammation by modifying cellular processes in adipocytes and the surrounding immune cells.
Numerous studies have emphasized the importance of metabolic homeostasis and plasticity in healthy aging. In cases of obesity, a number of innate and adaptive immune cells infiltrate into adipose tissue and cause inflammation. Notably, adipocytes act as key regulatory cells and control adipose tissue inflammation by providing an antigen to immune cells and by secreting a plethora of adipocytokines. Although the precise mechanisms involved in adipose tissue inflammation have not been fully elucidated, it is clear that a cross-talk between adipocytes and immune cells, or among immune cells, determines the immunometabolic phenotype of adipose tissue.
Experimental results accumulated over the course of a decade indicate the importance of Sirt6 as a regulator of adipose tissue inflammation, which involves the secretion of anti-inflammatory adipocytokines and transcriptional regulation of immune cell maturation. Nevertheless, several questions remain to be addressed in the future. First, the functional role of Sirt6 in several innate and adaptive immune cells, other than macrophages, is poorly understood. Second, although it is evident that Sirt6 is present in both adipocytes and SVCs, transcriptional regulatory networks and intracellular signaling pathways that lead to functional interplay among the cells which compose adipose tissue have not been identified. Third, the therapeutic role of Sirt6 specific modulators (activators and inhibitors) in metabolic diseases including obesity and diabetes need to be understood to a better extent, despite the fact that several compounds have already been identified [80,81]. However, Sirt6 is known to promote the development of certain types of cancer [82]. To increase the therapeutic efficacy of Sirt6 modulators and minimize their side effects when treating adipose diseases, adipose tissue-specific drug delivery or gene therapy should be considered. Suitable answers to the above questions will bring us one step closer to a comprehensive treatment for adipose tissue inflammation and insulin resistance.
Go to :
 XML Download
XML Download