

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Glioblastoma (GBL) is extremely malignant, and it is the most common type of intracranial glioma tumor [1]. While surgery and regulated adjuvant treatment, including simultaneous chemo-radiotherapy using temozolomide, has dramatically improved overall survival (OS) for a GBL, tumor progression remains certain. A leading cause of tumor progression is aggressive infiltration of glioma cells that migrate and invade the surrounding parenchyma. Especially, glioma cells must modify their rigidity and shape to interact with the surrounding microenvironment for migration and invasion. Modification of the rigidity and shape may be related to the rearrangement of the actin cytoskeletons [23].
Cadherins are a kind of surface protein related to cell-cell adhesion. They interact with catenins to regulate adhesion and migration with the surrounding parenchyma [456]. The cadherin-catenin complex plays an important role in regulating the rearrangement of the actin cytoskeleton [45678]. Especially, during tumor progression, cadherin switching is developed as the process by which expression of epithelial E-cadherin is converted to mesenchymal-associated cadherins like N-cadherin and cadherin-11 [910]. Eventually, cadherin-11 plays an important role in tumor invasion and progression, and is highly expressed in multiple aggressive tumors, including breast, prostate, colon, gastric, renal cell, and osteosarcoma [71112]. Recently, the role of cadherin-11 has been demonstrated in the migration of glioma cells in vitro [13]. However, the clinical importance of cadherin-11 in GBL in vivo has not yet been investigated.
The aims of this study were to evaluate progression-free survival (PFS) and OS relative to cadherin-11 expression following a new diagnosis of a primary GBL and to analyze the clinical significance of cadherin-11 expression relative to progression and prognosis.
Go to : 
MATERIALS AND METHODS
Patient population
Approval for this study was granted by the local Institutional Review Board of Kyungpook National University Hospital (KNUH 2021-07-025). The informed consent was waived by the committee. All radiographic and clinical findings were entered in an electronic medical record. From 2007 to 2016, 178 patients at a single institute received a diagnosis of GBL. For 90 out of 178 patients (50.6%), formalin-fixed and paraffin-embedded GBL tissues were checked. Eligible patients were 18 years of age or older. Postoperative MRI was taken within 24 hours of admission, and the extent of resection was analyzed. Gross-total resection was characterized as greater than 95% resection. Following histological confirmation of the GBL, adjuvant standardized chemo-radiotherapy for GBL was given [14]. When there was GBL recurrence, further treatments such as subsequent surgery, salvage chemotherapy, and additional radiation were considered.
A cohort of 38 out of 90 patients was formed according to the following exclusion criteria: secondary GBL with a lower grade glioma (n=15), less than 95% resection (n=14), and additional adjuvant therapies given (n=9). The remaining 52 patients were enrolled in the study. Radiological and clinical assessments were taken at the following times: 28 days after the concurrent-therapy phase; during cycles 2, 4, and 6 of the maintenance phase; every 3 months after the maintenance phase; and at the time of disease progression. Disease progression was confirmed radiologically by MRI following the Response Assessment in Neuro-Oncology (RANO) criteria [15], and survival was ascertained after surgery.
Clinical and radiological variables were as follows: age, sex, Karnofsky performance status (KPS), location, tumor size, and tumor number. Patients were categorized into groups according to age (<65 and ≥65 years) and KPS (<70 and ≥70), respectively. Locations were categorized as “lobar” or “other.” Cases where the tumor was in the corpus callosum or thalamus or where tumors were occipital were included in “other.”
Immunohistochemistry and tissue microarrays of cadherin-11
Cadherin-11 expression intensity was determined with an immunohistochemical study. Each formalin-fixed and paraffin-embedded human primary GBL tissue sample was examined in this study. Paraffin blocks that contained illustrative tumor regions were identified. Tissue microarrays (TMA) paraffin blocks were divided into 4-µm-thick slices and then attached to slides. Every slide cut from TMA blocks was deparaffinized and passed through a moisturizing process. They were then washed with distilled water. Immunohistochemical staining for the cadherin-11 antigen was done with an automated immunostainer (Ventana BenchMark XT, Ventana Medical Systems, Tucson, AZ, USA). The ultraView kit was used in accordance with manufacturer recommendations. Subsequently, a mouse monoclonal cadherin-11 antibody (IgG2B Clone #283416 Catalog Number: MAB1790 [R&D Systems; Minneapolis, MN, USA]) was used as the primary antibody. Then, each primary antibody was incubated at 37℃ for 30 min, and subsequently followed consecutively by normalized Ventana signal amplification and hematoxylin and bluing reagent counterstaining. Following the autostainer procedure, the slides were mounted and analyzed using light microscopy. Negative controls were employed on each tissue section through the replacement of primary antibodies with diluted isotype immunoglobulin (ImmunoCruz™ Staining system, Santa Cruz Biotechnology; Santa Cruz, CA, USA). Subsequently, the slides were incubated with streptavidinHRP and goat antipolyvalent (both Lab Vision; Stockholm, Sweden) for 60 min, then followed by incubation with 3-amino-9-ethylcarbazole. Slides were subsequently washed in PBS, counterstained with hematoxylin for 5 sec, and coverslipped [16].
Evaluation and interpretation of immunohistochemical staining of cadherin-11
Cadherin-11 was mainly immunostained with a cytoplasmic pattern. In normal brain tissue, cadherin-11 is moderately expressed in the cytoplasm of vascular endothelial cells. Although most tumor cells in our study showed stained cadherin-11, the intensity of cadherin-11 staining varied in each case. Cadherin-11 expression was assessed according to staining intensity for endothelial cells, scored from 1 to 3 as follows: 1, staining intensity lower than in endothelial cells; 2, staining intensity similar to that in endothelial cells; and 3, staining intensity higher than in endothelial cells. Illustrative examples of immunostaining are shown in Fig. 1.
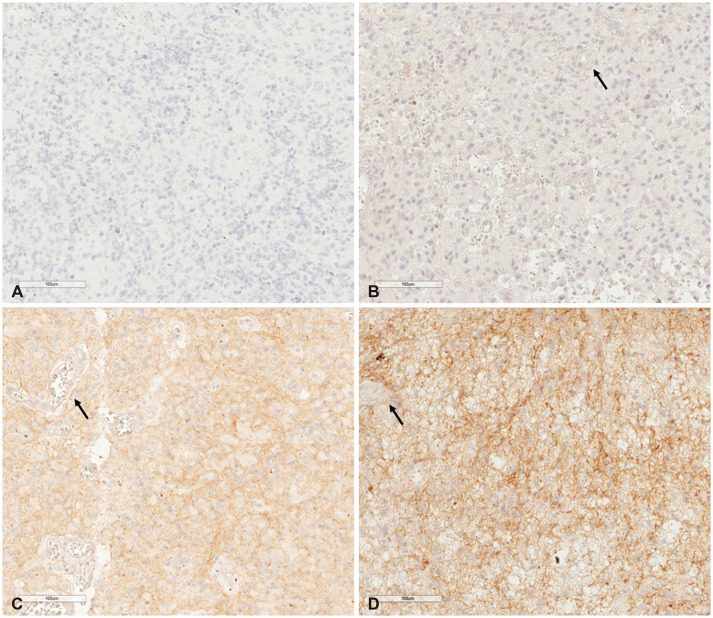 | Fig. 1Cadherin-11 immunohistochemical staining (original magnification ×200; scale bar=100 µm). A: Negative staining of tumor cells. B: Less staining intensity than endothelial cells (arrow). C: Similar staining intensity to endothelial cells (arrow). D: Greater staining intensity than endothelial cells (arrow).
|
Two pathologists (H.W.L. and T.I.P) interpreted immunohistochemical results independently without access to clinical or histological information. All 52 GBLs were positive for cadherin-11 in the staining. Regarding statistical analysis, immunohistochemical staining scores were categorized into two groups: a low (score 1 to 2) intensity group and a high (score 3) intensity group.
Statistical analysis
Comparisons between high-intensity and low-intensity groups were done with chi-square and Fisher exact tests. Survival curves were calculated with the Kaplan-Meier method, and comparisons of survival curves were performed using the log rank test. The Cox proportional hazards model was used for univariate and multivariate analyzes to examine the effect of cadherin-11 expression on PFS and OS. A p-value <0.05 was considered the threshold for statistical significance. Statistical analysis was performed using SPSS 14.0 for Windows (SPSS Inc., Chicago, IL, USA).
Go to :
RESULTS
Patient characteristics
Detailed characteristics of the 52 patients enrolled in this study are given in Table 1. The mean age at initial diagnosis was 56.9±13.3 years, with a proportion of ages <65 years and ≥65 years at 65.4% and 34.6%, respectively. According to sex, there were 25 males and 27 females. The mean preoperative KPS was 75.2±18.5, while the proportion of KPS at <70 and ≥70 was 19.2% and 80.8%, respectively. The most frequent tumor site was the frontal lobe, followed by the temporal, “other”, and parietal lobes. And, the mean maximal diameter of tumors was 4.6±1.1 cm. There were 4 patients with multiple tumors. However, there were statistically significant differences between the high-intensity and low-intensity cadherin-11 groups in terms of sex (p=0.013) and preoperative KPS (p=0.047).
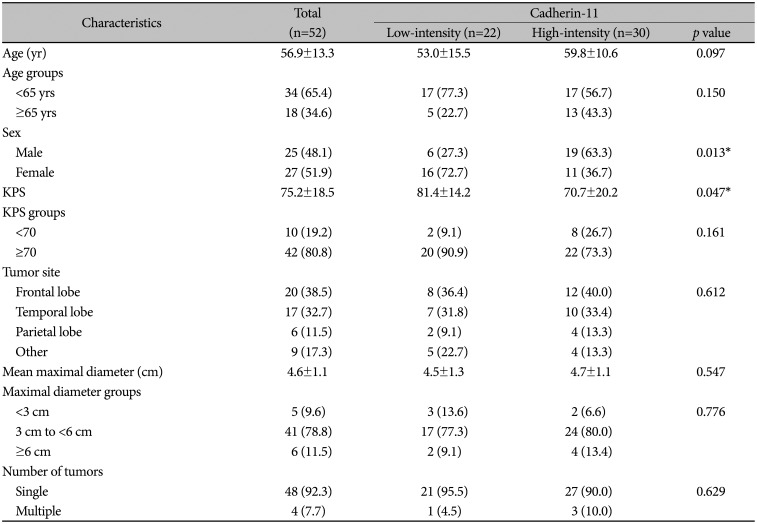
Table 1
Characteristics of 52 patients with glioblastoma between high-expression and low-expression of cadherin-11 groups
![]()
Cadherin-11 expression and patient prognosis
The median follow-up period for all patients with GBLs was 19 months (range, 8–95). Relative to cadherin-11 staining intensity, PFS and OS plots are presented in Fig. 2, and survival outcomes are presented in Table 2.

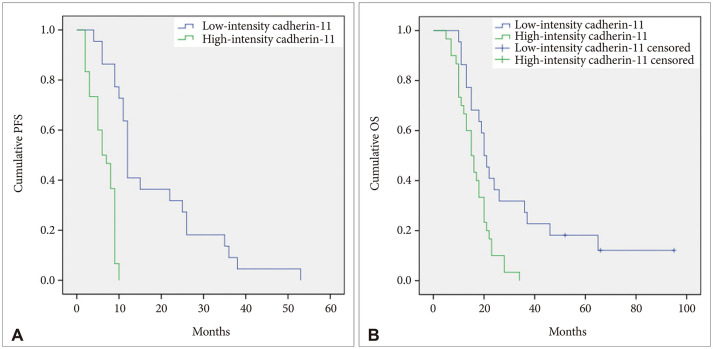 | Fig. 2The PFS (A) and OS (B) plots according to the staining intensity of cadherin-11.PFS, progression-free survival; OS, overall survival.
|
Table 2
Survival outcome of high-expression versus low-expression of cadherin-11
![]()
The median PFS was 9.0 months (95% CI, 8.4–9.6). The 1-year PFS rate was 17.3%, and the 2-year PFS rate was 13.5%. Relative to cadherin-11 staining intensity, the median PFS in the low-intensity cadherin-11 group (12.0 months; 95% CI, 11.1–12.9) was longer than the median PFS in the high-intensity cadherin-11 group (6.0 months; 95% CI, 3.7–8.3; p<0.001).
The median OS was 18.0 months (95% CI, 14.0–22.0). The 1-year OS rate was 75.0%, and the 2-year OS rate was 21.2%. Relative to cadherin-11 staining intensity, the median OS in the low-intensity cadherin-11 group (20.0 months; 95% CI, 11.8–16.6) was longer than the median OS in the high-intensity cadherin-11 group (15.0 months; 95% CI, 11.8–18.2; p=0.003).
Prognosis of newly diagnosed primary GBLs
Univariate analyses were done to determine parameters of prognostic significance, which were summarized in Table 3. Cadherin-11 staining intensity was the only statistically significant factor relative to PFS and OS (p<0.001 and p=0.005).
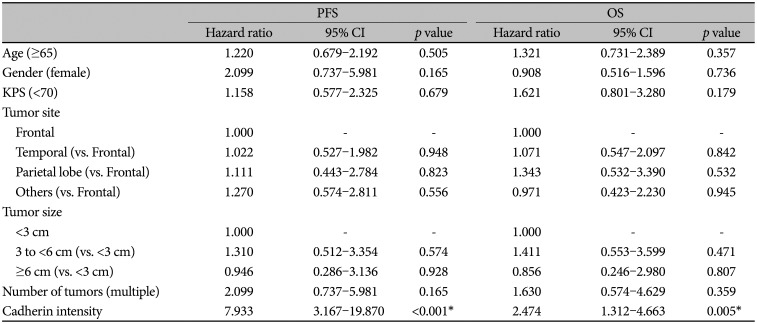
Table 3
Univariate analysis of prognostic factors on PFS and OS in patients with a newly diagnosed primary glioblastoma after gross-total resection
![]()
In multivariate analysis, cadherin-11 staining intensity was an independent predictor of poor PFS and OS (p<0.001 and p=0.005). High intensity of cadherin-11 raised the hazard for disease progression and survival (PFS: hazard ratio=7.933; 95% CI, 3.167–19.870, and OS: hazard ratio=2.474; 95% CI, 1.312–4.663) (Table 4).

Go to :
DISCUSSION
GBL presents variously relative to oncogenesis, limited treatment options, individualized treatment, and prognostic biomarkers. Moreover, there are various differences regarding our understanding of its etiology. Discovering additional biomarkers of GBL could lead to better patient classification for clinical trials and treatment protocols [17]. One large study, The Cancer Genome Atlas (TCGA), recently looked at genetic differences and protein expression correlated with GBL tumorigenesis. Nevertheless, proteins connected with GBL pathogenesis are still unclear relative to genetic alterations and protein expression correlated with the upstream etiology. As of late, increased cadherin-11 expression has been observed in several human neoplasms including breast, colon, prostate, renal cells, stomach, and osteosarcoma [71112]. Further, many studies suggest cadherin-11 expression may be correlated with glial tumors.
Cadherins are calcium-dependent cell-cell adhesion molecules that have interaction with catenins. Binding the intact cadherin-catenin complex to the actin cytoskeleton is crucial for cadherin-mediated cell-cell adhesion. Cell-cell adhesion, which is critical to ascertaining cell polarity and retaining tissue structure, is altered in tumorigenesis by several epigenetic and genetic modifications that increase tumor invasiveness and motility [18]. Some cadherins have been detected as probable oncogenic proteins and/or tumor suppressors, according to the type of cadherin and tumor [18]. In addition, cadherin plays a crucial role in epithelial-mesenchymal transition (EMT), which occurs in cancer progression and standard embryonic development [192021]. In standard tissue morphogenesis, EMT increases epithelial cell plasticity [20]. Nevertheless, tumor cells generate EMT into acquiring characteristics of a mesenchymal phenotype, such as increased invasiveness and motility, stem cell-like properties, aging, apoptosis, and further drug resistance [192021].
Many studies that profile global genetics have found abnormal mesenchymal lineages in GBL that may add to the heightened invasive ability of GBL [22]. In the development of a mesenchymal phenotype through EMT during tumor progression, E-cadherin expression loss is often correlated with mesenchymal cadherin expression, such as cadherin-11 or N-cadherin. This is known as cadherin conversion, and it occurs in prostate, breast, stomach, and pancreatic cancers [1823]. N-cadherin expression in tumors generates invasion, cell migration, and survival. Cadherin-11, a surface cell biomarker for EMT, is seen during development in many normal tissues, such as the mesenchymal tissue in the brain. In malignant prostate and breast cancers, the expression of cadherin-11 raises tumor cell migration and invasion [7111223]. And, the TGF-β signaling pathway is known to be an attractive candidate signaling pathway capable of regulating cadherin-11 expression by endothelial cells. Especially, TGF-β signaling plays an important role in both normal and pathogenic angiogenesis, and TGF-β activity confers tumor cell motility in GBL [24]. In accordance with previous studies, the current study demonstrated a correlation between cadherin-11 staining intensity and prognosis for patients with a newly diagnosed primary GBL. Specifically, there might exist a positive correlation between the expression of cadherin-11 in tumor tissue and poor prognosis relative to PFS and OS.
In addition, we investigated using TCGA dataset to more conclusively confirm the relationship between cadherin-11 expression and prognosis. In the TCGA dataset, there were 160 GBL patients with CDH-11 mRNA quantification data. According to correlation analysis, cadherin-11 expression revealed a negative correlation with OS, but there was no statistical significance (Pearson correlation coefficient: -0.102, p=0.424). This is considered to have some limitations in terms of the origin and treatment. To supporting the positive correlation between cadherin-11 expression and poor prognosis, the standardization of TCGA data is needed in the future.
Limitations
The present study has a few limitations. First, this was a single-institutional retrospective study. Therefore, selection bias could not be fully excluded. Second, there were a small number of patients with a newly diagnosed primary GBL. Therefore, the results of this study should be cautiously interpreted, and they should not be generalized. Third, there was no analysis of the interaction between cadherin-11 and other molecular markers. Based on the literature, there are various molecular markers related to prognosis in GBL including IDH mutation, MGMT methylation, 1p/19q codeletion, EGFR amplification, TERT promotor mutations, and PTEN deletion. In order to further elucidate the relationship between cadherin-11 and patients' prognosis with GBL, analyses about the relationship between cadherin-11 and molecular markers are needed. However, molecular analyses have been performed since 2012 in our institute and the data of molecular markers analyzed was very small, which made it difficult to perform statistical analyses. Therefore, the potential influence of other molecular markers could not be excluded. As a result, to demonstrate more definitively that cadherin-11 is a prognostic factor, a large prospective study is needed, and any association between cadherin-11 and molecular markers, such as genetic modifications and protein expressions associated with GBL tumorigenesis, should be investigated.
Conclusion
Cadherin-11 expression and clinical outcome may be positively correlated with primary GBL. While the results of the present study should be interpreted cautiously, we demonstrated in our study that the high intensity of cadherin-11 expression was associated with the PFS and OS of primary GBL.
Go to :
 XML Download
XML Download