

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Metastasis is a cancer that has spread beyond the point of origin to distant areas of the body [1]. Of all organs that these metastatic cells colonize, it is estimated that the brain is responsible for 9% to 17% of systemic cancers [2]. The occurrence of brain metastasis varies according to the type of primary cancer, occurring most frequently in primary lung (20%), melanoma (7%), and breast (5%) cancers [3]. As treatment modalities for primary cancers have improved, metastasis is becoming the major cause of morbidity and mortality in cancer patients [4]. The improved control of primary tumors results in longer survival and the possibility of brain metastasis is subsequently increased [5].
The mechanisms regulating and initiating brain metastasis are not clearly known due to lack of identifying the early stages of metastasis even though by using the detection modalities and visualizing techniques, which are hindering the development of preventive therapies [6]. Therefore, it is urgent to have experimental models in order to recapitulate clinical courses, to investigate the metastatic process, and to validate new therapeutic targets. However, few models can handle the range of unresolved questions. Thus, choosing an appropriate model depends on the scientific topics that the researcher is seeking. In this review, we summarize different experimental brain metastasis models. While most studies have used murine models, different models are also considered to complement these experimental resources and are introduced.
Go to : 
BRAIN METASTATIC PROCESS
After cancer cells have grown to form a primary tumor, several enzymes allow the cancer cells to invade local tissues by degrading basement membranes. The cancer cells can enter the blood stream or lymphatic system in a process called intravasation, by squeezing through the surface of blood vessels. Once within the circulatory system, they disseminate to distinct sites of the body and become lodged into the capillaries of other organs. These cells then undergo extravasation by moving through the vessel membranes and forming micrometastasis. At this point, the cancer cells can colonies, forming secondary tumors. Most cancer cells will not survive this process, particularly due to a hostile microenvironment. A period of latency is required [7]. Even if only organ colonization is considered, the number of successful metastatic cells that complete extravasation into the brain could be 1 in 100 [8]. The brain microenvironment is a unique compartment within the body. The resident cells, such as microglia, oligodendrocytes, astrocytes, and neurons, create a complex and dynamic microenvironment and are involved with metastatic cancer cells [9]. Additionally, the brain is separated from the peripheral vasculature by the blood–brain barrier (BBB), which is a selective filter that enters the systemic circulation. The BBB impairment is frequently observed in brain metastasis and the blood-tumor barrier (BTB) is created [10]. The formation of the tumor destroys the integrity of BBB and BTB, which causes damage to the original environment. This unique condition allows tumors located in the brain to have unique cell types, anatomical structures, metabolic constraints, and immune environments [11].
Go to :
IN VITRO MODELS OF METASTASIS
Even though the dynamic cascade of the metastasis makes it challenging to investigate each step precisely, many assays of high value have been developed. Scratch-wound and zone exclusion assays examine the migration and invasion of cancer cells [12]. This method has the variability in the locations selected at each experiment. Time-lapse microscopy allows the observation of real-time imaging of cell motility, from determining the rate of cell, characterizing the morphological changes achieved by metastatic cells and identifying the type of motility [1314].
The in vitro assay is largely two-dimensional (2D) lacking microenvironmental influence and cell to cell interactions. These deficiencies hinder the translation of the assays. Many efforts have been performed to develop three-dimensional (3D) assays retaining these essential interactions, such as assays incorporating stromal cells [15] and assays that allow adhesion [16]. The tumorsphere is spherical structures generated by cancer stem cells (CSCs). CSCs facilitate the migration. Genetic signatures in CSCs are thought to predict tumor recurrence and metastasis [1718].
Organoids generated from resected tumors can organize into hierarchical structures that reflect the original tissue [19]. Organoids have similarities to patient’s specimens than cells grown under 2D culture system [20]. A recent protocol using human cancer cells and embryonic stem cell-derived brain organoids successfully recapitulates the brain metastasis process and provides a useful platform for drug development [21].
The type of cell line can be selected according to the experimental purpose. Commercial cell lines are easily available and characterized by simple maintenance and consistent growth rates. Primary patient-derived cell lines are developed through several passages of culture. This cell line has a more accurate molecular characterization of the original patient tumor than commercial cell lines. Because these model systems serve to predict the efficacy of potential therapies, they help make clinical decisions and improve the application of personalized medicine [22].
Go to :
IN VIVO MODELS OF METASTASIS
The advantage of in vivo models to identify therapeutic candidates is their ability to replicate the clinical progression of a given disease (Table 1). Rodents have proven valuable for developing models that advance the knowledge of brain metastasis research [23] and evaluating novel candidate therapies [24]. Several patient-derived xenograft (PDX) models have been developed and evaluated for studying brain metastasis. PDXs can maintain better tumor heterogeneity, biology and microenvironment compared to in vitro models [25]. The use of these in vivo models has led to the proposal of relevant genetic alterations essential for metastatic progression and the identification of potential drug candidates to treat them.

Table 1
In vivo models for metastasis research
![]()
Syngeneic
The syngeneic mouse model is a brain metastasis model that involves direct injection of an allograft of immortalized mouse cancer cells into mice [26]. Both the donor mouse and the host mouse are usually from the same inbred lineage. Because the species of the cell origin and the host model are matched, the syngeneic model allows mice to maintain consistently competent immune system [27]. This feature ensures that the syngeneic model is particularly suitable for studying the interaction between cancer and immune cells as well as the efficacy of immunotherapy.
Humanized
Humanized models are created by grafting immune system tissues and immune cells derived from human into immunodeficient mice. In the beginning, this model was established by the transplantation of human-derived peripheral blood mononuclear cells or hematopoietic stem cells. Humanized mice could accept the xenograft steady growth and mimic human immune system. The establishment of cell-line-derived xenograft and PDX into humanized mouse models facilitate diverse experiments to explore cancer pathogenesis and therapeutic effects [28]. Mouse and human immune systems are fundamentally different [29]. However, humanized mouse models can be utilized to evaluate antitumor efficacy against human checkpoint molecules. Researchers use humanized mice as a more accurate model to study complex immune responses during immunotherapy [30]. New immunotherapeutic drugs underline the significance of humanized mouse models in reforming clinical practice.
Genetically engineered
Genetically engineered mouse models (GEMMs) are classified into two groups. Transgenic GEMMs are developed by inoculation of a zygote or embryonic stem cell in which an exogenous oncogene carries the construct of the gene of interest. Targeted GEMMs are achieved by incorporating homologous recombination into mouse embryonic stem cells [31]. With GEMMs, tumors develop in an environment of natural immunity, where the histopathological and molecular characteristics of the resulting tumor are very similar to those found in humans, and tumors can spontaneously metastasize. The benefit of GEMMs enable the study of the microenvironment and immunological components of the metastasis process, as well as mimicking the clinical state with the hallmarks of spontaneous metastatic development in immunocompetent hosts [32]. Because syngeneic cell lines can also be inoculated to GEMM, this model is a useful tool for the study of brain metastasis related to the immune system. This model forms brain metastasis more rapidly than the PDX model [3334].
Xenograft
PDX models of brain metastasis are established by transplanting fresh human cancer specimens or patient-derived cells cultured through early passage into immunodeficient or humanized mice. A deficiency of the immune system in the same species promotes a higher occurrence of tumor engraftment. The advantage of the PDX model is that it can better preserve the genomic, histopathological, and phenotypic heterogeneity of the original tissue. Therefore, this improves the screening of potential therapeutics and increases the value in assessing personalized medicine [3536].
Go to :
IN VIVO MODELS OF BRAIN METASTASIS
To generate brain metastasis in vivo, these cells are inoculated into animals via several routes (Table 2). The route of inoculation is intravenous (IV) and the dissemination of tumor cells varies due to circulation. Inoculation of cells via the tail vein often results in metastasis that progress primarily in the lungs and central nervous system. Delivery via this route has high survival rates and is easy to implement [37]. To avoid pulmonary metastasis of cells, inoculation through the left ventricle, called intracardiac, allows systemic circulation of cells throughout the body. The intracardiac model has a high survival rate in skilled hands and the procedure is relatively simple.
Table 2
In vivo models of brain metastasis
![]()
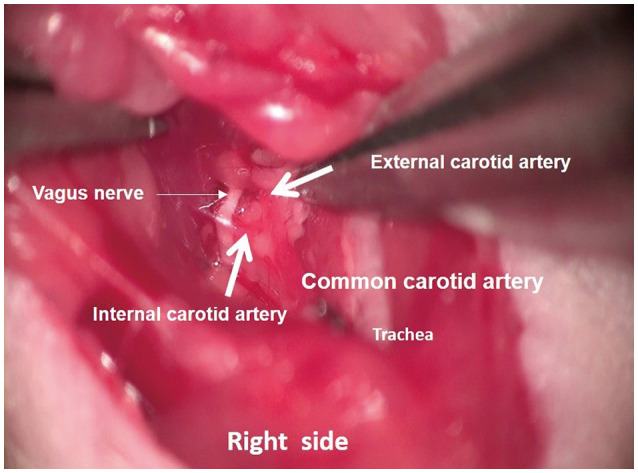
To minimize the spread of cancer cells to locations outside the brain, intra-carotid inoculation is preferred. This model can be time intensive, requires microsurgical techniques for ligation of the required arteries, and has high intraoperative mortality (Fig. 1, video clip at https://www.youtube.com/watch?v=TfoyzYVwRiA). Both methods involve robust selective steps of efflux, but early steps in the metastasis process, for example, invasion and formation of metastatic niches, are ignored [38].
Cancer cells are usually administered directly into the brain using a stereotaxic device. A single, precisely located, and established lesion is produced by the stereotactic procedure [3940]. However, intracranial inoculation does not fully represent metastatic cascades.
An ideal model of brain metastasis would require cancer cells to go through all stages of brain metastasis in orthotopiccally injected tumor cells, such as a mammary fat pad for breast cancer or a subcutaneous for melanoma or GEMM. They can spontaneously develop brain metastasis following the genetic manipulation of oncogenes or tumor suppressors. Due to the low incidence of brain metastases in this model, the high experimental variability requires a larger population of mice.
Go to :
METASTATIC BRAIN CANCER MODELS USING HUMAN CEREBRAL ORGANOIDS
One of the most human like model is the organoid. Recently, several protocols have been developed to generate cerebral organoids (COs) using human pluripotent stem cells (hPSCs) [414243]. CO is hPSC-derived organoids that self-assemble a form an organized architecture, composed of neural progenitors, neurons and glial cells. Unlike 2D cell cultures, CO recapitulates the human brain not only at the cellular level, but also in terms of general tissue structure [44]. Therefore, CO can overcome the limitations of metastatic mouse models [45]. A reproducible brain metastasis model is recently reported using human lung cancer cells and human embryonic stem cell (hESC)-derived CO [21]. Cancer cell proliferation, identification of specific gene functions, cell-cell interactions, and drug screening using metastatic brain cancer CO model were investigated. Such experiments are difficult to recapitulate in 2D cell cultures or animal models. The metastatic brain cancer CO model is an in vitro model for metastatic brain cancer and is located between 2D and animal models. However, it has an advantage to provide additional information over 2D or animal models. Although metastatic brain cancer CO models are promising, they have limitations such as the lack of important brain vasculature (e.g., BBB and BTB), immunological properties, the mature tissue, and the natural metastasis cascades when the real brain metastasis is considered. Disease modeling with new organoid techniques are also being developed through genetic and tissue engineering.
Even though 3D COs offer a complex model system that presents the opportunity to model various neurological diseases, there are still limitations. COs are different in size and shape, and the positions of the brain regions within each organoid also differ. Cells in brain organoids lack vascular systems as a result of the restricted culture techniques. The supply of gas and nutrients to COs mainly depends on simple diffusion from the medium, which causes a number of organoid cells to undergo apoptosis. In addition to the lack of a vascular system, stromal components, including microglia, are absent from current brain organoids, substantially limiting their application in the relevant research. Therefore, it is necessary to establish a circulatory system for long-term in vitro culture [46]. Recently, it is showen that on-chip hPSC-derived pericytes and endothelial cells sprout and self-assemble into organized vascular networks, and use COs to explore interactions with generated vasculature. Vascular cells physically interact with the CO and form an integrated neurovascular organoid on chip. The organoid vascularization approach opens several avenues for further studies in brain cancer and BBB [47].
Go to :
CONCLUSION
Brain metastasis remains an unmet medical problem because treatment is inadequate, but the incidence continues to rise. Various brain metastasis models provide an opportunity to investigate how cancer cells interact with cells in the brain microenvironment at different stages of the metastasis cascade. With the appropriate laboratory resources available, researchers can initiate and advance relevant research to change the current clinical reality of brain metastasis.
Go to :
 XML Download
XML Download