

 PDF
PDF Citation
Citation Print
Print



Introduction
Skeletal muscles act to produce joint movements. Usually, to avoid friction against the hard tissue at and near the joint, a tendon across the joint is not accompanied by muscle fibers that extend distally beyond the joint. To increase the stability of the joint during muscle contraction, however, some muscle fibers are likely to cover or surround the joint capsule; for example the muscle fibers surrounding or along the rotator cuff of the glenohumeral joint. These shoulder muscle fibers are not tightly attached to the joint capsule but rather separated by the aponeurosis or cuff, thus avoiding mechanical injury. In contrast, the muscle bellies likely to issue distally-extending muscle fibers beyond the level of the wrist and ankle joints are generally undetermined with an exception. Some muscle fibers of the flexor digitorum superficialis (FDS) were found to be inserted into the carpal tunnel toward or across the wrist joint. This morphology is thought to be a pathologic cause of pain in patients with carpal tunnel syndrome [1, 2].
The topographical relationship between muscle belly and joint differs in fetuses and adults. The brachialis is a major muscle that attaches to and stabilizes the anterior aspect of the humeroulnar joint in adults [3, 4], whereas the flexor pollicis longus and others muscles cover the joint capsule in late-term fetuses [5]. In fetuses, the subscapularis muscle belly tends to extend into the glenohumeral joint cavity along the thick tendon [6]. Because the subscapularis tendon provides high resistance against mechanical stress in adults [7], the intra-articular muscle fibers should degenerate during shoulder movements in infants. These findings suggest the need to compare the morphology of distally-extending muscle fibers in fetuses and adults. In late-term fetuses, we hypothesized usual over-production of muscle bellies reaching or extending over the joint that the muscle acts on.
In the present study, distally-extending muscle fibers at the wrists and ankles of human late-term fetuses were examined histologically. Based on the anatomy of candidate muscles across these joints in fetuses, long tendons across the wrists and ankles were macroscopically evaluated in donated cadavers of elderly adults. Muscle fibers in fetuses were difficult to identify macroscopically because of the pale color, whereas it is hard to make histological sections of the adult specimen containing a joint because of the size and stiffness. Although only wrists and ankles were examined, the present study would reveal a specific type or site of muscle-tendon interface even in adults.
Go to : 
Materials and Methods
This study was performed in accordance with the provisions of the Declaration of Helsinki 1995 (as revised in 2013). The unilateral wrists and ankles of 16 late-term fetuses of gestational age 30–40 weeks and crown-rump length 255–325 mm had been cut into semiserial sections of 200–300 micron intervals and stained with hematoxylin and eosin, as described [5, 8]. The sectional plane was sagittal in four wrists and ten ankles, frontal in six wrists and horizontal in six wrists and six ankles.
All fetuses were part of the collection of the Department of Anatomy, Akita University and had been donated by their families to the Department in 1975–1985 and preserved in 10% w/w neutral formalin solution for more than 30 years. Data on these specimens included the date of donation and the number of gestational weeks, but did not include the name of the family, obstetrician or hospital or the reason for abortion. The use of these specimens for research was approved by the Akita University Ethics Committee (No. 1428). Dr. Hiroshi Abe, Emeritus Professor at Akita University, kindly permitted us to use these materials. Before routine procedures for embedding in paraffin, the fetal limb specimens were decalcified by incubation at room temperature in Plank-Rychlo solution (AlCl2/6H2O, 7.0 w/v%; HCl, 3.6 w/v%; HCOOH, 4.6 w/v%) for 3–7 days. Most photographs for histology were taken with a Nikon Eclipse 80 (Nikon, Tokyo, Japan), whereas photographs at ultra-low magnification (objective lens less than ×1) were obtained using a high-grade flat scanner with translucent illumination (Epson scanner GTX970; Epson, Tokyo, Japan).
Unilateral wrists and ankles were dissected from the cadavers of 15 elderly individuals, mean age 82 years (range, 75–97 years) at time of death. All cadavers had been donated to Tokyo Dental College for research and education on human anatomy, and had been fixed by arterial perfusion of 10% v/v formalin solution and stored in 50% v/v ethanol solution for more than 3 months. The use of cadavers for research was approved by the ethics committee of Tokyo Dental College (No. 932). The cause of death in all individuals was ischemic disease of the heart or brain. For dissection, the carpal tunnel and extensor tendon sheathes at the wrist was opened and the retinaculum at the ankle was removed to expose long tendons of the flexor and fibular muscles. After labeling the joint surfaces, photographs were taken of the wrist and ankle tendons. Deep flexor tendons were visualized after cutting and retraction of the superficial tendons.
Go to :
Results
Findings in late-term fetuses
Most or all of the late-term muscle bundles of the flexor digitorum profundus (FDP) and the FDS in the carpal tunnel extended distally beyond the radiocarpal joint in all six wrists and hands for horizontal sections and in six of the 10 for sagittal and frontal sections. In the FDS and FDP, the second to fifth tendons were likely to carry distally-extending muscle belly (Figs. 1, 2), with the FDS muscle fibers being more likely than the FDP muscle fibers to extend beyond the lunate to reach the levels of the capitate and hamate (Fig. 1A, B). In all six wrists and hands for horizontal wrist sections, the FDS muscle belly to the third finger and the FDP muscle belly to the fourth finger were found to carry over-extending muscle fibers beyond the radiocarpal joint cavity. These over-extending muscle bundles were attached to the posterior or ulnar aspect of the proper tendon (Fig. 2A, B). The distally-extending muscle fibers often formed a thin layer attached to the proper tendon. Examination of the frontal sections provided a great advantage, ensuring distances of ≥3 mm between the distally-extending FDP muscle belly and the lumbricalis origin from the FDP tendon. On the extensor side of the wrist, the extensor digitorum and extensor pollicis longus frequently carried distally-extending muscle fibers (Fig. 2C), but these fibers did not extend beyond the distal end of the radius. In these extensors, the distal end of the muscle belly was located at the ulnar edge of the tendon.
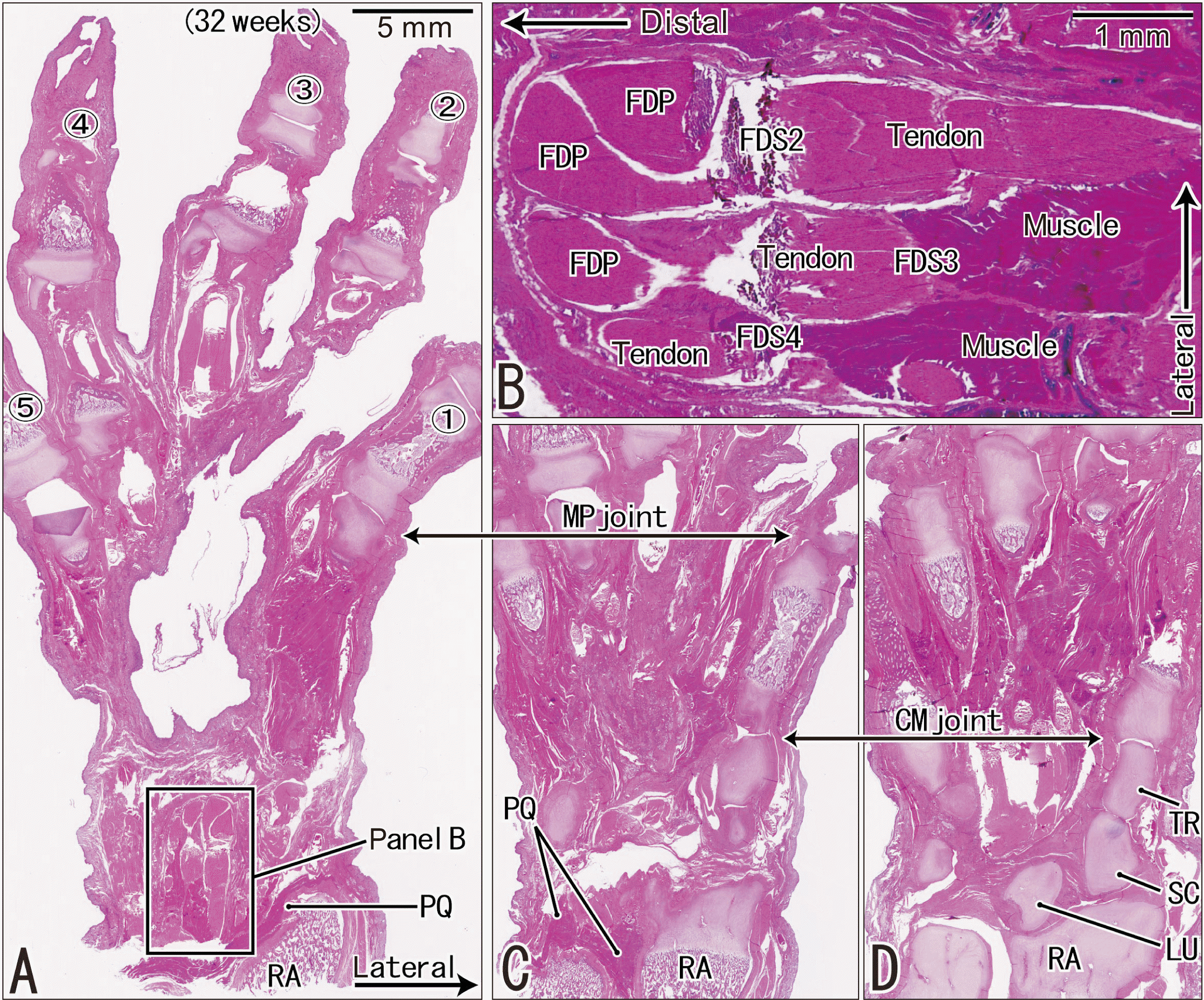 | Fig. 1Longitudinal sections of the hand and wrist showing distally extending muscle bellies of the flexor digitorum superficialis in a fetus at 28 weeks. H&E staining. Frontal sections tangential to the hand palmar surface. (A) (most superficial or palmar), (C, D) (deepest or most dorsal) display the proximodistal levels of the MP and CM joints at the lower magnification. (B) A higher magnification view of a square in (A), exhibits distal extensions of the FDS muscle. (A, C, D) were prepared at the same magnification. Between (A, C), the deep flexors also had distal extending muscle bellies (not shown). (A, C, D) were prepared at the same magnification. MP, metacarpopharangeal; CM, carpometacarpal; FDS, flexor digitorum superficialis, FDP, flexor digitorum profundus tendon; LU, lunate; PQ, pronator quadratus muscle; RA, radius; SC, scaphoid; TR, trapezium.
|
 | Fig. 2Transverse sections of the wrist showing distally extending muscle fibers of the long flexors and extensors in a fetus at 32 weeks. H&E staining. A single cross section. (A) displays topographical anatomy of the tendons at the wrist. (B, C) (same magnification) are higher magnification views of squares in (A), respectively. (D, E) (or F, G) are higher magnification views of squares in (B) (or C): these four panels were prepared at the same magnification. Muscle-tendon borders are encircled or indicated by dotted line (B–G). In the carpal tunnel, the 3th and 5th fingers’ bellies of the FDS muscle as well as all four bellies of the FDP muscle extend distally. The lumbricalis muscle appeared in the distal side of the figure. In the extensor side, a deep tendon of the ED muscle as well as the ulnar margin of the EPL muscle carries distally-extending muscle fibers. (B, C) (or D–G) were prepared at the same magnification. BPL, abductor pollicis longus muscle; ECRB, extensor carpi radialis brevis muscle; ECRL, extensor carpi radialis longus muscle; ECU, extensor carpi ulnaris muscle; MN, median nerve; UN, ulnar nerve; MP, metacarpopharangeal; CM, carpometacarpal; FDS, flexor digitorum superficialis; FDP, flexor digitorum profundus; ED, extensor digitorum; EPL, extensor pollicis longus.
|
Muscle bundles in the ankles were found to extend distally beyond the talocrural joint in the flexor hallucis longus (FHL) in sagittal sections of five of ten ankles and horizontal sections of five of six ankles. Likewise, the fibularis brevis (FB) extended distally in sagittal sections of eight of ten ankles and horizontal sections of all six ankles. The similar evidence was found in the extensor digitorum longus (EDL) in sagittal sections of five of 10 ankles and horizontal sections of four of six ankles. Since it provided a thin layer attaching to the tendon, whether the distally-extending muscle fibers was clear or unclear depended on the sectional histologic planes. Sagittal sections (Fig. 3) were liable to miss a longitudinal cut of a tendon but often detected tissue damage or an oblique cut. The FHL muscle fibers surrounded the tendon and were gradually inserted into the tendon, but they did not reach the flexor retinaculum (Fig. 4). The FB tendon was found to originate from the medial edge of the muscle belly, reaching the level of the talocalcaneal joint below the superior retinaculum. The EDL muscle fibers extended distally toward the retinaculum along the deep or posterior aspect of the aponeurosis-like tendon. In late-term fetuses, the muscle-tendon border provided a 2-dimensional irregularly curved line, with the interface occupied by loose fibrous tissue (i.e., the epimysium).
 | Fig. 3Longitudinal sections of the ankle showing distal extensions of long crural muscle bellies in a late-stage fetus at 30 weeks. H&E staining. Sagittal sections from a fetus at 30 weeks. (A) (or D) displays a medial (or lateral) plane through the ankle.at the ultralow magnification. (B) (or E) is a higher magnification view of a square in panel A (or D). (C) corresponds to a square in (B). In (A–C), a muscle belly of the FHL muscle changes into the tendon at a level below the talocrural joint surfaces. In (D, E), a muscle belly of the FB muscle also extends to a level below the joint surface. Both muscle bellies are adjacent to the joint capsule. (A, D) were prepared at the same magnification. AT, Achillis (or calcaneal) tendon; EHL, extensor hallucis longus tendon; FDL, flexor digitorum longus tendon; PTFL, posterior talofibular ligament; T nerve, tibial nerve; FHL, flexor hallucis longus; FB, fibularis brevis.
|
 | Fig. 4Transverse sections of the ankle showing crural muscle fibers extending distally in a fetus at 31 weeks. H&E staining. Horizontal section. (A) displays a level of the talocrural joint, while (D) exhibits the talocalcaneal joint. (B, C, E) are higher magnification views of squares in (A), respectively, while (F) is that in (D). In the four panels, a muscle tendon border (dotted line) is shown in the FB, EDL, and FHL. (A, D) (or B, C, E, F) were prepared at the same magnification. AT, Achillis (or calcaneal) tendon; FDL, flexor digitorum longus tendon; FL, fibularis longus; PTFL, posterior talofibular ligament; TN, tibial nerve; TP, tibialis posterior; FB, fibularis brevis; EDL, extensor digitorum longus; FHL, flexor hallucis longus.
|
Findings in adult cadavers
Based on the results showing muscle-tendon interfaces in fetuses, the FDS and FDP at the wrists, as well as the FHL, FB and EDL at the ankles, were macroscopically examined in adult cadavers. For observations, we cut and removed the transverse carpal ligament at the wrist and a set of the fibular and flexor retinaculum at the ankle. However, the dissection was unlikely to change a relative position of the distal muscle belly. The FDP, FHL and EDL did not carry muscle fibers extending distally beyond the joint (Fig. 5). In 12 of 15 wrists, however, one to three tendons of the FDS accompanied a very thin muscle layer reaching the level of the radiocarpal joint (Fig. 5A). In four of 15 wrists, the thin muscle extended distally across the joint along the tendon to the second or third finger. A distal muscle bundle of the FB was found to be thin, but it extended across the talocrural joint to reach the level of the talocalcaneal joint. The FB bundle, which faces a large loose tissue along the Achilles or calcaneal tendon, was found to be separated from the lateral malleolus by a thick tendon of the fibularis longus (Fig. 5C). The FHL tendon was also embedded in the same loose tissue, but it was attached tightly to the medial malleolus.
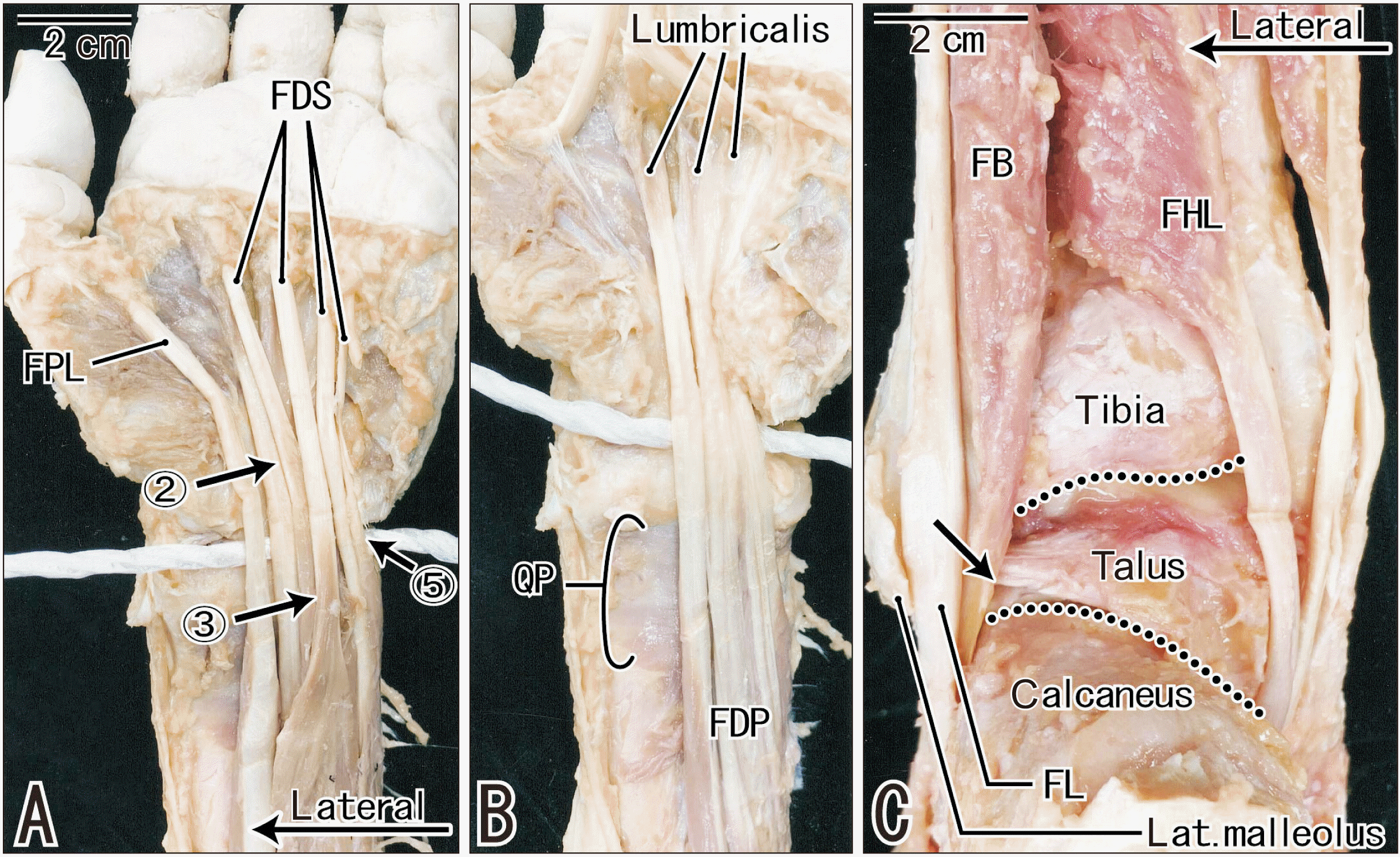 | Fig. 5Macroscopic observations of tendons and muscle bellies at the wrist and ankle of an 86 years-old man. (A) displays the FDS and FPL at the hand and wrist, while (B) exhibits the FDP and QP after removal of the superficial layer. A thick white string indicates a level of the radiocarpal joint surface. Arrows with number indicate the FDS muscle fibers extending across the joint along the tendons to the second, third and fifth fingers. The FDP muscle fibers do not reach a level of the joint (B). (C) is a posterior view of the ankle after removal of Achillis tendon and triceps surae muscles. Two dotted lines indicate the talocrural and talocalcaneal joints, respectively. Behind the lateral malleolus of the fibula, a muscle belly of the FB extends distally along the FL tendon and appears to reach a level of the talocalcaneal joint (arrow). The FHL muscle fibers does not reach the joint. (A, B) were prepared at the same magnification. FDS, flexor digitorum superficialis; FPL, flexor pollicus longus; FDP, flexor digitorum profundus; QP, quadratus plantae; FB, fibularis brevis; FL, fibularis longus; FHL, flexor hallucis longus.
|
Go to :
Discussion
In the Introduction, we had hypothesized usual over-production of muscle belly extending over the joint that the muscle acts on. The present observation in late-term fetuses confirmed the hypothesis although they did not correspond to all muscles at the wrist and ankle. However, the long extending fibularis muscle belly in adults was out of our expectation.
To our knowledge, the present study is the first to report that a fetus-specific muscle fiber or bundle extended distally across the involved joint. Of the candidate muscles examined, the FDP was the most striking example because of the consistency of the muscle fibers in fetuses and because of the association of tendon morphology and carpal tunnel syndrome. The distally-extending muscle fibers were found to surround the tendon and were clearly separated from the lumbricalis muscle. However, any extension movement at an infant’s wrist would result in a friction injury to the over-extending FDP. In fetuses, extensors also carried distally extending muscle fibers in the dorsum of the wrist. During the degeneration process after injury in infants, inflammation would occur in the tendon sheath, resulting in pain and/or mechanical resistance. It is likely therefore that infants ‘dislike’ extension and/or flexion at the wrist to avoid friction between the muscle fiber-associated tendon and the radius.
The distally-extending muscle bundle across the joint would likely result in the muscle action being less effective because the bundle may expand rather than contract in response to contraction of the proximally-located, usual muscle belly. The maintenance of the FB bundle in adults may be associated with its protection against friction stress, even during inversion and extension at the ankle. The FB bundle seemed to be separated from the lateral malleolus by a thick tendon of the fibularis longus and from the calcaneal tendon by thick loose tissue. Moreover, the FB action is masked, not only by strong eversion by the fibularis longus, but by strong flexion by the FHL and flexor digitorum longus. The lack of attachment of distally-extending muscle fibers to the joint capsule suggests that these fibers do not stabilize the joint.
The distally-extending muscle fibers provided a specific type or site of muscle-tendon interface, with the latter defined on a molecular basis [9, 10]. Similar to the usual type of muscle fiber convergence to an intramuscular tendon [11, 12], the unique muscle fibers described in the present study were also attached to and surrounded the proper insertion tendon near the wrist and ankle joints. Subsequent degeneration of the distal muscle fibers may be advantageous and its occurrence after birth may not be a rare developmental change, especially at the origin from a bone [5, 8]. Postnatal tendon maturation at the muscle-tendon interface may require some degree of degeneration of excess muscles fibers surrounding the tendon itself.
Overall, especially in children, a muscle belly might be able to extend over the joint that the muscle acts on. However, without any symptom such as pain and inflammation, a clinical imaging seemed to be difficult to identify the over extension of muscle fibers.
Study limitation
Since the present specimens were limited in number, we were not able to evaluate individual variations in site and incidence of the over-extending muscle belly. Moreover, a decreased length of muscles by postmortem fixation was likely to reduce the incidence. Ultrasound examination of living volunteers may provide a finding of not a few cases of the distally-extended muscle bellies.
Go to :
 XML Download
XML Download