

 PDF
PDF Citation
Citation Print
Print



Introduction
Cancer stem cells (CSCs) have been identified in various cancers, including liver cancer [1-3]. Owing to their biological characteristics, including chemoresistance, radio resistance, dormancy, and metastatic potential, they have drawn a lot of attention from the research field. Although chemotherapeutics can successfully reduce tumor volume, chemoresistant CSCs can survive and cause cancer relapse. Since the initial report of liver CSCs (LCSCs), many surface markers (epithelial cell adhesion molecule [EpCAM], leucine-rich repeat-containing G protein-coupled receptor 5 [LGR5], cluster of differentiation 133 [CD133], CD44, CD24, CD90, etc.) have been reported [2, 3]. In addition to identifying LCSC surface markers, the molecular maintenance mechanism of their stemness has also been explored. Signaling pathways, including WNT, hedgehog, and Notch pathways, and transcription factors, including Nanog, Oct4, Sox2, and Myc, have been reported to be important for the maintenance of stemness in CSCs. Based on these advances in CSC research, some of them have been applied to the development of therapeutics.
Liver cancer is the third most common cause of cancer-related death worldwide (GLOBOCAN 2020). It is the sixth most common type of cancer. The mortality rate has not improved over the last three decades. Notably, its incidence is expected to increase by approximately 60% between 2020 and 2040 in Asia. Hepatocellular carcinoma (HCC) is the most common form of primary liver cancer. Therefore, new insights into the mechanisms underlying HCC are required for the development of diagnostics and therapeutics for liver cancer.
Although therapeutics targeting CSC surface markers have been developed, their use frequently encounters new resistance, partly owing to the flexible hierarchy of CSCs [1]. New CSCs are generated from other daughter cells under inflammatory conditions, although CSCs that express specific CSC markers are eliminated by therapeutics. Another cause for the resistance in applying the therapeutics is the heterogeneity of CSCs. Single-cell transcriptomics and reports of many types of surface markers suggest the heterogeneity of CSCs. The CSC hypothesis is based on the assumption that these cells have the same genetic background as their daughter cells, although they are epigenetically different from each other. However, cancer cells within a tumor can have different genetic backgrounds [4]. Therefore the genetic heterogeneity of LCSCs needs to be examined in detail.
In this study, to reveal genetic heterogeneity of LCSCs, we examined the genetic mutation status of LCSCs based on their surface markers (CD133, CD44, CD24, CD47, CD13, CD90, ICAM1, EpCAM, LGR5, keratin 19 [KRT19]). By analyzing Exome-Seq and RNA-Seq data of HCC (n=366) in the Cancer Genome Atlas (TCGA) database, we found that LCSCs are genetically heterogenous depending on their surface markers and some mutations are associated with a specific surface marker of LCSCs.
Go to : 
Materials and Methods
RNA-Seq and Exome-Seq data from TCGA were downloaded from the c-BioPortal. The TCGA cohort includes genomic information of 366 HCC patients. RNA and DNA were extracted from tumor and adjacent normal tissue specimens using a modification of the DNA/RNA AllPrep kit (QIAGEN, not single cell-Seq). The expression data for mRNA were batch-corrected to adjust for platform differences between the GAII and HiSeq Illumina sequencers. Somatic exome variant analysis was performed to remove potential germline calls as well as non-exonic variants. RNA-Seq data of TCGA were further classified based on the expression level (RNA-seq value) of each CSC marker. The high or low group based on each marker’s expression level was defined as a higher 25% or lower 25% of patients among HCC patients in the TCGA HCC cohort. The intermediate group was defined as 25%–75% of patients. The genes which were harboring mutations were further selected based on their relationship with cancer, whether they were cancer genes (OncoKB), and the frequency of HCC (>1.0%). Somatic exome mutations for each gene were further selected by SIFT (deleterious group) or PolyPhen score (probably damaging group), which predicts significant functional changes.
The correlation between the expression level of each cancer stem marker and the mutation of each cancer gene was evaluated using the chi-square test. Results with a P-value of <0.05 were regarded as statistically significant. Data were analyzed using IBM SPSS Statistics for Windows, version 25.0 (IBM Corp., Armonk, NY, USA; RRID: SCR_002865).
Go to :
Results and Discussion
CD133 (PROM1)
CD133 is a five-transmembrane single-chain glycoprotein that is usually localized in membrane protrusions [5-8]. It may function as an organizer of cellular protrusion and regulate the number of microvilli and the structure of filopodia. Since its discovery in hematopoietic stem cells [5], its expression in other adult stem cells has been reported [9]. Its biological roles in proliferation, migration, metastasis, spheroid formation, and in vivo tumorigenicity of liver cancer cells have been reported [10-14]. Interestingly, its regulatory roles in angiogenesis, autophagy, metabolic reprogramming, radio resistance, and chemoresistance have also been reported [15-21]. Its high expression is associated with poor survival in colon, lung, and liver cancers [22-25]. Its potential as a surface marker for CSCs has been suggested in lung, stomach, pancreas, colon, and liver cancers [25]. Moreover, many signaling pathways, including AKT, IL8-CXCL1, JNK, and NFκB, have been associated with CD133-positive cancer cells [12, 15, 26, 27]. Recently, CD133-targeted therapeutics, including antibodies, aptamers, T-cell therapies, viruses, and compounds, have been actively examined [28-32].
When we compared CD133-high HCC tissues with CD133-low tissues, we found that many kinds of cancer gene mutations were enriched in the CD133-high group (Fig. 1A, Table 1). Mutations in IRF2, TRRAP, and ASXL2 were not observed in the CD133-low group. Notably, the frequency of BAP1 mutations correlated with the expression levels of CD133. In addition to the BAP1 mutation, the mutation frequency of FGFR2, TLE4, MECOM, PBRM1, NOTCH1, and ERBB3 was at least three-fold higher in the CD133-high group than in the CD133-low group.
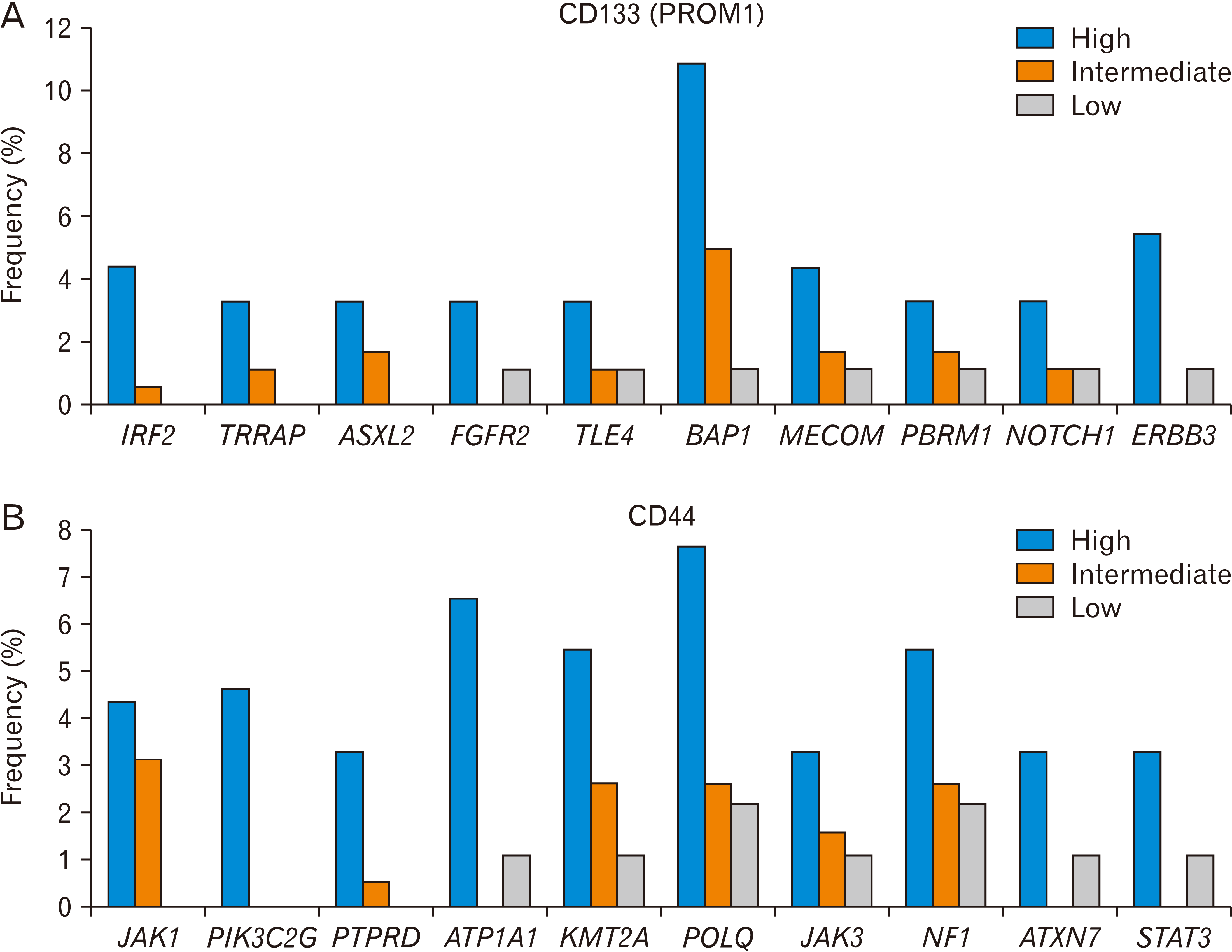 | Fig. 1Gene mutations associated with CD133 (A) or CD44 (B) expression level in liver cancer (TCGA, n=366). The frequency is based on the number of patients who harbor a specific gene mutation and is shown to depend on the level of CD133 or CD44 (high, intermediate, and low). The Ki-square analysis showed a significant association between the CD133 or CD44 expression level, and the gene mutations. CD, cluster of differentiation; TCGA, The Cancer Genome Atlas.
|
Table 1
Summary of frequently mutated genes in each LCSC group
![]()
The association of some mutations in these cancer genes with CD133-positive CSCs has been reported. BAP1 mutation is associated with the expression of stemness genes, including EpCAM and PROM1, and with their aggressiveness in liver cancer [33]. FGFR2-mediated signaling regulates the survival and proliferation of murine hepatoblasts and liver CSCs [34]. IRF2 knockdown has been associated with the chemoresistance of CD133-positive colon CSCs, in which IRF2 represses the promoter activity of PTPN13 leading to decreased expression of FAP1 [35]. Chemoresistance of colon CSCs is influenced by FAP1. The PBAF/PBRM1 pathway increased the expression of CD133 in prostate CSCs [36]. Notch1 regulated CD133-positive cancer stem-mediated melanoma growth [37] and directly induced CD133 expression in gastric cancer [38]. ERBB3 expression has been observed in CD133-positive glioblastoma stem cells [39], and ERBB3 targeting inhibited glioblastoma [40].
CD44
CD44 is a lymphocyte-homing receptor that interacts with the extracellular matrix, cell-cell interaction, adhesion, and migration [41-43]. It also functions as a glycoprotein receptor that interacts with various molecules, including hyaluronic acid, osteopontin, chondroitin, collagen, fibronectin, and metalloproteinases [44-48]. Notably, alternatively spliced variants are critical for cancer progression [49]. Moreover, its intracellular domain interacts with RNUX2 and regulates the transcription of MMP9 [50]. It regulates the migration, invasion, and metastasis of cancers and is associated with poor survival of cancer patients [49, 51]. Its potential as a surface marker for CSC has been suggested in various cancers, including colorectal, pancreatic, gastric, hepatocellular, and breast cancer [49]. The AKT, FoxM1, YAP/TEAD, and TGF-β signaling pathways have been associated with CD44-expressing cancer cells [52-55].
When we compared CD44-high HCC tissues with CD44-low tissues, we found that many kinds of cancer gene mutations were enriched in the CD44-high group (Fig. 1B, Table 1). Mutations in JAK1, PIK3C2G, and PTPRD were not observed in the CD44-low group. Notably, the frequency of POLQ mutations correlated with the expression level of CD44. In addition to POLQ mutations, the mutation frequency of ATP1A1, KMT2A, JAK3, NF1, ATXN7, or STAT3 was higher in the CD44-high group than in the CD44-low group by at least three-fold.
The association of some mutations in these cancer genes with CD44 expression has been reported. In addition to multiple myeloma [52, 56], the JAK-STAT signaling pathway regulates the expression of CD44. The interaction of STAT signaling and CD44 has been reported in breast cancer and ovarian CSCs [57, 58]. The inhibition of PIK3C2G inhibited the growth of breast CSCs [59]. The cooperation of PTPRD with CD44 for migration and progression has been reported in colon cancer, and its possible application to liver cancer has also been suggested [60, 61]. A KMT2A rearrangement was associated with CD44 in acute leukemia [62, 63]. The NF1-RAS pathway regulated mesenchymal transformation, leading to increased expression of CD44 in glioblastoma [64].
CD24
CD24 is a mucin-like glycoprotein that regulates the growth and differentiation of B-lymphocytes, neutrophils, and neuroblasts [65, 66]. It has been suggested to act as a brake on the immune system and as an antiphagocytic surface protein [67]. It has been reported to be expressed in ovarian cancer, breast cancer, non-small cell lung cancer, prostate cancer, pancreatic cancer, and HCC [68, 69]. It regulates proliferation, migration, and invasion of cancer cells [66, 68, 70]. It has been suggested as a surface marker for CSC in breast cancer, gastric cancer, cervical cancer, multiple myeloma, cholangiocarcinoma, and HCC and regulates the differentiation and metastasis of liver cancer cells [71-76]. Moreover, its expression was associated with a poor prognosis in patients with liver cancer. Its association with the STAT3-NANOG pathway has been reported [77].
When we compared CD24-high HCC tissues with CD24-low tissues, we found that many types of cancer gene mutations were enriched in the CD24-high group (Fig. 2A, Table 1). Mutations in CDH1, ERBB3, FLT1, and HIRA were not observed in the CD24-low group. Notably, the frequency of BAP1 mutations correlated with the expression level of CD24. In addition to the BAP1 mutation, the mutation frequency of IDH1, CLTCL1, NFATC2, HIP1 and CPS1 was higher in the CD24-high group than in the CD24-low group by at least three-fold.
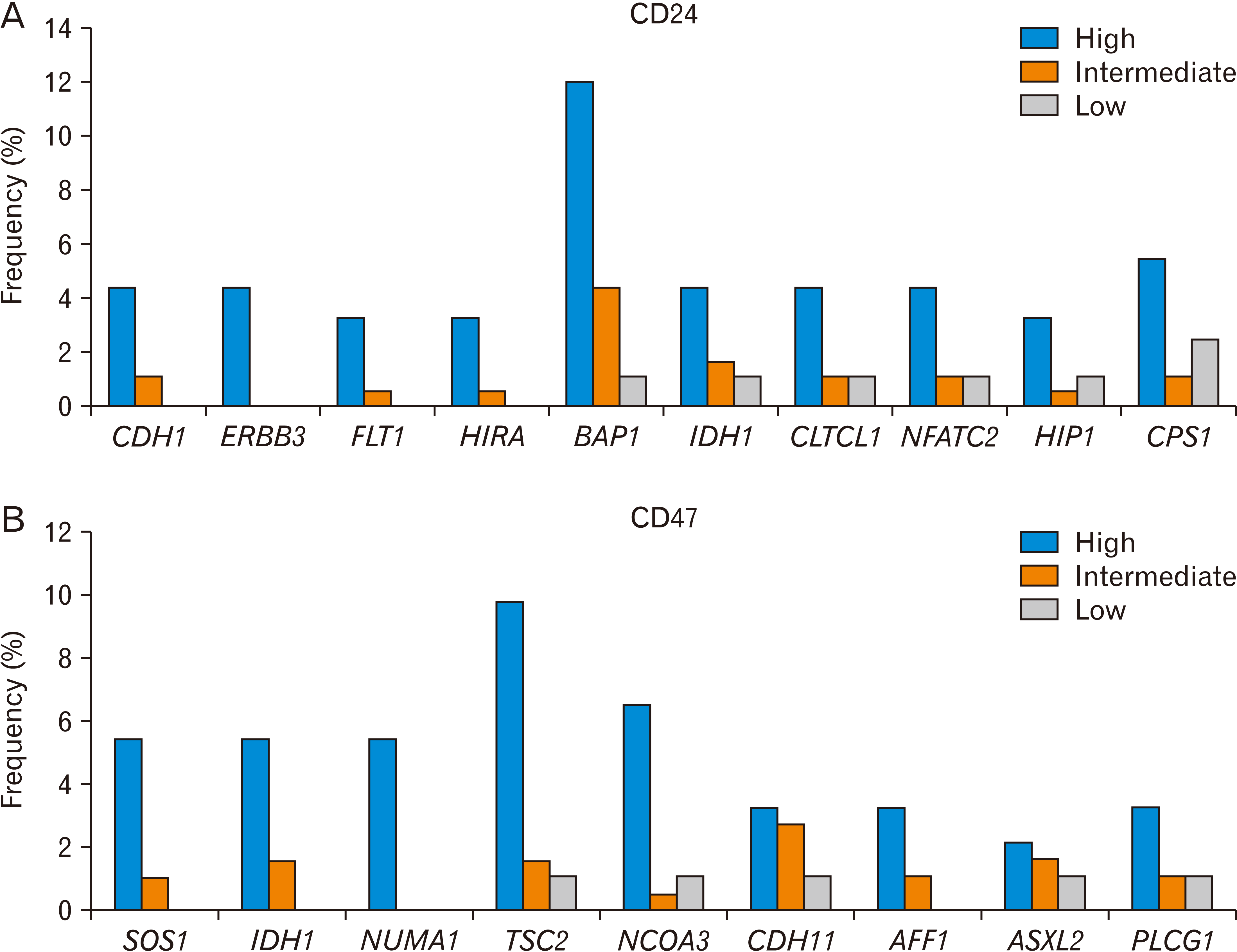 | Fig. 2Gene mutations associated with CD24 (A) or CD47 (B) expression level in liver cancer (TCGA, n=366). The frequency is based on the number of patients who harbor a specific gene mutation and is shown to depend on the level of CD24 or CD47 (high, intermediate, and low). The Ki-square analysis showed a significant association between the CD24 or CD47 expression level, and the gene mutations. CD, cluster of differentiation; TCGA, The Cancer Genome Atlas.
|
The association of some mutations in these cancer genes with CD24 expression has been reported. CDH1 knockdown has been associated with the enrichment of CD24-positive colon CSCs [78]. ERBB2/3 contributes to the conversion of induced pluripotent stem cells into CSCs. Hypoxia activates HIF, resulting in the upregulation of the “do not-eat-me” signal surface markers (CD24 and CD47) and vascular endothelial growth factor, which bind to FLT1 in liver cancer [79]. IDH mutation induced phenotypic reprogramming in glioma, resulting in the generation of CSCs and an increase in CD24-positive cells [80, 81].
CD47
CD47 is a transmembrane protein that belongs to the immunoglobulin superfamily and forms supramolecular complexes with integrins, G proteins, and cholesterol [82, 83]. It interacts with signal-regulatory protein α (SIRPα), thrombospondin 1, and integrins, and is involved in proliferation, migration, phagocytosis, apoptosis, and immune homeostasis. Its overexpression has been frequently observed in various cancer cells, including myeloma, leiomyosarcoma, acute lymphocytic leukemia, non-Hodgkin’s lymphoma, breast cancer, osteosarcoma, head and neck squamous cell carcinoma, and liver cancer [82]. It inhibits macrophage-mediated phagocytosis by interacting with SIRPα [84, 85]. It has also been found to be expressed in CSCs in leukemia, glioma, pancreatic cancer, and liver cancer [86-89]. It regulates self-renewal, metastasis, and chemoresistance in LCSCs [87]. It is a poor prognostic factor in liver cancer and is associated with the cathepsin S (CTSS)-protease-activated receptor 2 signaling pathway by preferentially secreting CTSS [87]. Therapeutics targeting CD47 have been actively investigated [90-93].
When we compared CD47-high HCC tissues with CD47-low tissues, we found that many kinds of cancer gene mutations were enriched in the CD47-high group (Fig. 2B, Table 1). Mutations in SOS1, IDH1, and NUMA1 were not observed in the CD47-low group. Notably, the frequency of the TSC2 mutation correlated with the expression level of CD47. In addition to the TSC2 mutation, the mutation frequencies of NCOA3, PTPRB, CDH11, and JAK1 were at least three-fold higher in the CD47-high group than in the CD47-low group. Interestingly, PLCG1 was overexpressed in chronic lymphocytic leukemia cells and CD47 was a good therapeutic target in these cells [94].
CD13 (ANPEP)
CD13 is a zinc-dependent type II exopeptidase located in the plasma membrane and engaged in the post-secretory processing of secreted signaling peptides and their binding to their receptors [95]. It is expressed in the kidney, intestine, liver, and central nervous system. It regulates proliferation, invasion, angiogenesis, chemoresistance, and radio resistance of cancer cells [96-98]. Its expression has been associated with poor prognosis in various cancers, including pancreatic and colon cancers, non-small cell lung cancer, malignant pleural mesothelioma, hepatoblastoma, and soft tissue sarcoma [97, 99-103]. It has been suggested to be a surface marker for CSC in liver cancer and is associated with the TGF-β signaling pathway [104-106].
When we compared CD13-high HCC tissues with CD13-low tissues, we found that many kinds of cancer gene mutations were enriched in the CD13-high group (Fig. 3A, Table 1). Mutations in BRCA2, NSD1, KDM4C, BAX, and HSP90AA1 were not observed in the CD13-low group. Notably, the frequency of BAP1 and RB1 mutations correlated with the expression level of CD13. In addition, the mutation frequency of PTPN13, TEK, MYH11, KDR, and RAD50 was at least three-fold higher in the CD13-high group than in the CD13-low group.
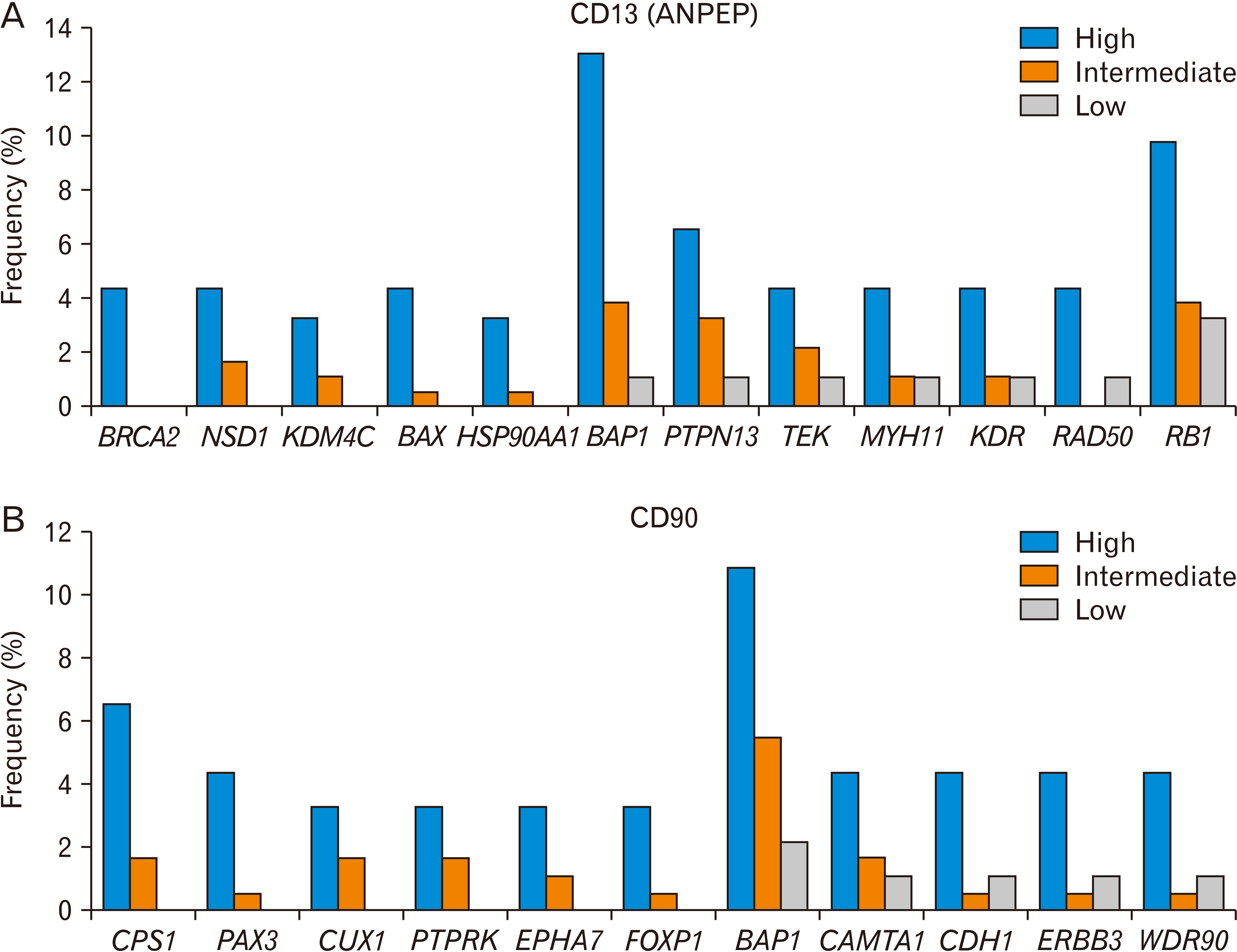 | Fig. 3Gene mutations associated with CD13 (A) or CD90 (B) expression level in liver cancer (TCGA, n=366). The frequency is based on the number of patients who harbor a specific gene mutation and is shown to depend on the level of CD13 or CD90 (high, intermediate, and low). The Ki-square analysis showed a significant association between the CD13 or CD90 expression level, and the gene mutations. CD, cluster of differentiation; TCGA, The Cancer Genome Atlas.
|
The association of some mutations in these cancer genes with CD13 expression has been reported. Overexpression of KDM4C was observed in CD13-positive LCSCs [107]. Its depletion decreased tumor initiation, as examined by sphere formation and xenograft assays. Angiopoietin increased the expression of CD13 via the TEK (TIE2) receptor in pericytes [108].
CD90 (THY1)
CD90 is a glycosylphosphatidylinositol-anchored glycoprotein expressed in thymocytes, neurons, mesenchymal stem cells, hepatic stem cells, natural killer cells, T cells, and endothelial cells [109, 110]. It is involved in cell-cell and cell-matrix interactions, apoptosis, and migration. It can promote tumorigenesis, metastasis, and chemoresistance and is a prognostic factor [111]. It has been suggested as a surface marker for CSC in HCC that does not express EpCAM [112, 113]. IL6, hedgehog, and AKT signaling pathways have been associated with CD90 [114, 115].
When we compared CD90-high HCC tissues with CD90-low tissues, we found that many types of cancer gene mutations were enriched in the CD90-high group (Fig. 3B, Table 1). Mutations in CPS1, PAX3, CUX1, EPHA7, and FOXP1 were not observed in the CD90-low group. Notably, the frequency of BAP1 mutations correlated with the expression level of CD90. In addition, the mutation frequencies of CAMTA1, CDH1, ERBB3 and WDR90 were at least three-fold higher in the CD90-high group than in the CD90-low group. A previous study reported that increased FOXP1 expression was observed in CD90-positive hematopoietic stem cells, which contributed to leukemic cell growth [116].
ICAM1
ICAM1 is a cell surface glycoprotein and an adhesion receptor in various cell types, including immune, endothelial, and epithelial cells [117]. It is a ligand for the leukocyte adhesion protein LFA1. It regulates the metastasis and tumorigenic potential of liver cancer cells and is a marker for CSC [118, 119]. It is a poor prognostic factor for liver cancer and is associated with Nanog expression [119].
When we compared ICAM1-high HCC tissues with ICAM1-low tissues, we found that many kinds of cancer gene mutations were enriched in the ICAM1-high group (Fig. 4A, Table 1). Mutations in TSC2, CDH11, and TLX3 were not observed in the ICAM1-low group. The mutation frequency of SETD2, NFATC2, CHD4, NOTCH3, JARID2, KMT2C, and ASLX2 was at least three-fold higher in the ICAM1-high group than in the ICAM1-low group.
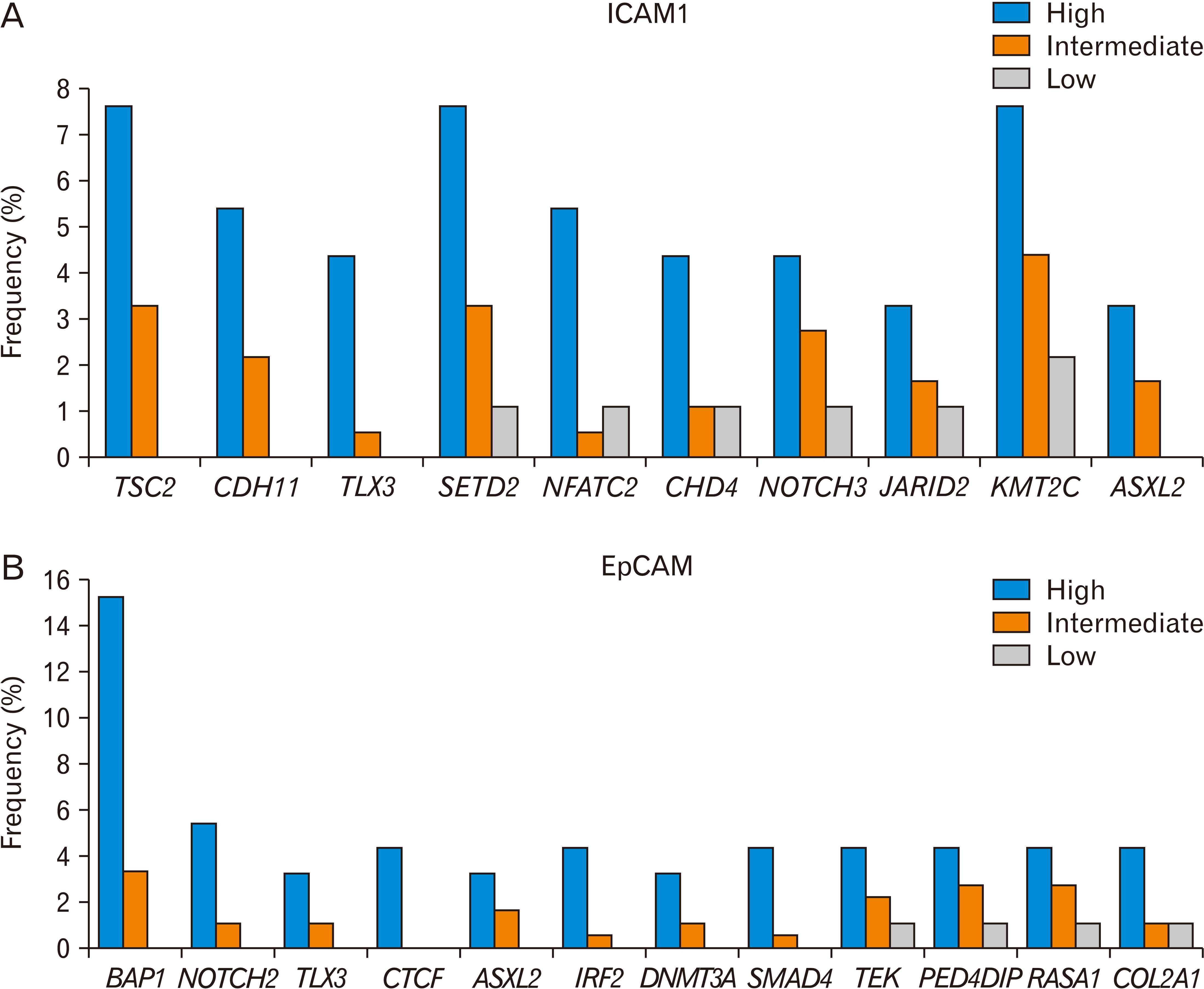 | Fig. 4Gene mutations associated with ICAM1 (A) or EpCAM (B) expression level in liver cancer (TCGA, n=366). The frequency is based on the number of patients who harbor a specific gene mutation and is shown to depend on the level of ICAM1 or EpCAM (high, intermediate, and low). The Ki-square analysis showed a significant association between the ICAM1 or EpCAM expression level, and the gene mutations. ICAM1, Intercellular adhesion molecule 1; EpCAM, epithelial cell adhesion molecule; TCGA, The Cancer Genome Atlas.
|
The association of some mutations in these cancer genes with ICAM1 expression has been reported. NFAT belongs to the Rel homology domain-containing family of transcription factors and can recognize DNA sequences that can be recognized by NF-κB [120, 121]. NFATC2 siRNA decreased TNFα-induced ICAM1 expression and cell adhesion in human retinal microvascular endothelial cells [122]. TGF-β1 increased Notch3 and ICAM1 expression levels in hepatic stellate cells, and the Notch pathway was important in TGF-β1 induced activation of hepatic stellate cells [123].
EpCAM
EpCAM is a type I membrane protein containing two EGF-like domains [124, 125]. Its expression has been reported in various types of carcinomas. It regulates the self-renewal and tumorigenesis of cancer cells and is a surface marker for CSC [126]. WNT, CHD4, and OSM signaling pathways have been linked to EpCAM-positive liver cancer cells [127-129].
When we compared EpCAM-high HCC tissues with EpCAM-low tissues, we found that many kinds of cancer gene mutations were enriched in the EpCAM-high group (Fig. 4B, Table 1). Mutations in BAP1, NOTCH2, TLX3, CTCF, ASXL2, IRF2, DNMT3A and SMAD4 were not observed in the EpCAM-low group. Notably, the frequency of the BAP1 mutation correlated with the expression level of EpCAM. In addition, the mutation frequency of TEK, PED4DIP, RASA1, and COL2A1 was at least three-fold higher in the EpCAM-high group than in the EpCAM-low group.
The association of some mutations in these cancer genes with EpCAM expression has been reported. BAP1 knockdown is associated with EpCAM overexpression in HCC [33]. Moreover, BAP1 knockout in human liver organoids also resulted in overexpression. NOTCH2 knockdown downregulated EpCAM expression [130]. Inhibition of Notch signaling reduced EpCAM expression [131]. IRF2 regulated the stemness of intestinal stem cells, and IRF2 deletion impaired regeneration of the colon epithelium [132]. Displacement of DNMT3A and DNMT3B by DNMT3L contributed to the overexpression of EpCAM in HCCs [133]. The TGF signaling pathway regulated EpCAM expression in liver cancer [134].
LGR5
LGR5 is a G-protein-coupled receptor that contains seven transmembrane domains [135]. It can bind to R-spondin 1–4 proteins and associate with phosphorylated LRP6 and frizzled receptors. It is expressed in stem cells of the intestine, ovary, hair follicle, mammary gland, and stomach [136-139]. It regulates regeneration of the intestine, colon, liver, pancreas, and stomach [140-142]. In liver cancer cells, it regulates tumorigenic potential, chemoresistance, and migration [143]. LGR5-positive liver cancer cells have been associated with the HGF, LSD1, Prickle, and WNT signaling pathways [144, 145].
When we compared LGR5-high HCC tissues with LGR5-low tissues, we found that many types of cancer gene mutations were enriched in the LGR5-high group (Fig. 5A, Table 1). Mutations in EP300, DNMT3A, GMPS, and EPHA5 were not observed in the LGR5-low group. Notably, the frequency of CTNNB1 mutations correlated with the expression level of LGR5. In addition, the mutation frequency of RELN, IL6ST, ROBO1, KIT, and COL2A1 was at least three-fold higher in the LGR5-high group than in the LGR5-low group.
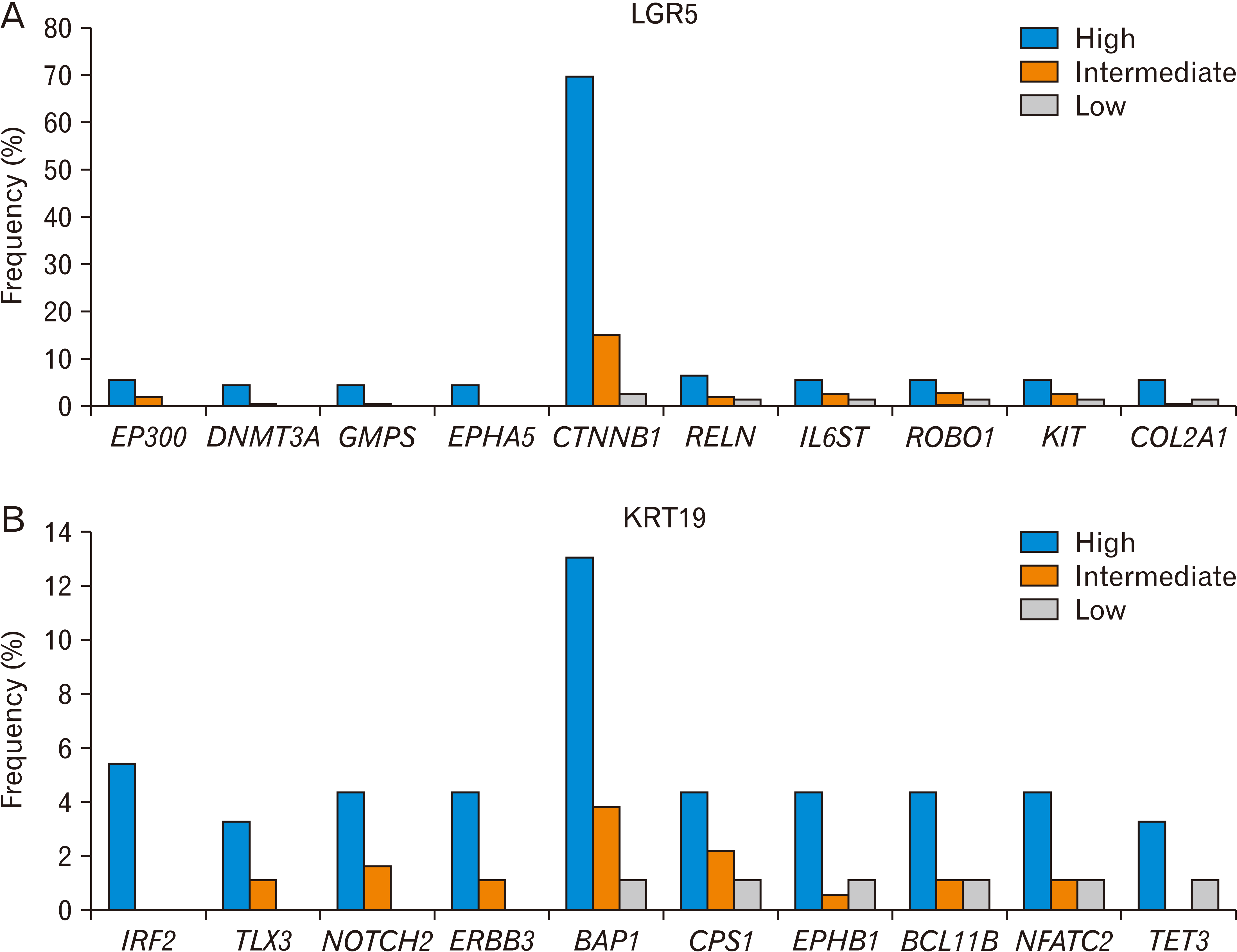 | Fig. 5Gene mutations associated with LGR5 (A) or KRT19 (B) expression level in liver cancer (TCGA, n=366). The frequency is based on the number of patients who harbor a specific gene mutation and is shown to depend on the level of LGR5 or KRT19 (high, intermediate and low). The Ki-square analysis showed a significant association between the LGR5 or KRT expression level, and the gene mutations. LGR5, leucine-rich repeat-containing G protein-coupled receptor 5; KRT19, keratin 19; TCGA, The Cancer Genome Atlas.
|
The association of some mutations in these cancer genes with LGR5 expression has been reported. LGR5 is a target gene of the WNT signaling pathway [146]. Therefore, the CTNNB1 mutation is associated with LGR5 expression in various cancer cells. EP300 modifies CTNNB1 and regulates the interaction between CTNNB1 and TCF4 [147]. DNMT3A expression was observed in LGR5-positive colon CSCs, and its deletion inhibited intestinal tumor development [148]. The YAP-IL6ST loop increased the expression of LGR5 during the progression of colon cancer progression [149]. Slit2 overexpression maintained LGR5+ stem cell proliferation in the intestinal crypt, and Robo1/2 partial knockout reduced the number of LGR5-positive stem cells [150]. cKit-positive cells in the mouse colon promote organoid formation via LGR5-positive stem cells [151].
KRT19
KRT19 is an intermediate filament protein involved in myofiber organization [152]. It is expressed in the bipotential cells of the liver and regulates fluorine-18 deoxyglucose accumulation [153]. It regulates proliferation, chemoresistance, tumorigenicity, invasion, and metastasis of liver cancer cells [154]. It has been reported to be a poor prognostic factor [155, 156]. TGF-β, PDGFR, and HGF signaling pathways have been associated with KRT19-positive liver cancer cells [153, 157, 158].
When we compared KRT19-high HCC tissues with KRT19-low tissues, we found that many types of cancer gene mutations were enriched in the KRT19-high group (Fig. 5B, Table 1). Mutations in IRF2, TLX3, NOTCH2, and ERBB3 were not observed in the KRT19-low group. Notably, the frequency of BAP1 mutations correlated with the expression level of KRT19. In addition, the mutation frequency of CPS1, EPHB1, BCL11B, NFATC2, and TET3 was at least three-fold higher in the KRT19-high group than in the KRT19-low group.
The association of some mutations in these cancer genes with KRT19 expression has been reported. Inhibition of Notch signaling reduced KRT19-positive cells during murine lacrimal gland formation [159]. Moreover, the expression of KRT19 in HCC cells was associated with the expression of NOTCH2 [154]. BAP1 knockdown was associated with the overexpression of KRT19 [33].
The heterogeneous composition of cancer cells in the tumor is a great challenge for oncologists because it is a critical cause of tumor relapse, chemoresistance, and metastasis. Genetic and epigenetic changes are the underlying mechanisms of cancer cell heterogeneity in tumors. In the present study, we examined the genetic mutation status of LCSCs and found genetic heterogeneity depending on markers for LCSCs (Table 1).
Different surface markers for LCSCs showed a unique pattern of propensity for genetic mutations (Table 1). CD133-positive cells showed frequent mutations in IRF2, BAP1, and ERBB3. However, LGR5-positive cells showed frequent mutations in CTNNB1, RELN, and ROBO1. These results suggest that targeting a specific surface marker cannot remove other types of CSCs that express different surface markers. Therefore, genetic classification of cancer is required for the development of stem cell therapies for cancer.
Notably, some genetic mutations were frequently observed irrespective of the surface markers of LCSCs (Table 1). BAP1 mutation was frequently observed in CD133-, CD24-, CD13-, CD90-, EpCAM- or KRT19-positive LCSCs. ASXL2 mutation was also frequently observed in CD133-, CD47-, ICAM1-, and EpCAM-positive LCSCs. Mutations in ERBB3, IRF2, TLX3, CPS1, and NFATC2 were observed in more than three types of LCSCs. Interestingly, some surface marker-positive cells showed common mutations. For example, ASXL2, BAP1, and IRF2 mutations were observed in both CD133-positive and EpCAM-positive cells. These results suggest that there are some common mechanisms for the development of LCSCs that need to be considered in the development of LCSC-targeting therapeutics.
In conclusion, based on above results we conclude that LCSCs are genetically heterogenous depending on their surface markers. In addition some mutations are frequently found in a LCSC group which expresses a specific surface marker. These results suggest that when therapeutics targeting LCSCs which express a specific marker, are considered for the treatment, mutational profiling of patients needs to be examined for the possibility of combination therapy. In addition, the existence or the new generation of other kinds of LCSCs needs to be considered for the treatment because the mutation status of patients is complex and always changing, which can lead to the new generation of other kinds of LCSCs.
Go to :
 XML Download
XML Download