

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Obesity is a major health problem worldwide and acts as a risk factor for various diseases, including metabolic, cardiovascular, neuromuscular, and rheumatological diseases, by inducing chronic systemic low-grade inflammation in the human body. Many studies have also suggested that obesity is one of the main causes of the exacerbation of chronic inflammatory airway diseases, such as asthma and chronic obstructive pulmonary disease (COPD) [1,2]. Obesity may induce a chronic systemic inflammatory condition by various proinflammatory mediators released from adipose tissues (called adipokines). These mediators predispose the human and animal respiratory tract to a chronic inflammatory state and oxidative stress [1-3]. Recent evidence suggests that allergic rhinitis (AR) and chronic rhinosinusitis (CRS) are chronic inflammatory conditions rather than infectious conditions of the sinonasal mucosa and show pathophysiological mechanisms similar to those of chronic lower inflammatory airway diseases [4]. Therefore, the effects of obesity on AR or CRS have been studied extensively. Many studies have shown that obesity increases the incidence or exacerbates the symptoms of AR or CRS [5-7]. In particular, pro-inflammatory adipokines are closely implicated in obesity-related airway inflammatory responses. Mucus hypersecretion is known to be an important cause of the exacerbation of inflammatory sinonasal diseases. Increasing evidence indicates that pro-inflammatory adipokines play important regulatory roles in a variety of biological processes associated with mucus secretion in the sinonasal mucosa [6-9]. However, the underlying mechanisms and treatment strategies of sinonasal inflammatory diseases mediated by obesity are not clear.
Ghrelin, an endogenous ligand for the growth hormone secretagogue receptor 1a (GHSR1a), is released from gastric endocrine cells and plays a major role in stimulating growth hormone release from the pituitary gland. Ghrelin was shown to inhibit the expression of proinflammatory cytokines, such as tumor necrosis factor-alpha (TNF-α), interleukin (IL)-6, and IL-1β [10], while activating the expression of anti-inflammatory cytokines in various inflammatory diseases that are known to be closely related to obesity. Moreover, several recent studies have reported that ghrelin has potential therapeutic effects on lower inflammatory airway diseases, such as asthma, COPD, and cigarette smoking-mediated airway diseases, which are exacerbated by the inflammatory condition caused by obesity [11,12]. However, the effects and mechanisms of ghrelin on inflammatory sinonasal diseases, which are highly likely to be exacerbated by obesity, have not been confirmed. This study aimed to elucidate the effect of ghrelin on the regulation of mucus secretion in human nasal epithelial cells (HNEpCs) and the underlying mechanisms. Therefore, we investigated the potential therapeutic effects of ghrelin as a topical nasal treatment for obesity-related inflammatory sinonasal diseases.
Go to : 
MATERIALS AND METHODS
This work was approved by the Institutional Review Board of Yeungnam University Medical Center (No. YUMC 2016-05-040). Informed consent from patients was waived due to this study is conducted using human nasal mucosa epithelial cells commercially available from PromoCell (C-12620, donor age/sex/race 14/female/Caucasian, nasal mucosa, freezing medium Cryo-SFM).
Materials
Recombinant human ghrelin and the GHSR1a antagonist; D-Lys-3-growth hormone-releasing peptide 6 (D-Lys-3-GHRP-6) were obtained from Phoenix Pharmaceuticals. Lipopolysaccharide (LPS) was purchased from Sigma-Aldrich. Recombinant human leptin was purchased from R&D Systems. The HNEpCs were obtained from PromoCell (C-12620).
Cell culture and treatment
HNEpCs were cultured in an airway epithelial cell growth medium at 37 °C in humidified air with 5% CO2 and subcultured according to PromoCell recommendations. Evaluation of the effect of ghrelin on LPS/leptin-induced MUC5AC expression. Cells were pretreated with LPS (1 µg/mL) or leptin (0.1 µM) for 1 hour. Pretreated HNEpCs by LPS or leptin were exposed to different concentrations of ghrelin (0, 0.01, 0.1, and 1 µM). To confirm the involvement of ghrelin in LPS/leptin-induced MUC5AC expression, D-Lys-3-GHRP-6, one of the GHSR1a antagonists, was treated simultaneously with ghrelin (0.1 µM). The control group was incubated in a culture medium for the same period. The cytotoxicity of ghrelin with/without LPS (1 µg/mL) or leptin (0.1 µM) on was checked using water-soluble tetrazolium salt-1 assay. Various concentrations of ghrelin (0, 0.01, 0.1, 1, 5 μM) were treated with/without LPS or leptin to HNEpCs for 24 hours. At all concentrations used (up to 5 μM), Agents did not affect cell viability (data not shown).
Reverse transcription-polymerase chain reaction
Total RNA was prepared using TRIzol reagent (Invitrogen). Reverse transcription-polymerase chain reaction (RT-PCR) was performed using the GeneAmp RNA PCR Core Kit (Thermo Fisher Scientific). Two-step PCR was conducted according to the protocol provided by the Thermo Fisher Scientific. Murine leukemia virus reverse transcriptase was used for the reverse transcription of RNA into cDNA and AmpliTaq DNA polymerase was used for subsequent PCR amplification. The PCR products were quantified and normalized to glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The products were then detected using 2% agarose gel, visualized by staining with SYBR Safe DNA gel stain (Invitrogen) and transilluminated with ultraviolet (UV) light.
Real-time PCR
Real-time PCR was performed using the iQ SYBR Green Supermix (Bio-Rad). Following the manufacturer’s protocol, 1 ng/μL of total RNA was reverse transcribed. A maximum of 2 μL of each cDNA sample was used per 20 μL of PCR mixture. PCR reactions were performed using CFX96 real-time PCR system C1000 Thermal Cycler (Bio-Rad) with triplicate wells and 40 cycles. The primer sequences and conditions were used according to previously published experimental protocols [7,13].
Enzyme-linked immunosorbent assay
Supernatants and lysates of HNEpCs were diluted with phosphate-buffered saline (PBS). Each sample was transferred to Nunc-Immuno Maxisorp 96 well plate (Thermo Fisher Scientific) and cultured at overnight (4 °C). The samples were blocked with 2% bovine serum albumin (BSA) for 1 hour and incubated with primary antibodies (rabbit anti-MUC5AC, sc-20118; Santa Cruz Biotechnology) for 1 hour in PBS containing Tween 20 (0.05%). Horseradish peroxidase (HRP)-conjugated secondary antibodies (goat anti-rabbit immunoglobulin G [IgG]-HRP; sc 2004) were applied to each well. After 4 hours, a color was formed using a 3,3´, 5,5´-tetramethylbenzidine peroxidase solution and stopped with 2N-H2SO4. Optical density measurements were performed using an EL800 enzyme-linked immunosorbent assay (ELISA) reader (BIO-TEK Instruments) at 450 nm. The results are expressed as a fold increase from the baseline control.
Western blot
Cell lysate samples were separated by 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred to a polyvinylidene fluoride membrane. The membranes were incubated overnight (4 °C) with the primary antibody. After washing, the blots were incubated with a secondary antibody for 1 hour. Bands were detected after exposing the membrane to an imaging system (Chemiluminescence Imaging System, FUSION-FX7 820wL).
Immunofluorescence staining
The HNEpCs were incubated until confluence on glass cover slips coated with poly-L-lysine (0.05%) in a growth medium for airway epithelial cells for 48 hours. Cells were pretreated with ghrelin (0.1 μM) with or without D-Lys-3-GHRP-6 (1 μM) for 1 hour. Pretreated cells were exposed to LPS (1 μg/mL) or leptin (0.1 μM) for 8 hours. The HNEpCs were fixed with formaldehyde (4%) in PBS for 15 minutes and blocked with BSA (5%) in PBS overnight. The HNEpCs were then incubated with MUC5AC antibodies (ab-198294; 1:100; Abcam). Subsequently, the samples were incubated with Alexa-488-labeled goat anti-rabbit IgG secondary antibodies. Nuclei were detected with 4´,6-diamidino-2-phenylindole (DAPI; AbCam). Stained samples were visualized by fluorescence microscopy (×40 magnification, Ti-S, 733551; Nikon). The intensity was measured using an infinite F200 PRO microplate fluorescence reader (Tecan).
Statistical analysis
Statistical analyses were performed using IBM SPSS ver. 22.0 (IBM Corp.). Means and standard deviations were calculated for the sets of measurements. Data were compared using an unpaired t-test or Kruskal-Wallis one-way analysis of variance followed by the Mann-Whitney test. For all tests, a P-value <0.05 was considered statistically significant.
Go to :
RESULTS
The presence of GHSR1a in HNEpCs
To identify the expression pattern of GHSR1a in HNEpCs, the HNEpCs were treated with ghrelin (0.1 μM) for 12 hours. PCR showed that ghrelin significantly induced GHSR1a messenger RNA (mRNA) expression (Fig. 1A and B). The effect of ghrelin on LPS-induced MUC5AC in HNEpCs
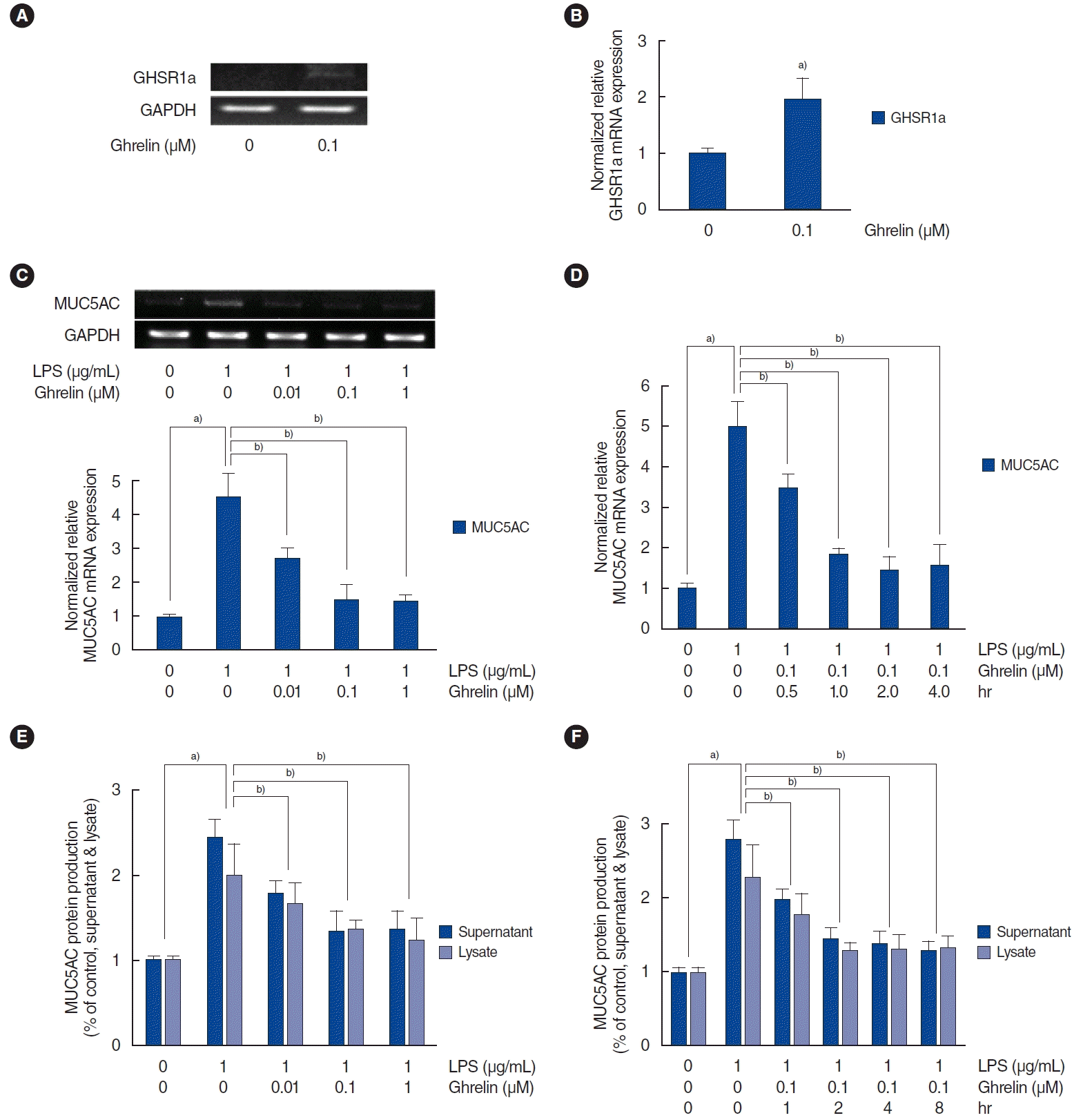 | Fig. 1.Effect of ghrelin on lipopolysaccharide (LPS)-induced MUC5AC expression in human nasal epithelial cells. (A, B) The results of reverse transcription-polymerase chain reaction (RT-PCR) and real-time PCR show that ghrelin significantly induced GHSR1a messenger RNA (mRNA) expression. (C) The results of RT-PCR and real-time PCR show that ghrelin significantly inhibited LPS-induced MUC5AC mRNA expression. LPS-induced MUC5AC expression started to be maximally suppressed at ta ghrelin concentration of 0.1 μM. (D) The results of real-time PCR show that the maximal inhibition of LPS-induced MUC5AC mRNA expression by ghrelin (0.1 μM) occurred after 2 hours. (E, F) The results of enzyme-linked immunosorbent assay also show that ghrelin significantly inhibited LPS-induced MUC5AC protein production. The images are representative of three separate experiments performed in triplicate. The bars indicate the mean±standard deviation of three independent experiments performed in triplicate. GHSR1a, growth hormone secretagogue receptor 1a; GAPDH, glyceraldehyde-3-phosphate dehydrogenase. a)P<0.05 compared to the baseline value. b)P<0.05 is compared with samples treated with LPS (1 μg/mL) alone.
|
The dose- and time-dependent effects of ghrelin on LPS-induced
MUC5AC expression and HNEpCs were assessed by pretreatment with LPS (1 µg/mL) before exposure to different doses (0, 0.01, 0.1, and 1 μM) of ghrelin. HNEpCs pretreated with LPS (1 µg/mL) were exposed to ghrelin (0.1 μM) for different time lengths (0.5, 1, 2, or 4 hours). RT-PCR and real-time PCR results showed that ghrelin significantly inhibited LPS-induced MUC5AC mRNA expression (Fig. 1C). LPS-induced MUC5AC expression started to be maximally suppressed at a ghrelin concentration of 0.1 μM. Furthermore, real-time PCR showed that LPS-induced MUC5AC mRNA expression was suppressed by ghrelin (0.1 μM) for up to 4 hours. The maximal inhibition of LPS-induced MUC5AC mRNA expression by ghrelin (0.1 μM) was observed after 2 hours (Fig. 1D). ELISA showed that ghrelin significantly inhibited LPS-induced MUC5AC protein production (Fig. 1E and F).
The inhibitory mechanism of ghrelin on LPS-induced MUC5AC in HNEpCs
RT-PCR, real-time PCR, ELISA, Western blotting, and immunofluorescence staining were conducted to evaluate the mechanism by which ghrelin inhibits LPS-induced MUCAC expression. RT-PCR and real-time PCR showed that ghrelin inhibited LPS-induced MUC5AC overexpression, which was abolished by D-Lys3-GHRP-6 (Fig. 2A and B). ELISA and immunofluorescence staining results also showed that ghrelin significantly inhibited LPS-induced MUC5AC protein overproduction, while the inhibitory effect of ghrelin on LPS-induced MUC5AC protein overproduction was abolished by D-Lys-3-GHRP-6 (Fig. 2C and D). Western blotting was also performed to confirm the involvement of the extracellular signal-regulated kinase 1/2 (ERK1/2) and p38 mitogen-activated protein kinase (MAPK) signaling pathways in the inhibitory mechanism of ghrelin on LPS-induced MUC5AC expression. Ghrelin significantly inhibited LPS-activated ERK1/2 and p38 MAPK. These ghrelin-mediated changes in MAPK activation were abolished by D-Lys-3-GHRP-6 (Fig. 2E).
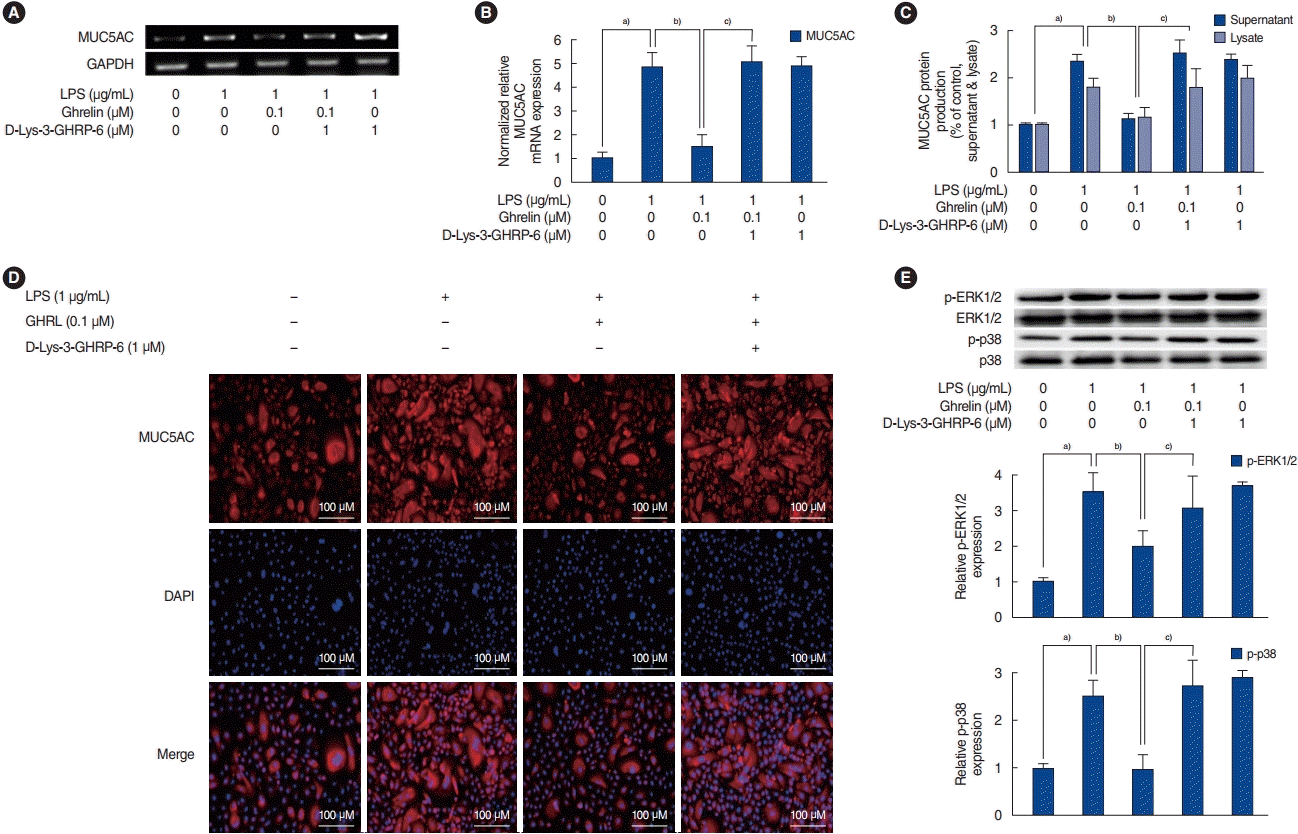 | Fig. 2.The regulatory mechanism of ghrelin on lipopolysaccharide (LPS)-induced MUC5AC expression in human nasal epithelial cells. (A, B) The results of reverse transcription-polymerase chain reaction (RT-PCR) and real-time PCR show that ghrelin inhibited LPS-induced MUC5AC expression, and this inhibition was abolished by D-Lys-3-growth hormone-releasing peptide 6 (D-Lys-3-GHRP-6). (C, D) The results of enzyme-linked immunosorbent assay and immunofluorescence staining also show that ghrelin significantly inhibited LPS-induced MUC5AC protein production, while the inhibitory effect of LPS-induced MUC5AC protein production was abolished by D-Lys-3-GHRP-6. (E) Western blot results show that ghrelin significantly inhibited LPS-activated extracellular signal-regulated kinase 1/2 (ERK1/2) and p38 mitogen-activated protein kinases (MAPKs). These ghrelin-mediated changes in MAPK activation were also abolished by D-Lys-3-GHRP-6. GAPDH, glyceraldehyde-3-phosphate dehydrogenase; DAPI, 4´,6-diamidino-2-phenylindole; p-ERK1/2, phosphorylated ERK 1/2; p-p38, phosphorylated p38. a)P<0.05 compared to the baseline value. b)P<0.05 compared with samples treated with LPS (1 μg/mL) alone. c)P<0.05 compared to samples treated with LPS (1 μg/mL) and ghrelin (0.1 μM).
|
The effect of ghrelin on leptin-induced MUC5AC in HNEpCs
To confirm the effect of ghrelin on leptin-induced MUC5AC expression, HNEpCs were pretreated with leptin (0.1 µg/mL) before exposure to different concentrations (0, 0.01, 0.1, and 1 μM) of ghrelin. HNEpCs pretreated with leptin (0.1 µg/mL) were exposed to ghrelin (0.1 μM) for different time lengths (0.5, 1, 2 or 4 hours). RT-PCR and real-time PCR showed that ghrelin significantly inhibited leptin-induced MUC5AC mRNA expression (Fig. 3A). Leptin-induced MUC5AC expression was maximally suppressed at a ghrelin concentration of 0.1 μM. Additionally, real-time PCR showed that leptin-induced MUC5AC mRNA expression was suppressed by ghrelin (0.1 μM) for up to 4 hours. The maximal inhibition of leptin-induced MUC5AC mRNA expression by ghrelin (0.1 μM) was observed after 2 hours (Fig. 3B). ELISA showed that ghrelin significantly inhibited leptin-induced MUC5AC protein production (Fig. 3C and D).
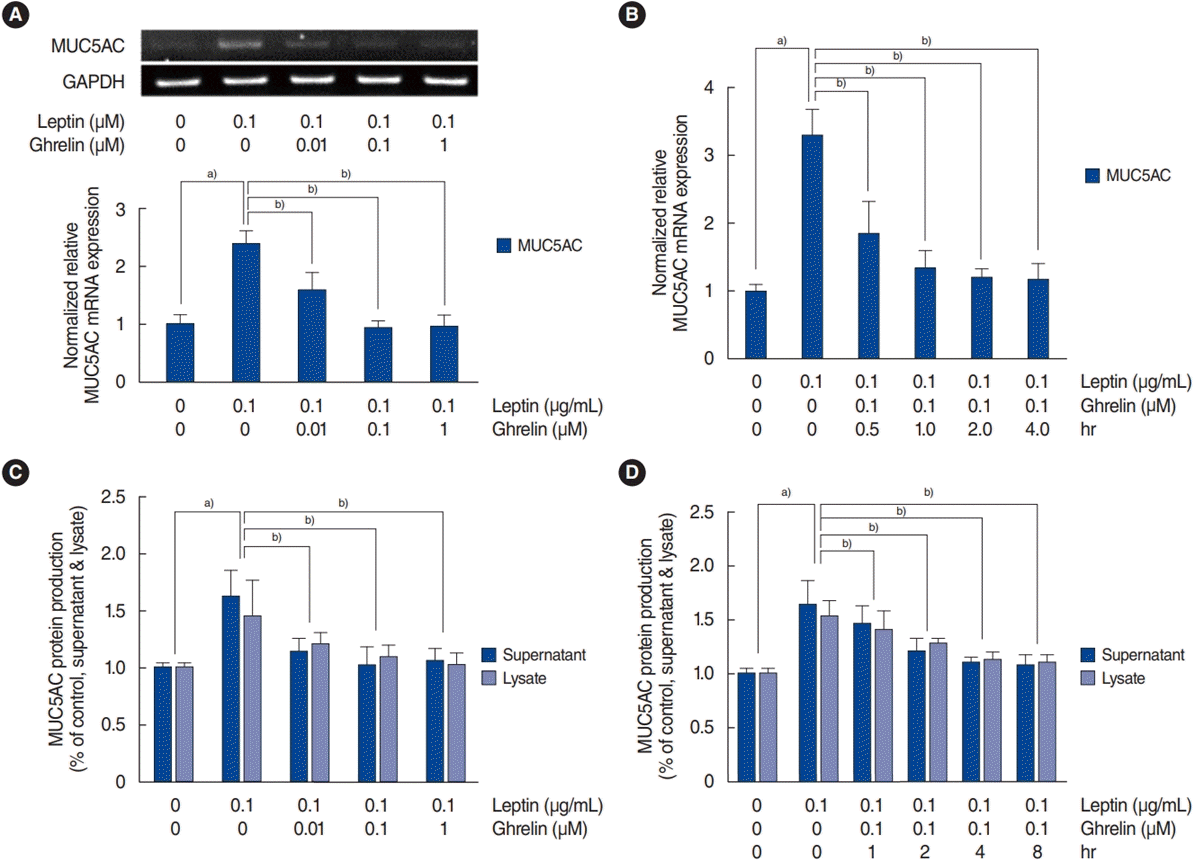 | Fig. 3.Effect of ghrelin on leptin-induced MUC5AC expression in human nasal epithelial cells. (A) The results of reverse transcription-polymerase chain reaction (RT-PCR) and real-time PCR show that ghrelin significantly inhibited leptin-induced MUC5AC mRNA expression. Leptin-induced MUC5AC expression started to be maximally suppressed at a ghrelin concentration of 0.1 μM. (B) Real-time PCR results show that the maximal inhibition of leptin-induced MUC5AC mRNA expression by ghrelin (0.1 μM) occurred after 2 hours. (C, D) The results of enzyme-linked immunosorbent assay also showed that ghrelin significantly inhibited leptin-induced MUC5AC protein production. The images are representative of three separate experiments performed in triplicate. The bars indicate the mean±standard deviation of three independent experiments performed in triplicate. GAPDH, glyceraldehyde-3-phosphate dehydrogenase. a)P<0.05 compared to the baseline value. b)P<0.05 compared with samples treated with lipopolysaccharide (LPS; 1 μg/mL) alone.
|
The inhibitory mechanism of ghrelin in leptin-induced MUC5AC in HNEpCs
RT-PCR, real-time PCR, ELISA, Western blotting, and immunofluorescence staining were conducted to evaluate the mechanism by which ghrelin inhibited leptin-induced MUCAC expression. RT-PCR and real-time PCR showed that ghrelin inhibited leptininduced MUC5AC overexpression, which was abolished by D-Lys-3-GHRP-6 (Fig. 4A and B). ELISA and immunofluorescence staining also showed that ghrelin significantly inhibited leptin-induced MUC5AC protein overproduction, while the inhibitory effect of ghrelin on leptin-induced MUC5AC protein overproduction was abolished by D-Lys-3-GHRP-6 (Fig. 4C and D). Furthermore, Western blotting was performed to confirm the involvement of the ERK1/2 and p38 MAPK signaling pathways in the inhibitory mechanism of ghrelin on leptin-induced MUC5AC expression. Ghrelin significantly inhibited leptin-activated ERK1/2 and p38 MAPK. These ghrelin-mediated changes in MAPK activation were abolished by D-Lys-3-GHRP-6 (Fig. 4E).
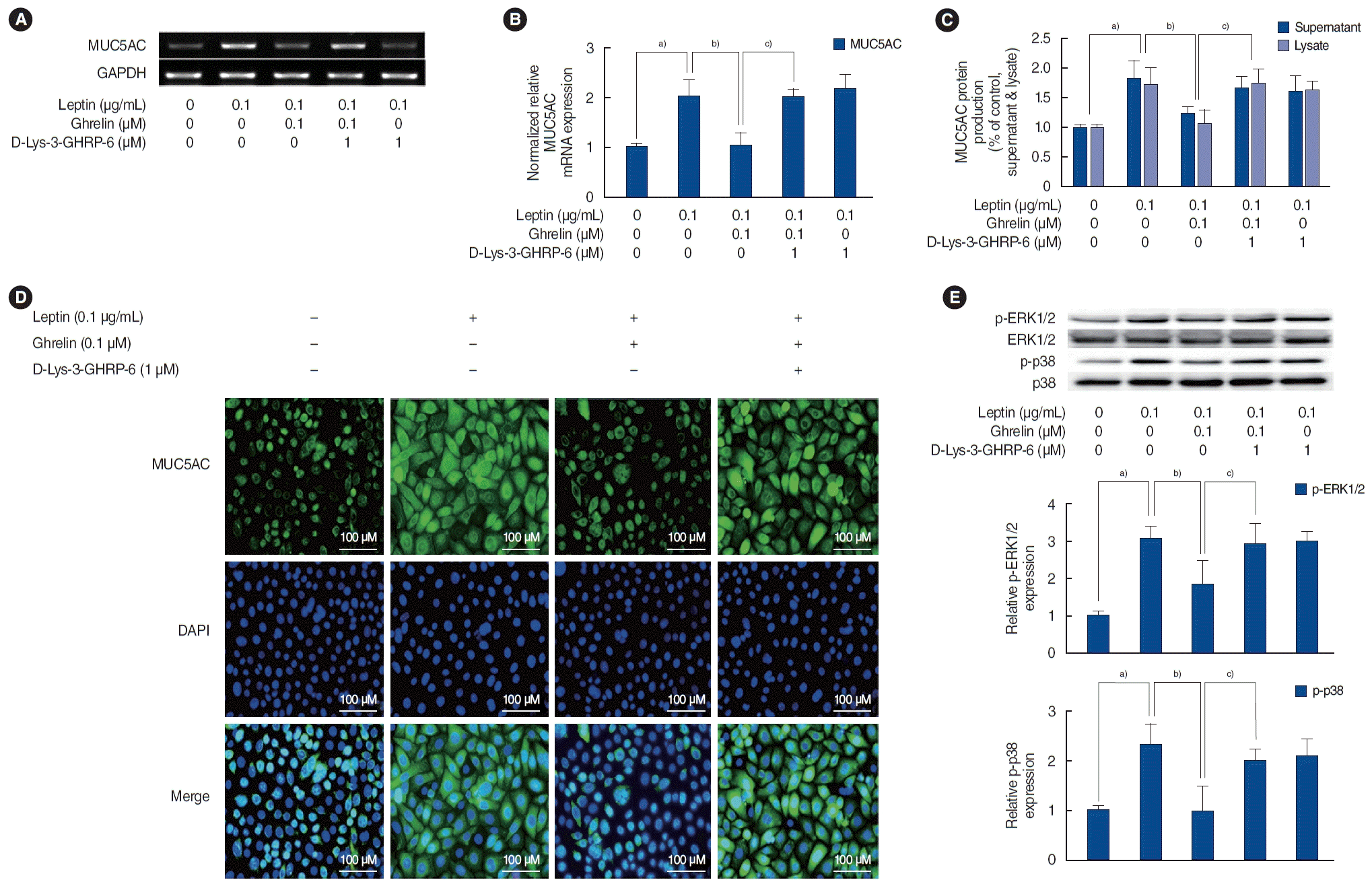 | Fig. 4.The regulatory mechanism of ghrelin in leptin-induced MUC5AC in human nasal epithelial cells. (A, B) The results of reverse transcription-polymerase chain reaction (RT-PCR) and real-time PCR show that ghrelin inhibited leptin-induced MUC5AC expression, and this inhibition was abolished by D-Lys-3-growth hormone-releasing peptide 6 (D-Lys-3-GHRP-6). (C, D) The results of enzyme-linked immunosorbent assay and immunofluorescence staining also show that ghrelin significantly inhibited leptin-induced MUC5AC protein production, while the inhibitory effect of leptin-induced MUC5AC protein production was abolished by D-Lys-3-GHRP-6. (E) Western blot results show that ghrelin significantly inhibited leptin-activated extracellular signal-regulated kinase 1/2 (ERK1/2) and p38 mitogen-activated protein kinases (MAPKs). These ghrelin-mediated changes in MAPK activation were also abolished by D-Lys-3-GHRP-6. GAPDH, glyceraldehyde-3-phosphate dehydrogenase; DAPI, 4´,6-diamidino-2-phenylindole; p-ERK1/2, phosphorylated ERK 1/2; p-p38, phosphorylated p38. a)P<0.05 compared to the baseline value. b)P<0.05 compared to samples treated with leptin (0.1 μg/mL) alone. c)P<0.05 compared to samples treated with leptin (0.1 μg/mL) and ghrelin (0.1 μM).
|
Go to :
DISCUSSION
Ghrelin is a hormone predominantly released from cells in the gastric mucosa of the fundus. Most circulating ghrelin originates from cells in the gastrointestinal tract and pancreas [10,14]. GHSR1a is a functional receptor for ghrelin. The activation of the ghrelin-GHSR1a system is known to have various physiological functions, such as the release of growth hormone and insulin, the control of energy metabolism, orexigenic regulation, and antiinflammatory and antioxidative effects [11,14]. GHSR1a is expressed in many organs, including the liver, intestine, kidney, heart, brain (especially the hypothalamus), and adipose tissue. It is also distributed in immune cells and bronchial epithelial cells in the respiratory tract [10-14]. Recent studies have demonstrated that ghrelin may be a novel therapeutic agent for chronic inflammatory airway diseases. In an asthmatic mouse model, ghrelin significantly downregulated ovalbumin-mediated inflammatory cytokines, including TNF-α, interferon-gamma, IL-5 and IL-13, and prevented endoplasmic reticulum stress activation by stimulating the Akt signaling pathway [15]. In cigarette smoke extract (CSE)-mediated airway inflammation, the predominant pathogenetic factor in the development of COPD, ghrelin inhibited CSE-induced IL-6 production by regulating the nuclear factor kappa-light-chain-enhancer of activated B cells pathway in human bronchial epithelial cells [11]. In an acute lung injury animal model, ghrelin attenuated lung damage by reducing the production of inflammatory cytokines and chemokines by alveolar macrophages [16]. However, no studies have investigated the effects of ghrelin on MUC5AC expression (as a marker of mucus hypersecretion in inflammatory sinonasal diseases) associated with local and systemic inflammation mediated by infectious or metabolic disorders. In the current study, we first identified that GHSR1a is expressed in HNEpCs and that ghrelin inhibits mucin gene overexpression induced by LPS or proinflammatory adipokines (leptin). These results suggest that the activation of the ghrelin-GHSR1a system may suppress mucus hypersecretion and goblet cell metaplasia caused by proinflammatory mediators (LPS or leptin) induced by local or systemic inflammatory conditions occurring in the human nasal mucosa.
Inflammatory sinonasal diseases are characterized by impaired function of the nasal mucosal epithelial immune barrier. LPS has emerged as a major stimulator of upper and lower airway inflammation. Exposure to LPS is implicated in airway epithelium integrity and the initiation and development of inflammatory sinonasal diseases [17,18]. Mucus hypersecretion is known to be an important pathogenetic factor in the exacerbation of inflammatory sinonasal diseases [19,20]. Among airway mucins, MUC5AC is known as a marker of goblet cell metaplasia, traditionally associated with inflammatory conditions of the epithelium of sinonasal mucosa [21,22]; therefore, clarification of the mechanisms through which the overexpression of MUC5AC is downregulated is essential for identifying treatment strategies for inflammatory sinonasal diseases. LPS is also known to induce MUC5AC overexpression in HNEpCs [17,18]. The current study also showed that LPS increased MUC5AC mRNA expression and MUC5AC glycoprotein production. Ghrelin significantly inhibited LPS-induced MUC5AC expression. Furthermore, the inhibitory effect of ghrelin on LPS-induced MUC5AC expression was eliminated by the GHSR1a antagonist.
Leptin, a proinflammatory adipokine predominantly released from adipocytes, plays a key role in the regulation of food intake, body weight, and energy metabolism. Several recent studies have reported that serum leptin levels increase in various inflammatory airway diseases, including COPD, asthma, AR, CRS, and nasal polyposis [1,2,5,23-25]. Leptin receptors are expressed in bronchial epithelial cells, airway immune cells, and nasal epithelial cells [1-3]. Many recent studies have suggested that upper and lower inflammatory airway diseases mediated by obesity are closely associated with epithelial cell proliferation and mucus hypersecretion caused by activation of the leptin-leptin receptor system [1-3,7,8]. Leptin also has profound effects on both the innate and adaptive immune systems, potentially impacting obesity-related inflammatory airway diseases. Leptin induces neutrophil chemotaxis and the generation of reactive oxygen species, activates immune cells (natural killer cells, macrophages), and increases the release of inflammatory cytokines [1-3]. In our previous studies, overexpression of the leptin receptor was observed in human nasal polyps. The overexpression of major airway mucins was confirmed to be induced by leptin in the human nasal mucosa. These findings suggest that obesity is closely related to nasal polyp formation and nasal mucus hypersecretion [7,8]. However, the precise mechanism underlying the inflammatory responses of the sinonasal mucosa in obese patients has not been clearly elucidated. There are limitations to creating experimental conditions similar to the nasal mucosa of patients with obesity and there is no clearly proven cell line model. However, unlike cytokines secreted from adipose tissues, such as TNF-α, IL-6, and IL-1β, leptin is an obesity-specific cytokine. Therefore, leptin was used as a substance to create conditions similar to the nasal mucosa of obese patients in the present study. Furthermore, LPS was also used to investigate whether ghrelin inhibits mucus hypersecretion induced by other stimulators, such as LPS, as well as leptin. Similar to the results of LPS treatment, the results of this study showed that leptin also increased MUC5AC mRNA expression and MUC5AC glycoprotein production. Ghrelin significantly inhibited leptin-induced MUC5AC expression. Furthermore, the inhibitory effect of ghrelin on leptin-induced MUC5AC expression was eliminated by treatment with a GHSR1a antagonist.
MAPK pathways are major signaling cascades that regulate various cellular processes. Among MAPKs, ERK1/2 and p38 are key cellular regulatory mechanisms of the inflammatory response in the human respiratory tract. These pathways play a role in the production of inflammatory cytokines, viral replication, immune cell activation, and mucus production, which are important processes in inflammatory airway diseases [26]. In our previous studies, different airway mucins including MUC 4 and MUC5AC/B, are regulated by various stimuli through the ERK1/2 and p38 signaling pathways [8,9,27]. MAPK pathways are also important regulatory mechanisms in airway inflammatory responses mediated by proinflammatory adipokine such as leptin. Leptin induces allergic airway inflammation by targeting the unfolded protein response via MAPK pathways in Th2 cells [28]. Moreover, the effect of leptin on Th2 cell proliferation, survival, and cytokine production is also dependent on MAPK pathways [29]. Furthermore, the anti-inflammatory effect of ghrelin is also known to occur through the inhibition of activated MAPK pathways. Recent studies suggested that ghrelin can protect against cerebral microvascular leakage and pericyte damage by effectively downregulating the expression of proinflammatory cytokines through inhibition of p38 phosphorylation [29]. In addition, ghrelin was found to inhibit angiotensin II-induced expression of TNF-α, IL-8, and monocyte chemoattractant protein-1 by inhibiting ERK1/2 MAPK activation in human endothelial cells [30]. In the current study, the activation of the p38 and ERK1/2 MAPK signaling pathways by LPS or leptin was significantly inhibited by ghrelin. These ghrelin-mediated changes in MAPKs activation were reversed by treatment with a ghrelin receptor antagonist. Therefore, ghrelin could inhibit LPS/leptin-induced MUC5AC overexpression in HNEpCs by inhibiting LPS/leptin-induced ERK 1/2 and p38 MAPK activation.
However, there are some limitations of this study. We only focused on the effect of ghrelin on mucus secretion in the nasal mucosa epithelium. Therefore, more precise evidence could not be obtained because we did not investigate the effect of ghrelin on the various cytokines involved in T helper (Th) cell-mediated inflammatory response, which is an important mechanism of inflammatory sinonasal disease. Future studies should confirm the effect of ghrelin on the expression of Th cell-mediated cytokines as well as mucus secretion through additional experiments. In addition, the effect of ghrelin on the expression of MUC5AC was only evaluated by an in vitro study using a single type of submerged culture HNEpCs, but not in other types of cells grown in different methods, such as air-liquid interface cultures or ex vivo cultures. Furthermore, additional experiments with animal models and human tissue sampling are needed to obtain more precise evidence regarding the therapeutic effects of ghrelin on inflammatory sinonasal diseases.
In conclusion, we confirmed that ghrelin inhibits LPS/leptin-induced MUC5AC expression in the human nasal epithelium. This is considered a meaningful basic study that identifies the potential therapeutic effect of ghrelin as a nasal topical treatment in patients with inflammatory sinonasal disease. It also shows the possibility of using alternative treatments for inflammatory sinonasal diseases in obese patients who face many limitations in medical treatment.
Go to :
HIGHLIGHTS
▪ Ghrelin inhibits lipopolysaccharide (LPS)/leptin-induced MUC5AC expression in human nasal epithelial cells.
▪ The inhibitory effect of ghrelin on LPS/leptin-induced MUC5AC is regulated by mitogen-activated protein kinase (MAPK) pathways.
▪ Ghrelin could be a topical agent for treating obesity-related inflammatory sinonasal diseases.
Go to :
 XML Download
XML Download