

 PDF
PDF Citation
Citation Print
Print



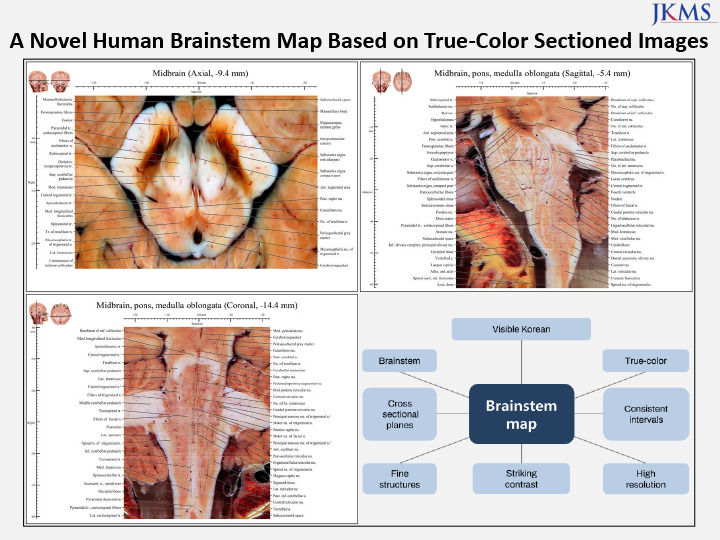
INTRODUCTION
The brainstem is a neural center relaying afferent and efferent information among the cerebrum, cerebellum, and spinal cord.12 As it consists of numerous organized nuclei and fibers that allow neural transmission in narrow and compact spaces, even extremely mild mechanical injury of the brainstem can be fatal.3 Therefore, neurosurgeons and neuroscientists would benefit from an accurate and fine-scale map of the complicated tomography and anatomy of the brainstem. In existing maps of the brainstem, cross-sectional views were used extensively for clinical instruction, scientific research, and basic science teaching. For cross-sectional views, the atlases were generated from magnetic resonance imaging (MRI) images obtained using 7 Tesla scanners45678 and from traditional histologically stained slides.91011
However, given the small, narrow space in which brainstem nuclei and tracts were located, together with the inherent limits of MR scanning and poor color of stained tissues,12 a brainstem atlas comprising images of desirable resolution, color, and field of view was still lacking. Moreover, only a few major planes of the midbrain, pons, and medulla oblongata were shown in the currently available atlases.101314 This has hindered tracing of the entire course of motor and sensory pathways in the brainstem, and the observation of almost nuclei. A further problem in most existing brainstem atlases was that the axial plane was perpendicular to the longitudinal axis of the brainstem,1013 unlike the anterior commissure-posterior commissure (AC-PC) reference system of the cerebrum.1415 Since the axial planes of the brainstem and cerebrum were not parallel to each other, the pathways from the brainstem to the cerebrum, and the nuclei therein, could not be imaged directly and continuously.
Therefore, we present a brainstem atlas in which the true color, shape, and location of the nuclei and tracts are shown at high spatial resolution in sectioned images. From the atlas of this study, nuclei and tracts in the brainstem were identified in detail in axial, coronal, and sagittal sectioned images (48-bit true color, 0.2 mm intervals, pixel size of 0.06 mm × 0.06 mm).16 Our data will be valuable in for clinical, scientific, and neuroanatomical education.
Go to : 
METHODS
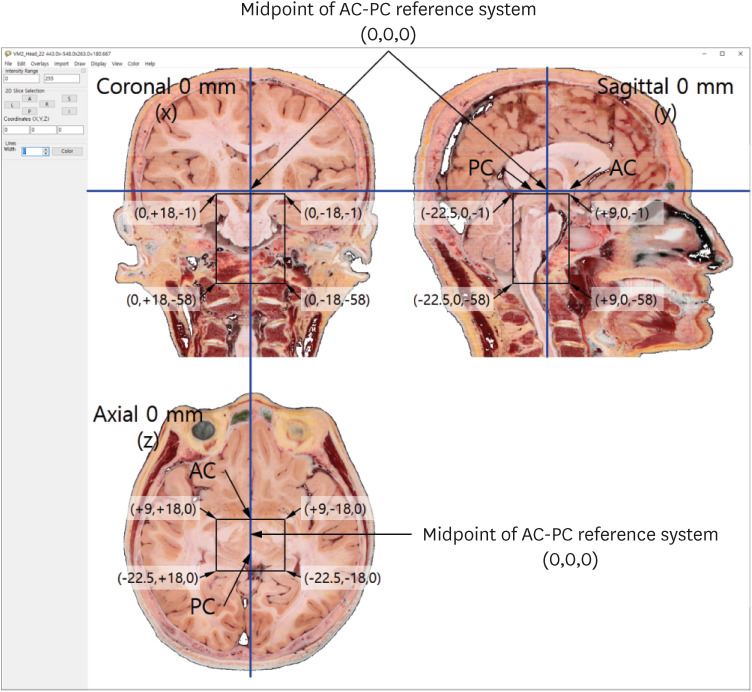
In 2021, we received a 70-year-old male cadaver without brain disease.16 We developed techniques and equipments for making the sectioned images of the cadaver whole body.17 Using the techniques and the equipments, the cadaver was frozen and embedded for a week. The frozen cadaver was milled serially, and the sectioned surface was photographed using high performance digital camera. By repeating the milling and photographing of a cadaver, we produced whole-body axial sectioned images. In the high-quality images, both macro- and micro-structures could be identified.16 The images were obtained following the AC-PC reference system; the axial, coronal, and sagittal planes were accurately defined (Fig. 1).15
From among the axial sectioned images of whole-body, 319 images of the brainstem were selected, extending from superior entrance of the cerebral aqueduct to the first cervical segment of the spinal cord (Fig. 1). In the selected images (resolution, 8,688 × 5,792 pixels), only the brainstem area was retained (resolution, 804 × 641 pixels). The image resolution was decreased by cropping, but the pixel size was unchanged (0.06 mm × 0.06 mm).
The 319 axial sectioned images of the brainstem (resolution, 804 × 641; intervals, 0.2 mm; pixel size, 0.06 mm × 0.06 mm) were imported into the in-house Exslice vol 2.4 software to produce 641 coronal sectioned images (resolution, 804 (width) × 319 (length); interval, 0.06 mm; pixel size, 0.06 mm × 0.2 mm) and 804 sagittal sectioned images (resolution, 641 (width) × 319 (length); 0.06 mm interval; pixel size, 0.06 mm × 0.2 mm). Adobe Photoshop 2021 (Adobe Systems, Inc., San Jose, CA, USA) was used to adjust the resolution of the coronal images to 804 (width) × 1,052 (length × 3.3) (0.06 mm interval; pixel size, 0.06 mm × 0.06 mm) to obtain images with square pixels. Sagittal images (641 × 319) were similarly selected from among horizontal images of the brainstem, with the resolution adjusted to 641 (width) × 1,052 (length × 3.3) (0.06 mm interval; pixel size, 0.06 mm × 0.06 mm) (Fig. 1).
To generate the volume model of whole brainstem, TIFF format sectioned images were converted into DICOM format using Adobe Photoshop 2021; the images were then imported into the DicomBrowser software and a script was applied. The images were then imported into MRIcroGL freeware (https://people.cas.sc.edu/rorden/mricro/) to obtain a brainstem volume model in NIFTI.nii.gz format. The volume model of whole brainstem comprised either three-dimensional (3D) or two-dimensional (2D) axial, coronal and sagittal images, could be viewed using MRIcroGL (Fig. 2).18
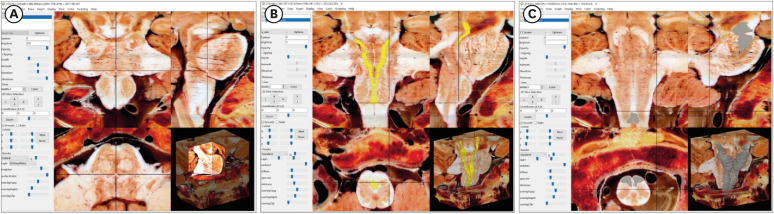
In addition, the volume-of-interest (VOI) tool within MRIcroGL was used to manually segment specific regions and render surface 3D models of structures. The tract was first depicted as a 2D image series, a surface model was then produced and used to precisely track the orientations and tendency of white matter fibers, and to visualize the associated structures in 3D coordinate space (Fig. 2).
The AC and PC were easily and accurately identified in midsagittal sectioned images at 0 mm, and were thus used as the reference system. In the midsagittal image, a plane passing through both the AC and PC, perpendicular to the midsagittal plane, was defined as the 0 mm axial plane.15 A plane passing through the midpoint between the AC and PC, perpendicular to the midsagittal plane, was defined as the 0 mm coronal plane. Based on the AC-PC reference system, sectioned images with dimensions of 18 mm (right-left length) × 31.5 mm (anterior-posterior length) × 57 mm (superior-inferior length) were used to construct the brainstem atlas (Fig. 1).
In the axial sectioned images, nuclei, tracts, and other brainstem structures were identified by reference to photographic10 and stereotaxic19 atlases. The axial, coronal, and sagittal images were incorporated into PowerPoint (2016) slides (Microsoft Corp., Redmond, WA, USA) and 212 structures were annotated (Table 1) according to official anatomy terminology20 to make a brainstem atlas.
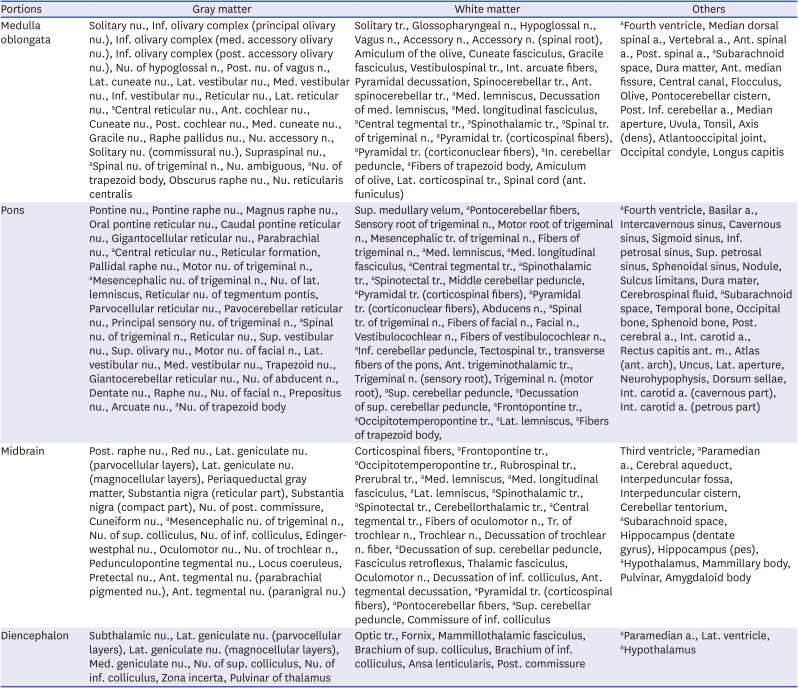
Table 1
212 Annotated structures of brainstem and adjacent portions in a brainstem atlas
a. = artery, ant. = anterior, inf. = inferior, int. = internal, lat. = lateral, m. = muscle, med. = medial, n. = nerve, nu. = nucleus, post. = posterior, sup. = superior, tr. = tract.
aOverlapping structures in medulla oblongata, pons, or midbrain.
![]()
During structure annotation, other planes were referred based on the volume model in MRIcroGL. In addition, by reference to the other 2D planes of the brainstem (Fig. 2), structures could be verified via appearing at the same coordinate of the three planes (axial, coronal, and sagittal),18 thus enhancing the precision and accuracy of the atlas.
Ethics statement
The present study dissecting the donated cadaver was approved by the Institutional Review Board of Wonju Severance Christian Hospital (CR321181).16 The board waived informed consent of the guardians.
Go to :
RESULTS
In 115 axial (0.4 or 0.6 mm interval), 36 coronal (0.9 mm interval), and 20 sagittal (0.9 mm interval) images of the brainstem (pixel size, 0.06 mm × 0.06 mm; color depth, 48-bit; file format, tiff), 212 structures of the midbrain, pons, and medulla oblongata, as well as their junctions, were identified (Table 1). Based on the AC-PC reference system, x, y, and z coordinates in the brainstem’s axial, coronal, and sagittal planes were selected.15 On those planes, the brainstem was located at −3 mm to −60 mm, −3 mm to −38 mm, and 0 mm to 26.1 mm, respectively (Fig. 1). Several structures of interest were included among the 212 brainstem structures, mainly along the axial plane; almost all of them were bilaterally symmetrical in their distribution but were labeled on only one side: white matter on the left and gray matter and other structures on the right (Figs. 3, 4, 5, 6, 7).
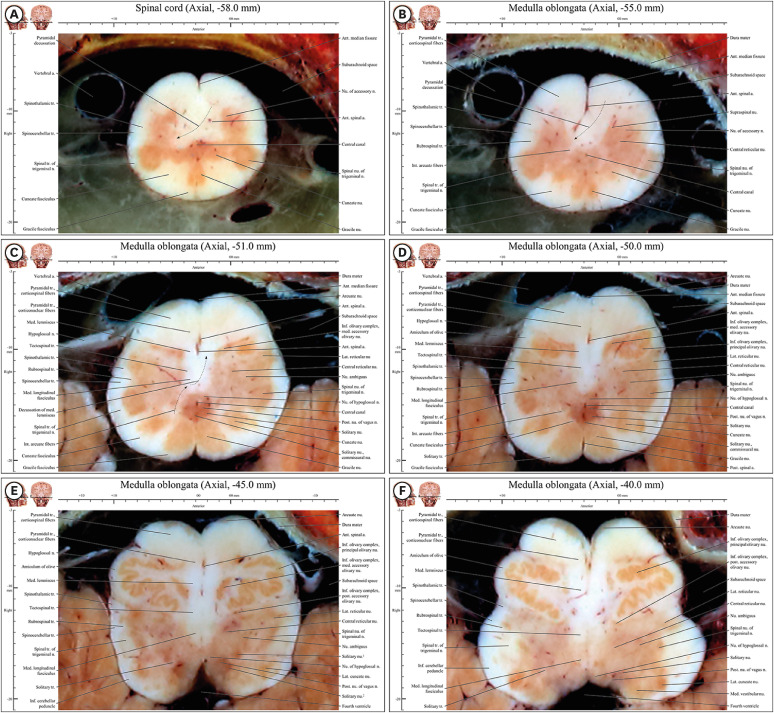 | Fig. 3Axial images of the spinal cord (−58 mm) to the medulla oblongata (−37 mm) in the novel human brainstem atlas. (A) All structures mentioned in the result are noted in underline. Three colonies of gray matter (cuneate nucleus, gracile nucleus, and spinal nucleus of trigeminal nerve) are observed in posterior horn of spinal cord, whereas there is gray matter of a colony in anterior horn of spinal cord. (B) Pyramidal decussation is identified by a curve of anterior spinal artery. In three colonies of gray matter in posterior area, spinal nucleus of trigeminal nerve is still larger than other two nuclei. (C) Three colonies of gray matter in posterior area become similar in size. There are internal arcuate fibers for decussating of medial lemniscus. (D) Inferior olivary complex compose to principal olivary nucleus, medial accessory olivary nucleus, and posterior accessory nucleus. Cuneate and gracile nuclei and tracts exist from (E) caudal end of medulla oblongata to (F) rostral medulla oblongata with inferior olivary complex.
|
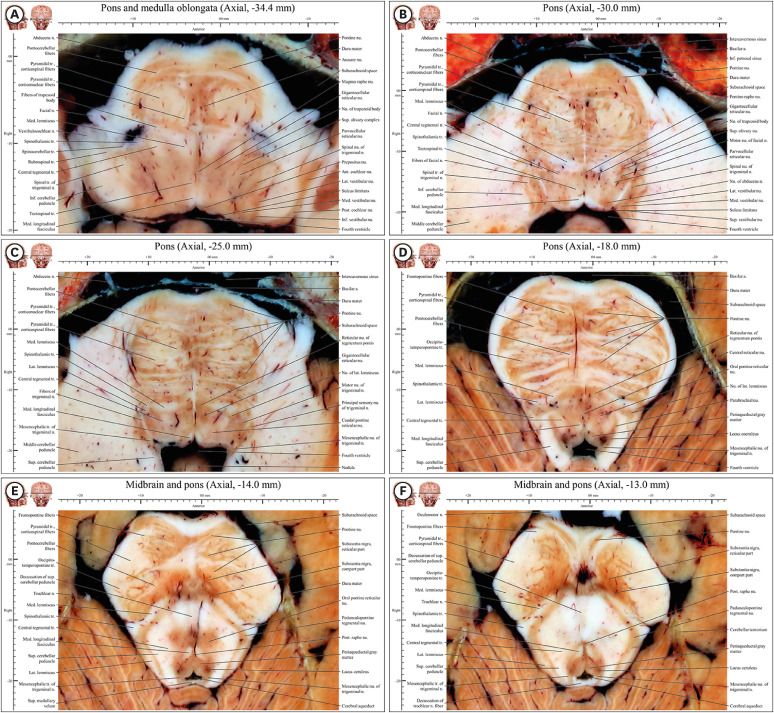 | Fig. 4Axial images of the junction of the rostral medulla oblongata and pons (−36.4 mm) to the junction of the pons and midbrain (−13 mm) in the novel human brainstem atlas. All structures mentioned in the result are noted in underline. In anterior area, pyramidal tract of round shape is located in posterior side of pontocerebellar fibers but the tract is not encircled fully in the fibers unlike them in medulla oblongata. Posterior cochlear nucleus appear in lateral area in (A) and medial vestibular nucleus can be seen continuously from rostral medulla oblongata in (E). Medial lemniscus is moved from medial area to between anterior area and posterior area in (A-F). Sites of nucleus and fibers of facial nerve and nucleus of abducens nerve are lateral area and posterior area, respectively. (B) In posterior area, fibers of facial nerve wrap around behind the nucleus of abducens nerve and then moved forward in. (C) In lateral area, principal sensory nucleus, motor nucleus, and fibers of trigeminal nerve are situated and lateral lemniscus and its nucleus are situated in their lateral side in. (D) In posterior area, locus coeruleus of dark red color is located with lateral side of periaqueductal gray matter. (E) In lateral area, spinotectal tract, spinothalamic tract, and medial lemniscus are connected obliquely. (F) In central area, decussations of trochlear nerve in posterior area and superior cerebellar peduncle are observed.
|
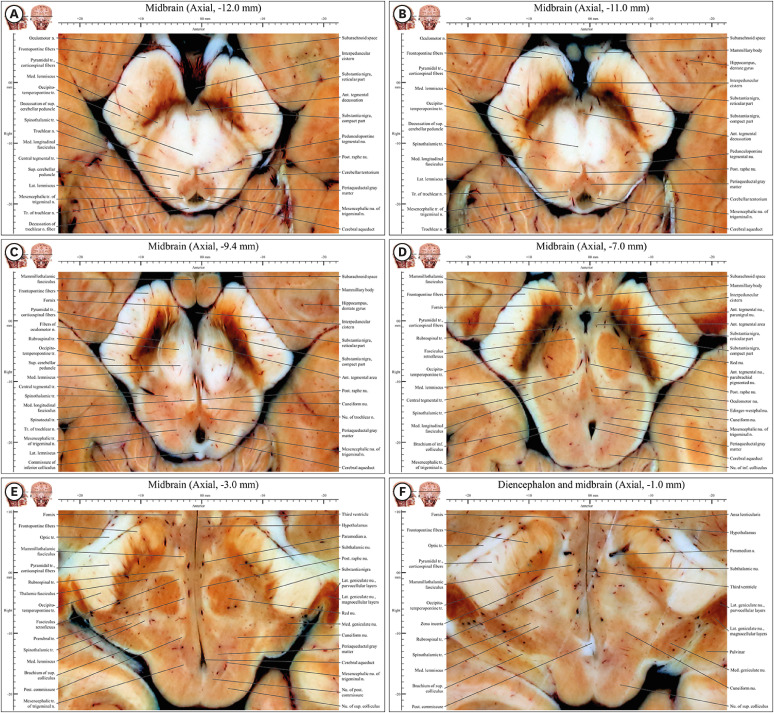 | Fig. 5Axial images of the caudal midbrain (−12 mm) to the rostral midbrain (−1 mm) in the novel human brainstem atlas. All structures mentioned in the result are noted in underline. (A) Substantia nigra is appearing and anterior tegmental nucleus, cerebellar peduncle, and trochlear nerve fibers are decussating. (B) Substantia nigra is divided into in compact and reticular parts. Tract of trochlear nerve is connected with trochlear nerve outside midbrain. (C) Fibers of oculomotor nerve move forward to interpeduncular fossa after the fiber penetrates substantia nigra. Nucleus of trochlear nerve is appearing. (D) Oculomotor nucleus, Edinger-Westphal nucleus, and sensory tracts have well defined features. (E) Subthalamic nucleus and medial and lateral geniculate nuclei are appearing whereas substantia nigra is disappearing. Both diencephalon including lateral geniculate nucleus, medial geniculate nucleus, subthalamic nucleus and midbrain including nucleus of superior colliculus can be seen. (F) This junctional plane has more properties of diencephalon than midbrain. Crus cerebri is clearly divided into frontopontine fibers, pyramidal tract (corticospinal fibers), and occipitotemperopontine tract in entire midbrain.
|
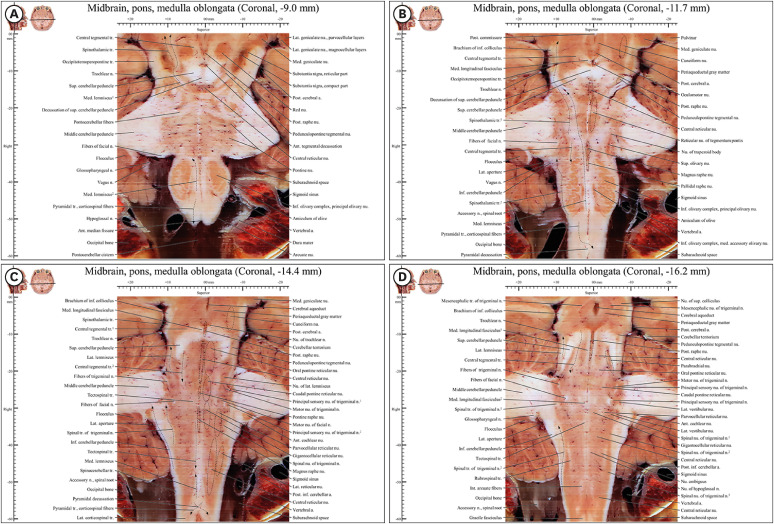 | Fig. 6Coronal images from −9.0 mm to −16 mm in the novel human brainstem atlas. (A) In anterior region (pyramid) of medulla oblongata, there is only pyramidal tract. (B) The medial lemniscus and the spinothalamic tract are located distantly at a distance in medulla oblongata and closely in pons and midbrain because of inferior olivary complex, superior olivary nucleus, and nucleus of trapezoid body. (C) In junction of medulla oblongata and spinal cord, pyramidal decussation can be observed. In middle region, various nuclei of reticular formation can be observed. (D) Posterior raphe nucleus, central reticular nucleus, and parabrachial nucleus are observed.
|
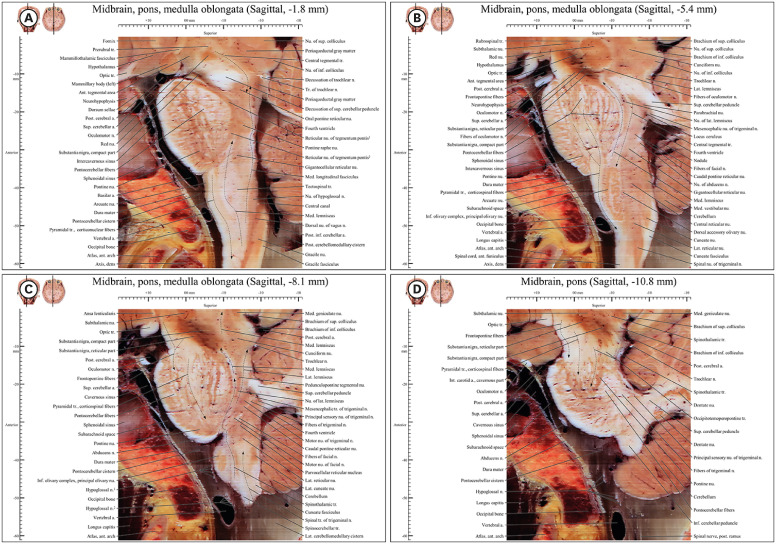 | Fig. 7Sagittal images from −1.8 mm to −10.8 mm in the novel human brainstem atlas. (A) In anterior region (pyramid) of medulla oblongata, there is pyramidal tract in front of medial lemniscus like coronal plane in (Fig. 6A). In posterior region, there is descending fiber of central tegmental tract in (A, B) in front of medial lemniscus. (C) Motor pathway and sensory pathway are located mainly anterior region and posterior region respectively. (D) In pons and midbrain, there are frontopontine fibers, pyramidal tract, and occipitotemoperopontine tract.
|
Spinal cord (axial −58.0 mm)
The anterior, lateral, and posterior area of the central canal of the spinal cord contained the anterior horn (nucleus of the accessory nerve), as well as the anterior funiculus (pyramidal decussation), lateral funiculus (spinothalamic tract, spinocerebellar tract, and spinal tract of trigeminal nerve), and posterior funiculus (cuneate and gracile fasciculus), respectively (Fig. 3A).
The shape of the cervical segment in the spinal cord was not as expected. Three colonies of gray matter (cuneate nucleus, gracile nucleus, and spinal nucleus of the trigeminal nerve) were observed, whereas there were usually a colony of gray matter in the posterior horn of the spinal cord. In this segment of the spinal cord, the pyramidal decussation, gracile fasciculus, and cuneate fasciculus of caudal medulla oblongata were still detected (Fig. 3A).
Junction of the spinal cord and medulla oblongata (axial −57.4 mm and −55.0 mm)
At the border of the foramen magnum, the inferior (axial −58 mm) and superior (axial −57.4 mm) regions of the spinal cord and medulla oblongata, respectively, were identified. The anterior area included the pyramidal tract (corticospinal fibers) and pyramidal decussation, as typical structures of the junction of the spinal cord and medulla oblongata.
The spinal nucleus and trigeminal nerve, spinothalamic, and spinocerebellar tracts were located in the lateral area.
In the posterior area, the cuneate nucleus, cuneate fasciculus, gracile nucleus, and gracile fasciculus were distinctly identifiable (Fig. 3B).
In the central area, right corticospinal fibers in the pyramidal decussation passed from the anterior area of the junction to the lateral area in left side (Fig. 3A). Left fiber in the decussation passed to right area in left side. Movement of fibers to the opposite side corresponded to right or left curves of white matter in planes identified based on a curve for the anterior spinal artery (Fig. 3B).
Caudal medulla oblongata (axial −54.4 mm to −51.0 mm)
In the anterior area, the pyramidal tract was divided into corticospinal and corticonuclear fibers depending on the site. Corticospinal fibers in the lateral and middle parts were light red in color (Figs. 3C, D, 8B, and D). In the medial area, the tract consisted of corticonuclear fibers (Fig. 3C and D).
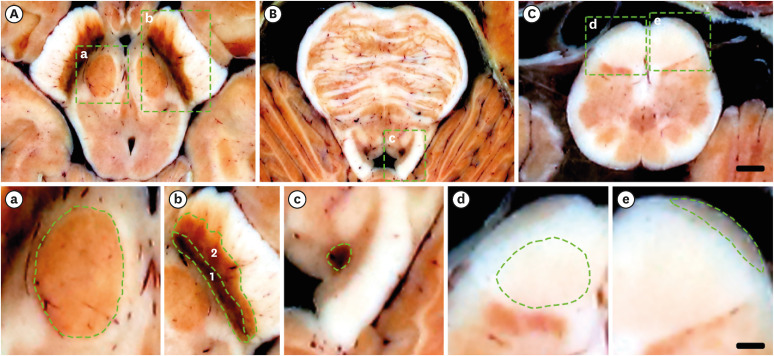 | Fig. 8Specific color of some structures in brainstem. a, b, c, d, e are magnifications of square areas in A, B, C, respectively. (A) It come from the atlas of axial −7.0 mm (Fig. 4D). (B) It come from the atlas of axial −18.0mm (Fig. 3D). (C) It come from the atlas of axial −50.0mm (Fig. 5D). (a) It is the magnifications of red nucleus with red color. (b) It is the magnifications of substantia nigra (compact part) (1) with dark color, and substantia nigra (reticular part) (2) with dark red color. (c) It is the magnifications of locus coeruleus, with dark red color. (d) It is the magnifications of corticospinal fibers with light red color. (e) It is the magnifications of arcuate nucleus with brown color. Scale bar of (A-C): 3 mm; scale bar of (a-e): 1 mm.
|
In the lateral area, the spinothalamic and spinocerebellar tracts, as well as the spinal nucleus of trigeminal nerve, were still observed in both the cervical segment of the spinal cord and the medulla oblongata (Fig. 3B and C).
In the posterior area, both the cuneate and the gracile nucleus and their tracts could still be clearly observed side by side. Cell colonies of the three nuclei (cuneate nucleus, gracile nucleus, and spinal nucleus of trigeminal nerve) were of similar size (Fig. 3C), whereas cell colonies in the spinal nucleus of trigeminal nerve below this region were larger than those of the other nuclei (Fig. 3A).
In the central area, the internal arcuate fibers were those of the decussation of the medial lemniscus.
The medial longitudinal fasciculus could be identified. Around the central canal, the nucleus of hypoglossal nerve, posterior nucleus of vagus nerve, and solitary nucleus appeared side by side. The solitary nuceus was characterized by bilateral nuclei in most regions of the caudal medulla oblongata, but the commissural nucleus of solitary nucleus was seen at the bottom of the latter’s caudal region. In the central area, from posterior to anterior, the commissural nucleus of solitary nucleus, solitary nucleus, posterior nucleus of vagus nerve, and nucleus of hypoglossal nerve were seen. The nucleus ambiguus was also present (Fig. 3C and D).
Rostral medulla oblongata (axial −50.4 mm to −37 mm)
Throughout the anterior area, either the principal olivary nucleus and medial accessory olivary nucleus of inferior olivary complex (Fig. 3D), or the principal olivary nucleus and posterior accessory nucleus of inferior olivary complex (Fig. 3E), occupied the entire area of the rostral medulla oblongata, together with corticospinal and corticonuclear fibers of pyramidal tract.
In the lateral area, the spinothalamic and spinocerebellar tracts, as well as the spinal tract of trigeminal nerve, were still observed from the cervical segment of the spinal cord to the medulla oblongata (Fig. 3E).
In the posterior area, the nucleus of hypoglossal nerve, dorsal nucleus of vagus nerve, and vestibular nucleus were located medially to laterally, contributing to the typical shape of the anterior wall of the fourth ventricle (Fig. 3E).
In the central area, the medial lemniscus and medial longitudinal fasciculus were observed. However, while in most brainstem atlases the cuneate and gracile nuclei and tracts were localized only to the caudal end of the medulla oblongata,1021 we found that these nuclei extended from the caudal end of the medulla oblongata (Fig. 3A) to the rostral medulla oblongata, together with the inferior olivary complex (Fig. 3E). Within this range, the medial lemniscus was “longitudinal” in shape in the anterior to posterior direction. This atypical shape of the rostral medulla oblongata still accommodated the gracile nucleus and cuneate nucleus (Fig. 3A and E).
The reticular nucleus and nucleus ambiguus were between the central and lateral areas (Fig. 3E and F). The solitary nucleus was bilateral in most regions of the rostral medulla oblongata. The commissural nucleus of the solitary nucleus was not present but the solitary nucleus, posterior nucleus of the vagus, and nucleus of the hypoglossal nerve were seen laterally to medially in the central area (Fig. 3D and E), unlike the nuclei of the caudal medulla oblongata.
Junction of the medulla oblongata and pons (axial −36.4 mm to −33 mm)
In the anterior area, corticonuclear and corticospinal fibers of the round-shaped pyramidal tract were located only on the posterior side of the pontine nucleus and the pontocerebellar fibers, as in the medulla oblongata (Fig. 4A), but neither of these fibers was fully encircled by the pontocerebellar fibers, unlike in the pons (Fig. 4B-D). The inferior olivary complex was no longer seen (Fig. 4A).
In the lateral area, spinothalamic tract and spinocerebellar tract, as well as the spinal tract of trigeminal nerve, were still observed (Fig. 4). The lateral and medial vestibular nuclei were observed continuously from the rostral medulla oblongata (Fig. 3F) to the caudal pons (Fig. 4E), whereas the anterior, posterior, superior, and inferior cochlear nuclei were seen only in the pons (axial −36 mm to −31.4 mm) (Fig. 4A and B).
The posterior area contained the medial longitudinal fasciculus and tectospinal tract. The hypoglossal nucleus, together with the prepositus nucleus (motor nucleus) and vestibular nuclei (sensory nucleus), appeared on the medial and lateral sides of the sulcus limitans, respectively, which borders the basal plane (motor nucleus) and alar plate (sensory nucleus) during the embryonic period (Fig. 4A).
In the central area, the medial lemniscus was no longer “longitudinal” in shape (Fig. 3); rather, it was “triangular” (Fig. 4A) and shifted from the medial area, close to the midsagittal line in the medulla oblongata (Fig. 3), toward the lateral area, between the pontine nuclei and tegmentum in the pons (Fig. 4).
In the central area, the pallidal raphe nucleus was present in the same plane as the inferior olivary complex, whereas the magnus raphe nucleus was in the plane without this complex. Because these bilateral nuclei meet in the midsagittal plane, they are referred to as raphe nuclei. On the lateral side of the raphe nucleus are the gigantocellular and parvocellular reticular nuclei (Fig. 4A and B).
Pons (axial −32.4 mm to −15.0 mm)
In the anterior area, the round pyramidal tract was encircled by the pontine nuclei and pontocerebellar fibers across the entire range of the pons (Fig. 4B-D).
The lateral area included the motor nucleus of facial nerve, on the medial side of fibers of facial nerve on the inferior side (Fig. 4B), and the nucleus of lateral lemniscus on the superior side (Fig. 4C). Centered around fibers of the trigeminal nerve were the motor nucleus, principal sensory nucleus of trigeminal nerve, and mesencephalic nucleus of trigeminal nerve; their tracts were on its medial, lateral, and posterior sides, respectively. The mesencephalic nucleus of trigeminal nerve extended from the pons to the midbrain. Between the spinothalamic tract and motor nucleus of trigeminal nerve were the lateral lemniscus and its nucleus (Fig. 4C).
In the posterior area, fibers of facial nerve wrapped around the posterior aspect of the nucleus of abducens nerve and then continued forward (Fig. 4B). The locus coeruleus was dark red in color and located on the lateral side of the periaqueductal gray matter (Fig. 4D, 8A, and C). There were four vestibular nuclei on the lateral side of the fibers of facial nerve. Medial and lateral vestibular nuclei were well separated by white matter (Fig. 4A and B). However, as the border of the superior and inferior vestibular nuclei was not clear, it was defined as the bottom of the abducens nucleus (Fig. 4B), as described in textbooks and existing atlases.101314 Fibers of facial nerve wrapped around the posterior aspect of the nucleus of abducens nerve and then continued forward (Fig. 4B).
In the central area, the medial lemniscus, spinothalamic tract, and lateral lemniscus were situated medially to laterally. Between bilateral medial lemnisci, the reticular nucleus of tegmentum pontis and gigantocellular reticular nucleus were identified (Fig. 4C).
Junction of the pons and midbrain (axial −14.4 mm and −12.4 mm)
In the anterior area, the pyramidal tract shifted from the center of the pontine nuclei and pontocerebellar fibers (Fig. 4A-C) to their lateral side, and entered the crus cerebri of the lateral area in the midbrain. The pontine nuclei and pontocerebellar fibers gradually disappeared and the substantia nigra gradually appeared (Fig. 4E and F).
In the lateral area, the medial lemniscus, spinothalamic tract, spinotectal tract, and lateral lemniscus were connected obliquely (Fig. 4E).
In the posterior area centered around the cerebral aqueduct, the periaqueductal gray matter, locus coeruleus, and mesencephalic nucleus of trigeminal nerve were observed on the anterior, lateral, and posterolateral sides, respectively. In this junction, decussations of trochlear nerve fibers in the posterior area and superior cerebellar peduncle in the central area were first observed (Fig. 4F).
In the central area, the anterior tegmental decussation, decussation of superior cerebellar peduncle, and decussations of trochlear nerve were seen. The medial longitudinal fasciculus was still present (Fig. 4E and F).
Caudal midbrain (axial −12.0 mm to −9.4 mm)
In the anterior area, the pyramidal tract in the crus cerebri could be divided medially to laterally into the frontopontine tract, corticospinal fibers, and occipito-temporo-pontine tract (Fig. 5). Fibers of oculomotor nerve entered through the substantia nigra to become the oculomotor nerve in the interpeduncular fossa (Fig. 5B). The fibers of oculomotor nerve were well seen in the substantia nigra not in the superior cerebellar peduncle because of their white color (Fig. 5C).
In the lateral area, the medial lemniscus, spinothalamic tract, and lateral lemniscus could still be seen (Fig. 5A-C). The lateral lemniscus was connected to the nucleus of inferior colliculus, thereby forming the auditory pathway (Fig. 5C).
In the posterior area, after its decussation (Fig. 5A), the trochlear nerve tract connected with the trochlear nerve outside the midbrain (Fig. 5B). At the upper level of the decussation of trochlear nerve fibers (Fig. 5A), the commissure of inferior colliculus was present (Fig. 5C).
In the central area, the nucleus of trochlear nerve on the anterior side and mesencephalic nucleus of trigeminal nerve on the posterior side centered around the periaqueductal gray matter (Fig. 5C and D).
Four decussations could be identified: the anterior tegmental decussation, decussation of superior cerebellar peduncle in the central area (Fig. 5A and B), decussation of trochlear nerve fibers (Fig. 5A), commissure of inferior colliculus (Fig. 5C) in the posterior area.
Rostral midbrain (axial −9.0 mm to −3 mm)
In front of the interpeduncular fossa, the mammillary body and its connecting fibers (fornix and mammillothalamic fasciculus) were observed (Fig. 5D).
In the anterior area, medial to the substantia nigra, the dark-colored paranigral nucleus of the anterior tegmental area was located between the compact part of substantia nigra and the red nucleus (Fig. 5D). The subthalamic nucleus was located on the upper side of the substantia nigra (Fig. 5E).
In the lateral area, sensory fibers, such as the brachium of inferior colliculus, spinotectal tract, spinothalamic tract, and medial lemniscus, were assembled (Fig. 5D).
In the posterior area, the nucleus of inferior colliculus connected with the medial geniculate nucleus through the brachium of superior colliculus. The lateral geniculate nucleus contained the parvocellular and magnocellular layers (Fig. 5E).
In the central area, the oculomotor nucleus and Edinger-Westphal nucleus were seen around the periaqueductal gray matter (Fig. 5D). The mesencephalic nucleus and tract of trigeminal nerve were located at the lateral boarder of the periaqueductal gray matter (Fig. 5D and E). The medial longitudinal fasciculus from the medulla oblongata was connected to the posterior commissure in the midbrain (Fig. 5D and E).
Junction of the midbrain and diencephalon (axial −2.4 mm to −1 mm)
In the anterior area, the ansa lenticularis and optic tract were located in front of the crus cerebri; the subthalamus was behind the crus cerebri. On the medial side of the crus cerebri, the fornix and mammillothalamic fasciculus were present (Fig. 5F), starting in the mammillary body (Fig. 5D).
In the lateral area, the lateral geniculate nucleus could be divided into parvocellular and magnocellular layers. In the visual pathway, the nucleus of superior colliculus connected to the lateral geniculate nucleus through the brachium of superior colliculus (Fig. 5F). Along the auditory pathway, the medial geniculate nucleus (Fig. 5F) connected to the nucleus of inferior colliculus through the brachium of inferior colliculus (Fig. 5D).
In the central area, the red nucleus disappeared and the zona incerta appeared (Fig. 5F).
Motor and sensory pathways
The motor pathways included the frontopontine fibers, pyramidal tract, and occipito-temporo-pontine tract in the anterior brainstem. The pyramidal tract extended from the precentral gyrus of the cerebrum to the lateral funiculus of the spinal cord, through the internal capsule in cerebrum and whole brainstem. At the junction of the medulla oblongata and spinal cord, the pyramidal decussation was observed (Fig. 6A-C). As the destination of the frontopontine fibers and occipito-temporo-pontine tract is the pons, they were seen only in the anterior region of the midbrain (Figs. 4D-F, 7C, and D).
In the motor pathways, descending fibers of the central tegmental tract extended from the red nucleus (Fig. 6A) to the inferior olivary nucleus (Fig. 6B). When the tract crossed from the midbrain to the pons, its fibers mixed with those of the posteromedial part of the superior cerebellar peduncle (Figs. 6D and 7A)
The spinothalamic tract was identified in the sensory pathway. As soon as the tract entered the posterior horn of the spinal cord, it decussated to the opposite lateral funiculus in the spinal cord. The tract climbed all the way up to the lateral area of the medulla oblongata, medial area of the pons, and lateral area of the midbrain (Fig. 6A and B).
The sensory pathway included the medial lemniscus pathway. In the caudal medulla oblongata, the internal arcuate fibers of the medial lemniscus decussated from the cuneate and gracile nuclei to the medial area of the opposite side, between the anterior median fissure and central canal (Fig. 3C). In the image series used in this atlas, the complete route of the internal arcuate fibers could be easily traced, unlike in other atlases.101314 In the pons, the medial lemniscus and spinothalamic tract met at the top of the nucleus of trapezoid body (Fig. 6B), together passing to the thalamus (Fig. 6A).
Cranial nerves II–XII
Many brainstem structures exert their functions through the cranial nerves. The optic nerve (cranial nerve II) was associated with the optic tract, lateral geniculate nucleus, and nucleus and brachium of superior colliculus (Fig. 5F), and the oculomotor nerve (cranial nerve III) with the oculomotor nucleus and Edinger-Westphal nucleus, both located in the midbrain (Fig. 5D). Regarding the trochlear nerve (cranial nerve IV), associated brainstem structures include its nucleus (Fig. 5D) and the decussation of its nerve fibers (Fig. 5A), while for the trigeminal nerve (cranial nerve V) they include the mesencephalic nucleus and tract in the midbrain (Fig. 5A), principal sensory and motor nuclei in the pons (Fig. 4C), and spinal nucleus and tract in the pons and medulla oblongata (Fig. 3). Regarding the abducens nerve (cranial nerve VI), major structures include its nucleus, located in front of the facial nerve fibers (Fig. 4B), while for the facial nerve (cranial nerve VII) they include the motor nucleus, solitary nucleus, and spinal nucleus and their tracts (Fig. 3A and B). The vestibulocochlear nerve (cranial nerve VIII) is associated with several nuclei of the vestibular and cochlear nerves (Fig. 3A and B). Important brainstem structures associated with the glossopharyngeal nerve (cranial nerve IX) include the nucleus ambiguus (Fig. 3F), solitary nucleus, and spinal nucleus, while for the vagus nerve (cranial nerve X) they include the nucleus ambiguus and posterior nucleus (Fig. 3E). Where the solitary nucleus splits bilaterally, the solitary tract passes between them (Fig. 3E). The nucleus of the accessory nerve (cranial nerve XI) extends from the cervical spinal cord and caudal brainstem (Fig. 3A and B). The nucleus of hypoglossal nerve (cranial nerve X) is located in the medulla oblongata (Fig. 3).
Reticular formation
In the midsagittal area of the brainstem, the posterior raphe nuclei were identified in the midbrain, and the central reticular nucleus, pontine raphe nucleus, magnus raphe nucleus, and pallidal raphe nucleus in the pons. “Raphe” refers to the contact of these structures with both sides of the midsagittal plane (Figs. 6C, D, and 7A).
The reticular formation of the medial area is situated only in the pons. The reticular nucleus of the tegmentum pontis is on the anterior side of the pons. The oral pontine reticular nucleus, caudal pontine reticular nucleus, and gigantocellular reticular nucleus were observed on the posterior side (Fig. 6B and C).
In the lateral area, the cuneiform nucleus is located between the medial lemniscus and superior and inferior colliculi in the midbrain. The parabrachial nucleus and parvocellular reticular nucleus were seen medial to the superior cerebellar peduncle and lateral to the gigantocellular reticular nucleus (Fig. 6C and D).
Go to :
DISCUSSION
In this study, the drawbacks of existing brainstem atlases were resolved by using human sectioned images together with the image processing techniques of the Visible Korean Project.1618 This allowed dynamic changes of the tract bundles and neural nuclei to be tracked in detail using axial, coronal, and sagittal sectioned images.
The first essential factor for imaging brainstem structures is pixel size, which in the sectioned images used in this atlas was 0.06 mm × 0.06 mm.16
The second factor is consistent intervals. In existing brainstem atlases, wide image intervals (> 3 mm) were employed1014 such that structures lying between one image and the next were missed. In this study, axial, coronal, and sagittal sectioned images were obtained at 0.2 mm intervals such that all structures larger than 0.2 mm could be identified. This revealed structures in the anterior-posterior and lateral-medial directions along the axial plane, such as the tract of trochlear nerve (Fig. 5C). In the motor and sensory pathways, this tract travels in the superior and inferior directions. The presence and direction of these structures were confirmed by combining coronal and sagittal images using MRIcroGL (Fig. 2), and then annotating them in an atlas of planar images (Fig. 6 and 7).
Furthermore, in existing brainstem atlases, major planes in the midbrain, pons, and medulla oblongata were defined at irregular intervals,101314 which prevents users from tracing the complete course of the motor and sensory pathways in the brainstem, and omits some nuclei. In particular, existing atlases often failed to show junction regions between the midbrain and pons, pons and medulla oblongata, and medulla oblongata and spinal cord. In this study, sectioned images were obtained at 0.2 mm intervals from the spinal cord to the rostral midbrain, regardless of plane type (major or minor). This approach allowed continuous tracing of the shape and site of each pathway, such as the medial lemniscus, spinothalamic tact of the lateral area in the brainstem (Figs. 3-5) and pyramidal tract (Figs. 6 and 7).
The third factor is color depth. Histologically stained slides910 and micro-computed tomography (micro-CT) of the brainstem are better than sectioned images16 if image quality is based only on pixel size. However, although they may be of microscopic resolution, micro-CT are compromised by 8-bit gray scale (gray, 28). Thus, despite a small pixel size, more than 256 shades of gray will not be displayed and a pixel will not be distinguishable from a boundary pixel. In our study, 48-bit true color sectioned images [color numbers: 216 (red) × 216 (green) and × 216 (yellow)] were used, allowing 281,474,976,710,656 colors to be displayed (Figs. 3-7). Furthermore, to preserve the vivid color of the sectioned images, the cadaver used in our study was not injected with formalin fixative.16 Thus, all of the images in this database reflected the original state and color of the examined structures (Figs. 3-7). For example, in case of locus coeruleus with dark red color, corticospinal fibers with light red color, arcuate nucleus with brown color, red nucleus with red color, compact part of substantia nigra with dark color, reticular part of substantia nigra with dark red color, and so on, we were able to identify diverse colors (Fig. 8).
Fourth, this database used the AC-PC reference system in the cross-sectional plane. To define axial, coronal, and sagittal planes in all cross-sectional images of the brain, including those obtained with MRI and CT, the Talairach and midpoint AC-PC reference systems were established.1415 However, in most brainstem atlases neither system was used,1013 such that the axial planes of the brainstem and cerebrum were not in parallel and the respective structures could not be directly and continuously imaged. Non-parallel axial planes arise in the absence of cross-sectional images continuously showing the brainstem and cerebrum, even if the reference point of the reference system is located in the cerebrum. In case of MRI, small brainstem structures cannot be seen in detail, whereas histologically stained slides do not capture large structures like cerebrum. In the whole-body sectioned images in this study,16 not only the entire outline of a brainstem structure, but also the details of structures between the brainstem and cerebrum, could be observed (Figs. 1-7). The reference point (0 mm) of the AC-PC reference system was accordingly set in the axial image of the cerebrum, and the reference system was then applied in the brainstem (Fig. 1). Furthermore, in the images in this study, structures involved in pathways from the peripheral to the central nervous system, as well as surrounding structures, were continuously visible.16
Structures of the brainstem could be seen in detail along the three planes (Figs. 3-7). However, regardless of image quality, some structures could not be identified in only a plane. We therefore used the 3D volume model in MRIcroGL software, which can display not only stereoscopic, but also sectional views of the coronal, sagittal, and even oblique planes.18 In addition, the VOI tool in MRIcroGL software was employed to highlight structures simultaneously in three 2D planes and 3D coordinate space (Fig. 2).
The medial lemniscus and spinothalamic tract are not in close proximity from each other by the inferior olivary complex, superior olivary nucleus, and nucleus of trapezoid body in the medulla oblongata. From the pons to the thalamus, the two structures are in close proximity when they finally meet at the ventral postero-lateral nucleus in the thalamus (Fig. 6B). At the junction of the medulla oblongata and pons, the shape and size of the medial lemniscus changes from “longitudinal” (Fig. 3F) to “triangular” (Fig. 4A), and from medial (Fig. 3) to lateral (Figs. 4, 5), perhaps due to disappearance of the complex, the nucleus, and the body.
The human brainstem is generally similar to that of paleo-structures, as a primitive controller that sustains life. However, the sectioned images of the brainstem obtained in this study suggest a division into paleo- and archi-structures, because in the medulla oblongata and pons the cortex consists of white matter and the medulla mainly of gray matter (Fig. 3), similar to the spinal cord of the paleo-structure. In the midbrain, most areas consist of gray matter, except the crus cerebri (Fig. 5C-F), also similar to the archi-structure.
In head sectioned images of our previous study, we connected the central nervous system with related structures.2223 In this study, whole structures related to the cranial nerves, except the olfactory nerve, were identified in sectioned images of the brainstem alone. In the full versions of the sectioned images in this study, the whole body could be seen in the axial plane.16 We plan to generate a cross-sectional atlas of the whole body allowing each nerve in the central nervous system to be detected and connected with its target structures.
Neuroanatomists and neuroscientists have long been frustrated by existing brainstem atlases, which mostly consist of cross-sectional images obtained at intermittent and wide intervals.101314 We resolved this problem by creating a brainstem atlas using sectioned images of three planes, and by adhering to four essential conditions (Fig. 1). In brainstem education and research, depiction of each nucleus and tract in neighboring structures, as well the whole brainstem, by our brainstem atlas will be of great value.
Go to :
 XML Download
XML Download