

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Breast cancer, as the malignant tumor with the highest incidence, seriously threatens women's health worldwide. Currently, the incidence of breast cancer shows a trend toward younger women [1]. Triple-negative breast cancer (TNBC) is highly invasive and easily metastasizes, and patients tend to have a poor prognosis due to the lack of effective therapeutic targets. Based on previous reports, TNBC patients suffer from a high recurrence rate and short overall survival within the first five years [2]. Therefore, current research is focused on finding effective therapeutic targets and improving the prognosis for TNBC.
Exosomes are nano-scale extracellular vesicles usually excreted by various cells, e.g., tumor cells [3]. The biological functions of tumor cell-derived exosomes are widely investigated due to their intriguing properties. Exosomes derived from tumor cells contain various biological molecules, including proteins, miRNAs, mRNAs, lipids and so on [4], which could act as the message molecules in intercellular communication. Recent study has highlighted the role of tumor-derived exosomes in educating macrophages, which polarizes macrophage to an anti-inflammatory phenotype to facilitate the tumor progression [5].
As a member of the RAS family, Rab proteins are a key regulator of intracellular endosome transport. Rab proteins with mutations or posttranslational modifications dysregulate intracellular vesicle trafficking, which may contribute to tumorigenesis [6]. Rab5 participates in the key steps of protein sorting to regulate the early endocytic pathway [7], which is essential for intracellular homeostasis and biosynthesis [8]. Studies have reported ectopic Rab5 expression in various tumors [9], and the potential underlying mechanism includes exosome secretion in cancer cells and stromal cells regulated by Rab5, which impact communication among cancer cells, immune cells and the microenvironment to form a tumor microenvironment that facilitates the growth of tumor cells [10]. It has been reported that silencing Rab5 and Rab27 impairs exosome secretion in cervical cancer cells [11]. In addition, the aberrant expression of Rab5 promotes the release of matrix metalloproteinase by stimulating the endocytosis/exocytosis cycle, thus enhancing the invasion of tumor cells [12]. Additionally, Rab5 knockdown was observed to delay the kinetics of FAK dephosphorylation and phosphorylation on Y397 and focal adhesion disassembly, consequently regulating cancer cell migration and invasion [13].
In conclusion, we speculated that the biological characteristics of TNBC, including frequent recurrence and metastasis, might be related to the abnormal expression of Rab5. Therefore, we examined the expression of Rab5 in TNBC as well as other subtypes and normal tissue. We generated Rab5 knockdown MDA-MB-231 cells using small interfering RNA (siRNA), and migration, invasion and exosome secretion were then determined to investigate the role of Rab5 in TNBC tumorigenesis and progression.
METHODS
Sample collection
Eleven breast cancer tissues and four normal breast gland tissues were collected in the Department of Breast and Thyroid Surgery, Xinjiang Medical University Affiliated Tumor Hospital; all patients provided written informed consent recognizing the purpose of the study. The protocol was approved by the Ethics Committee of Xinjiang Medical University Affiliated Tumor Hospital. The tumor tissues were identified by independent pathologists, including luminal A (3 patients), luminal B (4 patients), human epidermal growth factor receptor 2 (HER-2)-positive (4 patients), and triple-negative (4 patients).
Cell culture
MDA-MB-231 cells and the human monocytic cell line THP-1 were purchased from Procell Life Science & Technology Co., Ltd. Cells were cultured in DMEM containing 10% fetal bovine serum and 1% penicillin‒streptomycin at 37°C and 5% CO2. Cells were digested using ztrypsin upon reaching 80% confluence, and the cells in the logarithmic growth phase were used for subsequent experiments.
Cell transfection
MDA-MB-231 cells were seeded in a 24-well plate (5 × 104 cells/well), and 50 μl of Block-iT Alexa Fluor Red (Thermo Fisher), lipofectamine RNAiMAX (Thermo Fisher) and siRNA or vectors were added to the plate and incubated at 37°C for 24 or 48 h. The fluorescence was captured using fluorescence microscopy (Eclipse TS100-F; Nikon) to determine the transfection efficiency. The sequences of siRNAs used in this study were as follows: siNC, 5'-UUCUCCGAACGUGUCACGUTT-3' and 5'-ACGUGACACGUUCGGAGAATT-3'; siRAB5A#1, 5'-GGAAUCAGUGUUGUAGUAATT-3' and 5'-UUACUACAACACUGAUUCCTT-3'; siRAB5A#2, 5'-CCAGUUCAAACUAGUACUUTT-3' and 5'-AAGUACUAGUUUGAACUGGTT-3'; and siRAB5A#3, 5'-GGCAAGCAAGUCCUAACAUTT-3' and 5'-AUGUUAGGACUUGCUUGCCTT-3'.
Quantitative RT-PCR (qRT-PCR)
RNA was extracted using the TRIzol method (Invitrogen), and cDNA was synthesized using a 5x All-In-One RT MasterMix kit (Applied Biological Materials). Quantitative RT-PCR was conducted using EvaGreen Express 2×qPCR MasterMix-Low Rox (Applied Biological Materials) on an ABI QuantStudio 6 Flex Real-Time PCR System (Applied Biosystems). The primers for Rab5 and GAPDH are listed in Table 1.
Table 1
Primer information of qRT-PCR
![]()
Western blot
Total protein was extracted and quantified using a BCA kit (TransGen Biotech). Total proteins were isolated by SDS-PAGE and then transferred to PVDF membranes (Millipore). After blocking with 5% nonfat milk, primary antibodies were added and incubated overnight. TBST buffer was used to wash the membrane, and then secondary antibody was added and incubated at room temperature for 1 h. The blot signals were visualized using SuperSignal West Pico PLUS Chemiluminescent Substrate (Thermo Fisher). The primary and secondary antibodies used in this study were as follows: anti-Rab5 (ab218624; Abcam), anti-CD163 (ab182422; Abcam), anti-CD63 (ab134045; Abcam), anti-CD9 (ab92726; Abcam), anti-β-actin (D110001; D110001; Sangon Biotech), and goat anti-rabbit IgG H&L (HRP) (ab205718; Abcam).
Cell migration and invasion
Cells were plated on the upper chamber of the Transwell (Corning Costar) in serum-free medium, and the lower chamber was filled with complete medium. Cells were incubated at 37°C for 24 h and subsequently washed using PBS and fixed using 4% formaldehyde. Cells were then stained with Giemsa, and the cells on the surface of the upper chamber were removed. The migrating cells were observed under a light microscope, and three random fields were selected for counting. For the invasion assay, the only difference was that the Transwell insert was coated with Matrigel. Each assay was conducted in triplicate.
Isolation of exosomes
The supernatant (8 ml) was collected from the cell medium, and the exosomes were extracted using an exoEasy Maxi Kit (Qiagen) according to the manufacturer’s instructions.
Nanoparticle tracking analysis (NTA)
The isolated exosomes were first diluted 100 times with PBS, and homogenized using a vortex mixer. The NTA was performed using Nanosight NS300 (Malvern Instruments). The data was analyzed using NTA 2.3 Analytical Software (Malvern Instruments).
Induction of macrophage polarization
THP-1 cells were treated with 100 ng/ml phorbol 12-myristate 13-acetate (PMA) for 48 h to induce differentiation into M0-type macrophages. M0 macrophages were seeded in 6-well plates with complete growth medium, and conditioned medium or exosomes collected from MDA-MB-231 cells were added to PMA-treated THP-1 cells. After 48 h of incubation, THP-1 cells were collected for the detection of M1 and M2 markers using PCR.
For the coculture assay, PMA-treated THP-1 cells were seeded into the lower chamber of the Transwell, whereas MDA-MB-231 cells were seeded into the upper chamber, separated by a 3-μm insert. THP-1 cells were collected after 48 h for PCR analysis.
Statistical analysis
All data are expressed as the mean ± standard deviation (SD). The differences were analyzed using SPSS 19.0 software (IBM Co.). A normal distribution was tested using the Kolmogorov‒Smirnov test. A Student’s t-test or one-way ANOVA was used to compare the differences between groups with a normal distribution. For non-normal data, the comparisons were analyzed using a Wilcoxon-Mann‒Whitney test. A p < 0.05 indicated a significant difference.
RESULTS
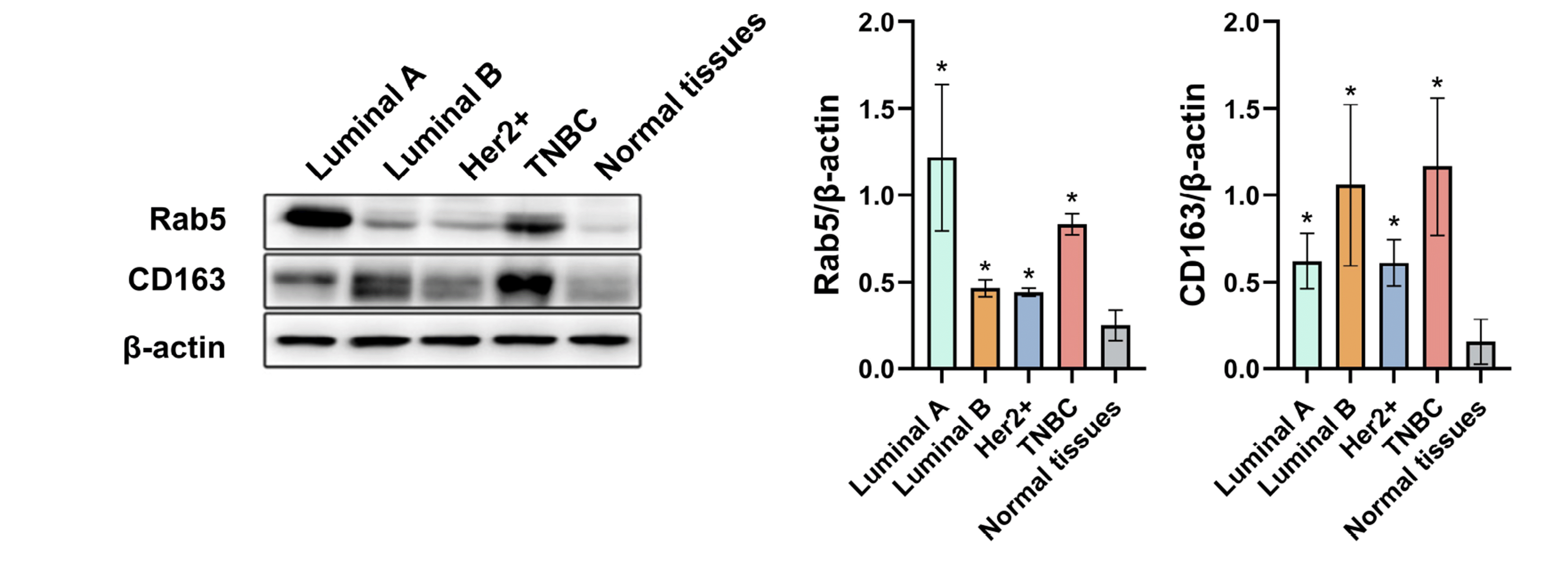
The expression of Rab5 and CD163 in breast cancer and normal tissues
To investigate the expression of Rab5 in breast cancer, we first detected its protein levels in normal breast tissue and different molecular subtypes of breast cancer, including luminal A, luminal B, Her-2-positive, and TNBC by Western blot. As indicated in Fig. 1, enhanced expression was observed in TNBC compared with normal tissues (p < 0.05), and luminal A showed the highest level of Rab5 among the four subtypes (p < 0.05). Because Rab5 is the primary participant in endosome transport, exosomes are believed to be derived from the endosomal system and play a vital role in intercellular communication [14,15], particularly for communication between cancer cells and tumor-associated macrophages [16]. We next tested the level of CD163, a marker for M2 macrophages, which are widely known as the protumor phenotype [17], the abnormal expression in tumor tissue usually suggests the evaluated infiltration of tumor-associated macrophages [18]. The highest level of CD163 was observed in TNBC and was over 7 times that in normal tissue (Fig. 1).
Establishment of Rab5-knockdown and overexpressed MDA-MB-231 cells
Given the overexpression of Rab5 in TNBC tissues, we explored the role of rab5 in TNBC. First, we generated three small interfering RNAs targeting human Rab5, and transfection was conducted in MDA-MB-231 cells. The fluorescence was evaluated 24 and 48 h post-transfection, and our results indicated that the optimal transfection conditions were 0.01 μM siRNA for 48 h (Fig. 2A). The knockdown efficiency of Rab5 was further determined by qRT-PCR, and the results suggested that siRNA#2 exhibited the greatest knockdown efficiency (Fig. 2B). In addition, the protein levels in MDA-MB-231 cells transfected with siRNA#2 as well as blank and negative control groups were evaluated using a Western blot (Fig. 2C), and siRNA#2 dramatically decreased the protein level of Rab5 (p < 0.05). Moreover, we overexpressed Rab5 in MDA-MB-231 cells using vector containing full-length human Rab5 cDNA, and the mRNA and protein levels indicated the success in building the overexpression cells (Fig. 2D, E).
Fig. 2
Knockdown of Rab5 in MDA-MB-231 cells by small interfering RNA.
(A) Effect of siRNA concentration and incubation time on transfected with siRNAs (n = 5), cells were stained with Block-iT Alexa Fluor Red, magnification 100×. (B) mRNA level of Rab5 in MDA-MB-231 cells transfected with siRNAs (n = 5). (C) Protein level of Rab5 in MDA-MB-231 cells transfected with siRNAs (n = 3). (D) mRNA and (E) protein levels of Rab5 in MDA-MB-231 cells transfected with empty or Rab5-containing vectors. Data was represented as mean ± SD. *p < 0.05 vs. control.
![]()
Rab5 regulated the migration and invasion of MDA-MB-231 cells
After the establishment of Rab5 knockdown MDA-MB-231 cells using siRNA, we next evaluated the role of Rab5 knockdown in cell migration and invasion. As suggested in Fig. 3A, Rab5-depleted MDA-MB-231 cells showed a reduced capacity for migration and invasion compared with the control groups (p < 0.05). In cells with Rab5 overexpression, the migration and invasion capacity were significantly enhanced (Fig. 3B, p < 0.05), when compared to the cells with empty vector transfection (Vector) and without transfection (Con).

Rab5 regulated the release of CD63 and CD9 proteins in MDA-MB-231 cells
To gain insight into whether the knockdown of Rab5 influences exosome secretion in MDA-MB-231 cells, we collected the supernatant of MDA-MB-231 cell culture medium and extracted the exosomes. Two hallmarks of exosomes, CD63 and CD9, were assessed by Western blot, and these two proteins were decreased in MDA-MB-231 cells with Rab5 depletion (Fig. 4A, p < 0.05). While in Rab5 overexpression cells, protein abundance of CD63 and CD9 were increased (Fig. 4B, p < 0.05). NTA was performed in MDA-MB-231 cell transfected with siNC or siRab5, and the results in Supplementary Fig. 1 demonstrated that Rab5 knockdown decreased the amount of exosomes (p < 0.05), while the diameter of exosomes remained unchanged (p > 0.05). The results indicated that Rab5 silencing inhibited whereas Rab5 overexpression promoted the secretion of exosomes in TNBC cells.
Rab5 may be implicated in the polarization of macrophages
Cancer cell-derived exosomes play an important role in the polarization of macrophages, which may determine the progression of tumors [19]. To uncover whether Rab5 in TNBC cells affects the polarization of macrophages, we incubated macrophages with conditioned medium or exosomes collected from MDA-MB-231 cells or cocultured macrophages with MDA-MB-231 cells (Fig. 5A). PMA-treated THP-1 cells (macrophages polarized to M0) were incubated with conditioned medium from MDA-MB-231 cells, and the levels of M2-macrophage, CD206, interleukin (IL)-10, and CD163 in THP-1 cells were dramatically increased when treated with the conditioned medium from siNC MDA-MB-231 cells compared with the mock group (THP-1 cells alone). Similar but lesser increases were observed in the levels of the M1-macrophage markers IL-1β and TNF-α, but no change was observed in iNOS. When THP-1 cells were treated with conditioned medium from siRab5 MDA-MB-231 cells, the three M1-macrophage markers were all increased, but the M2-macrophage markers, except for IL-10, remained unchanged (Fig. 5B). We further tested the polarization of THP-1 cells using exosomes derived from MDA-MB-231 cells or a coculture of THP-1 and MDA-MB-231 cells together, and similar results were observed (Fig. 5C, D). siRab5 knockdown in MDA-MB-231 cells tended to induce the M1 polarization of THP-1 cells through exosome-dependent mechanisms. In addition, we cocultured THP-1 cells with MDA-MB-231 cells in which Rab5 was overexpressed. M2-macrophage markers, CD206 and CD163, were observed increased in THP-1 cells cocultured with Rab5 overexpression MDA-MB-231 cells (Fig. 5E), when compared to those cocultured with un-transfected MDA-MB-231 cells. While for IL-10, coculture with Rab5 overexpression MDA-MB-231 cells led to a slight decrease in THP-1 cells. The expression of TNF-α was slightly decreased in Rab5-overexpressed MDA-MB-231 cell cocultured THP-1 cells, however, iNOS and IL-1β were not altered, when compared to those cocultured with normal MDA-MB-231 cells (Fig. 5E). These results suggest the involvement of Rab5 in the polarization of macrophages.
Fig. 5
The implication of Rab5 in the polarization of macrophages.
(A) Human THP-1 macrophages were treated with conditioned medium, exosomes collected from MDA-MB-231 cells, or were cocultured with MDA-MB-231 cells. The M2 (CD206, IL-10, and CD163) and M1 (iNOS, IL-1β, and TNF-α) markers in THP-1 cells treated with conditioned medium (B) or exosomes (C) or cocultured with MDA-MB-231 cells (D) were determined by qRT-PCR. (E) The expression of M1/M2 macrophage markers in THP-1 cells cocultured with MDA-MB-231 cells with Rab5 overexpression. The mock treatment represents THP-1 cells treated with normal medium alone. Data was represented as mean ± SD. *p < 0.05, **p < 0.001, ***p < 0.0001 vs. mock treatment.
![]()
DISCUSSION
The incidence of breast cancer is increasing worldwide, and increasingly, patients are younger [20]. As a molecular subtype, TNBC is negative for estrogen receptor, progesterone receptor, and HER-2. In addition, the characteristics of the clinical process, pathological morphology and molecular spectrum are highly heterogeneous among TNBCs [21]. TNBC is characterized by early onset, aggressiveness and metastasis, and poor clinical prognosis, accounting for 15%–20% of new breast cancer [22]. TNBC patients are highly variable in their response to standard chemotherapy. Currently, the lack of universal targeted therapy for TNBC is a great challenge in clinical practice.
Rab5 enhances the communication between cancer cells and macrophages by influencing the secretion of exosomes and facilitates the occurrence and development of breast cancer
This study demonstrated that the expression of Rab5 and the M2-like macrophage marker CD163 in breast cancer tissues were significantly higher than those in normal breast tissues, particularly in TNBC. These sresults suggest that the abnormal expression of Rab5 and the presence of M2-like macrophages in the microenvironment are closely related to the biological characteristics of aggressiveness and metastasis in TNBC, which is consistent with a previous investigation by Ivan and colleague [23] that showed that patients with breast cancer are more susceptible to relapse and metastasis and have a poor prognosis. Additionally, high expression of Rab5 was reported to be closely related to the high metastatic characteristics of other tumors [24-26]. It has been reported that CD163 is absent in breast cancer cell lines but is frequently present in breast cancer tissues [27]. We speculated that this difference may be due to communication between cancer cells and macrophages in the tumor microenvironment. Such communication is largely accomplished by the delivery of exosomes containing DNA, mRNA, and metabolic intermediates [28].
Rab5 protein is a key regulator during exosome biogenesis [29]. Two recent reports by Gorji-bahri and colleagues [30,31], however, appear to describe a contradictory correlation between Rab5 and CD63 in hepatocellular carcinoma, but suggested the importance of Rab5 in exosome secretion. Emerging evidence has revealed that exosomes derived from tumor cells can stimulate macrophages to transform into an M2-like phenotype, which supports tumor development [32-34].
For example, Ham and colleagues demonstrated that IL-6 receptor beta (glycoprotein 130, gp130) is included in breast cancer cell-derived exosomes, which are transported into macrophages and subsequently promote macrophage IL-6 secretion and M2 polarization through the STAT3 signaling pathway [35]. M2-like macrophages play vital roles in facilitating tumor cell growth by enhancing angiogenesis, remodeling the matrix, and secreting anti-inflammatory cytokines [36] as well as via immunosuppression and assisting tumor cells in escaping immune surveillance [37]. In addition, genetic exchange between tumor-associated macrophages and tumor cells could be another mechanism by which M2-like macrophages promote tumorigenesis [38,39]. Intriguingly, breast cancer patients with cancer cells expressing macrophage antigen CD163 are generally vulnerable to distant metastases and short survival [23]. We are interested in the possibility that Rab5 contributes to the occurrence and development of breast cancer by educating macrophages in an exosome-dependent manner.
Rab5 protein plays an important role in the regulation of migration, invasion and exosome secretion of triple-negative breast cancer cells
To test our abovementioned hypothesis, loss-of and gain-of-function of Rab5 in MDA-MB-231 cells was carried out by knockdown or overexpression, and our results implicated Rab5 in the migration, invasion and exosome secretion of TNBC cells. The aggressiveness, ease of metastasis and recurrence of TNBC with ectopic Rab5 expression might be attributed to 1) forced Rab5 expression facilitating the exosome secretion of cancer cells, thus enhancing communication between cancer cells and tumor-associated macrophages, the latter contributing to the protumor microenvironment; 2) high Rab5 expression leading to focal adhesion disassembly, which is conducive to the migration and invasion of TNBC cells [13]; or 3) Rab5 promoting the endo/exocytic cycle of membrane-type 1 matrix metalloprotease and β3 integrin cargos in cancer cells, which is necessary for MET signaling to drive invasion [12].
In conclusion, this study demonstrated high expression of Rab5 in TNBC and its potential correlation with aggressiveness, ease of metastasis and recurrence of TNBC. Therefore, Rab5 is expected to be a potential therapeutic target in triple-negative breast cancer.
SUPPLEMENTARY MATERIALS
Supplementary data including one figure can be found with this article online at https://doi.org/10.4196/kjpp.2023.27.2.157.
 XML Download
XML Download