

 PDF
PDF Citation
Citation Print
Print



Introduction
Tremendous advances in vascular biology have been achieved in recent years. Physiologically, the endothelium comprises highly specialized cells that line the blood and lymphatic vessels, weigh 1 kg and build approximately 400 m2 in an adult person that maintains approximately 60,000 miles of blood vessels [1]. Endothelial cells (EC) performs a wide range of crucial metabolic homeostatic activities to maintain a balance between aggression and protection factors by responding to extrinsic and intrinsic signals and releasing specific active molecules, as well as remodeling signaling pathways [1-3]. Endothelial cell homeostasis is crucial for maintaining the functionality of the whole organism. Classically, EC regulate vascular permeability, angiogenesis, immune system, pro- and anti-coagulant system, promotion of vasodilation, antioxidant effects, anti-inflammatory effects, inhibition of leukocyte adhesion and migration, inhibition of smooth muscle cell proliferation and migration, inhibition of platelet aggregation and adhesion, profibrinolytic effects, and regeneration of tissues [4].
Autophagy belongs to the four main types of cell death; apoptosis, autophagy, necrosis, and entosis [5]. However, stressed EC experience one of the following types of cell death, apoptosis, autophagy, or pyroptosis [6, 7]. According to the pathophysiological changes in the molecular signaling pathway and the triggering molecules, and severity of oxidative stress as well as the likelihood of endothelial cell to survive, the EC decide the most appropriate pathway of cell death. EC apoptosis is upregulated during oxidative stress, nutrient starvation, tumor necrosis factor-α (TNF-α), high glucose, lipopolysaccharide, homocysteine [8]. However, the full mechanism of regulation of endothelial cell death and the decision on the death pathway is not fully studied.
Vascular EC impairment involved in the pathogenesis of various diseases, including sepsis, diabetes, cerebral ischemia, cardiac ischemia/reperfusion injury, atherosclerosis, and chronic kidney disease [9-11]. Therefore, the analysis of the function of the endothelial cell is crucial and involved in the treatment strategy of such common diseases.
Several kinds of basal autophagy present in norm including macroautopahgy, microautopahgy, and chaperon mediated autophagy. Macroautophagy is the most explored type, which involves the turnover of intracellular components such as proteins, lipids, and organelles, as well as micronucleophagy. Selective autophagy targeting induces safer and more effective therapeutic results.
Potential Endothelial Cell Autophagy Triggers and Mechanism
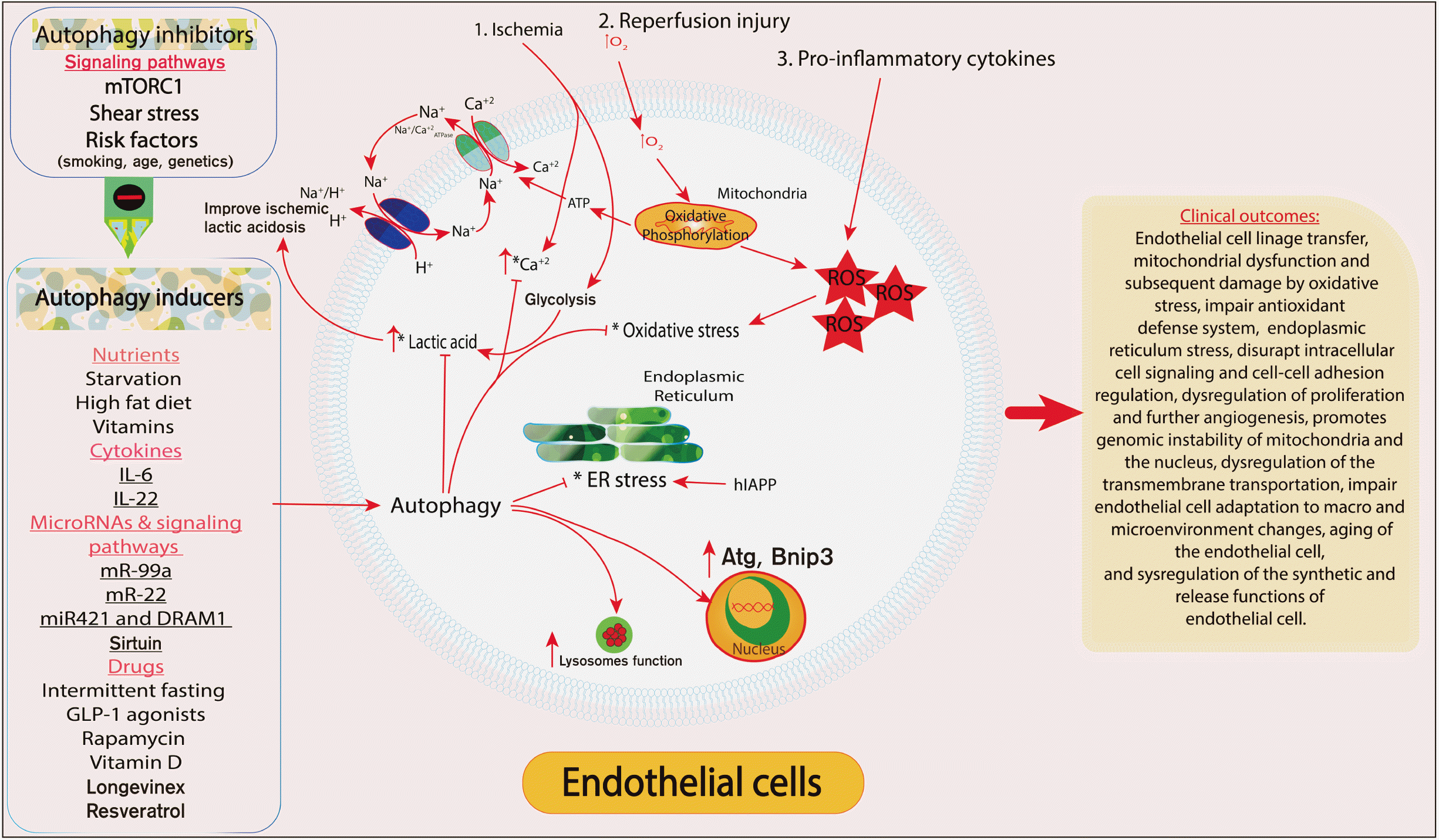
Endothelial cell damage can be induced by uncontrolled external mechanical shear stress and contractile forces such as mechanical stretching and vascular wall stress (biologically uncontrolled accidental cell death) or by regulated cell death [6]. Autophagy is a classical variant of regulated cell death. Usually, endothelial damage occurs due to mechanical shear stress and damage to the cell wall membrane. Whereas, regulated cell death (autophagy) is the primary target of future therapeutic strategy. Typically, early stages of regulated cell death are functional endothelial cell dysfunction. The boundaries between the reversible and irreversible change tends to be crucial during development of new therapeutic strategy for endothelial cell dysfunction.
Induction of endothelial cell damage is multifactorial and involved several pathways, including activation of autophagy stimulatory signaling pathways (miRNA-126, Apelin), inhibition of inhibitory signaling pathways (Notch1), stimulation of autophagy inducing receptors, inhibition of inhibitory receptors (caspase inhibitor Z-VAD-FMK, autophagy inhibitor 3-MA) and activation of autophagy inducing molecules and inhibiting autophagy inhibitory molecules [12-15].
Each of these has its role in the pathogenesis of endothelial cell autophagy dysregulation that subsequently results in endothelial cell dysfunction. Usually, a combination of these disorders coexists.
Cytokines and chemokines are the leading cause of endothelial cell dysfunction. Interleukin-6 (IL-6) is one of the first cytokines studied that induces endothelial cell dysfunction and the release of the endothelial plasminogen activator inhibitor-1 [16]. Classically, IL-6 levels are elevated during sepsis, bacterial infection, and acute respiratory distress syndrome. Hyperproduction of pro-inflammatory cytokines such as TNF-α, IL-8, and monocyte chemotactic protein-1 leads to cytokine release syndrome [16]. Cytokine release syndrome is a major stress to EC, prolonged exposure of EC to pro-inflammatory mediators.
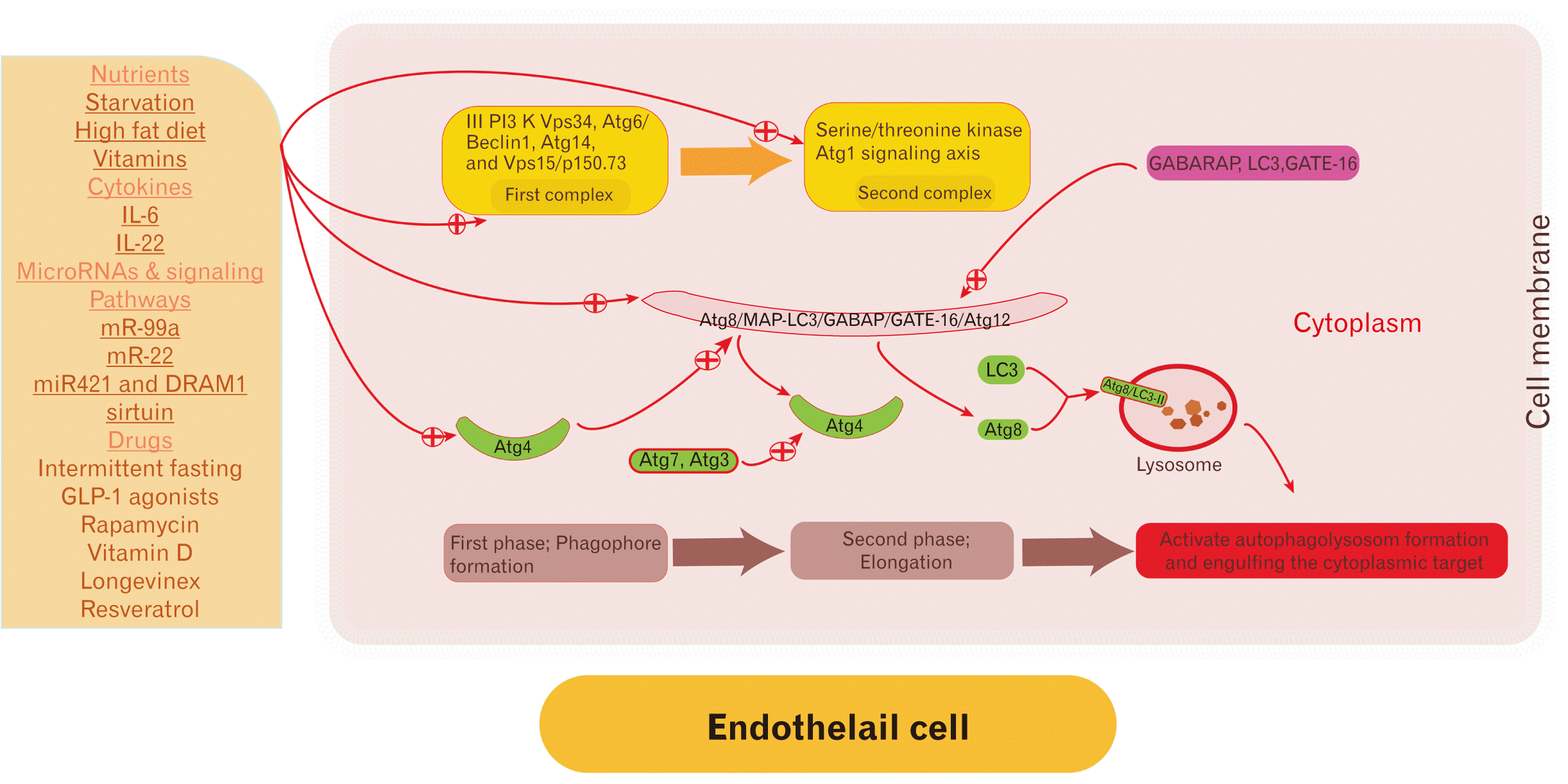
Generally, degradation of intracellular debridement’s can be achieved through microautophagy, macroautophagy, and chaperone-mediated autophagy depending on the debridement marker flag [17]. Endothelial cell autophagy mechanism is identical to that of myocardiocytes. However, the molecular triggers and inhibitors are different. Classically, autophagy is regulated by several autophagy-related proteins such as systems that produce modified complexes Atg8-PE and Atg5-Atg12-Atg16 [18]. More than 35 autophagy-related protein (Atg) genes have been identified, 17 of which are indispensable.
Mechanically, endothelial cell autophagy involves -Atg14L-Vps15-Vps34 proteins. Phagophore is the first phase of autophagosome formation. Two primary complexes signaling pathways involved in autophagosome formation [19, 20]. Autophagosome initiation induced by unc-51 like autophagy activating kinase 1 (Atg1) or by phosphatidylinositol 3-kinase (PI3K) Vps34 with Atg6 [21]. Autophagosome distention is regulated by transcription and post-transcription modulations, and epigenetics.
Initially, the first complex is composed of III PI3K Vps34, Atg6/Beclin1, Atg14, and Vps15/p150.73. Whereas the second complex mainly involves the serine/threonine kinase Atg1 signaling axis that its functionally depends on the presence of LC3, GATE-16 and GABARAP proteins [18].
Subsequently, autophagy elongation occurs that involves ubiquitin-like conjugation pathways; Atg8/MAP-LC3/GABAP/GATE-16 and Atg12 systems. Thereafter, the Atg8 protein undergoes cleavage for its carboxyl-terminal by the cysteine protease Atg4. Then, Atg4 undergoes a long complexed signaling cascade involved activation of Atg4 by Atg7 and Atg3 that culminate in activation of Atg8 and it is binding with the LC3–II of the lysosome and formation of Atg8/LC3–II complex. The Atg8/LC3–II complex bind with autophagosome in a covalent bound unless Atg4 cleaves it to be recycled and degraded by the phagolysosome [18]. In human, the fusion of the autophagosome with the lysosome remains not fully understood. Small GTPase Rab7 and lysosomal acidification through lysosomal H+-ATPase are required for successful fusion (Fig. 1).
Autolysosome formation regulated by genetic/epigenetic/transcriptional/posttranscriptional modifications [22].
Induction of endothelial cell autophagy occurs physiologically under hypoxic conditions. Several mechanistic pathways are potentially possible to take place, Inhibition of mTOR receptor, proapoptotic gene Bnip3, IP3 and lysosome inhibitors depending on the hypoxia severity. During anoxia, hypoxia-inducible factor 1, α subunit (HIF1A) is activated. HIF1A induces the expression of the Bnip3 gene (encode for BH3-only protein) which is required for disrupting the BCL2–BECN1 interaction by competitive binding of BCL2 (B-cell CLL/lymphoma 2) [23]. Bnip3 is an important gene for activating autophagy during hypoxic and non-hypoxic states.
The pathogenesis of endothelial dysfunction is multifactorial and involves chronic mild inflammatory reactions, direct physical stimulation and shear stress, chemical poisons and cytokine/chemokine release syndrome, concurrency of related diseases, and molecular changes on the cellular and subcellular level. The cell in rule has a precise equilibrium between the protective and aggression factors. Disruption to the natural balance and shifting the balance to the aggression side induces local endothelial dysfunction. Subsequently, over time, the prolonged predominant of the aggression factors resulted in multi-organ systemic changes in endothelial cell function. Development of an effective tool to identify the early stage of local endothelial dysfunction is crucial to prevent further progression into systemic multiorgan endothelial cell dysfunction. Seemingly, first place where endothelial cell get dysfunction and damage is in the branches and bifurcations and in regions of sharp vessel curvature where high shear stress. Early detection of level of the endothelial dysfunction markers such as E-selectin, ICAM-1, and VCAM-1, endoglin, endocan, von Willebrand factor, and soluble thrombomodulin is crucial to evaluate the risk of development of complications such as atherosclerosis [24-32]. Changes in the endothelial cell predicts the risk of endothelial cell injury. Recent study formulated an equation to early asses the risk of cardiovascular disease development depending on the flow-mediated dilation (FMD) of the arteries (FMD=–0.096BMI–0.069age–4.551WHR–0.015AST–0.242LDL+17.938) [33].
The most likely hypothesis is that in obese people with dyslipidemia, hyperglycemia, and hypertension, the EC develop functional memory and overtime reduces its response to the changes in the glucose level, blood pressure, and lipid profile. Persistent fluctuations in the metabolic syndrome component alter the functionality, biosynthesis, and response of EC to external stimuli. Prolonged changes results in mild chronic inflammation of the EC and the underlying smooth muscle cells. A vicious circle develops in the background each time the endothelial cell activity is impaired more and in the final stage there are anatomical and topographical changes in the EC. loss of endothelial cell function reduces the bioavailability of nitric oxide which subsequently leads to persistent vascular smooth muscle constriction and hypertrophy of the vascular smooth muscle cells [34]. Final outcome of this process is vascular damage and loss vascular tone regulation.
Types of Endothelial Cells
According to the topographical anatomy of EC, vascular and lymphatic endothelial cell exist. Several fundamental biological distinctions are present between lymphatic and blood EC despite the topographical similarities, more than 400 differences in the expressed genes, returning to the difference in the functions of each (Table 1) [39-46]. Lymphatic EC present semiprofessional antigen presentation to lymphatic circulatory T cytotoxic CD8 cells through an invadosome-shaped protrusion and express specific surface antigens and receptors such as podoplanin, vascular endothelial growth factor receptor-3 (VEGFR-3) or lymphatic vessel endothelial hyaluronan receptor-1 [47, 48].
Remarkable evidence concluded that sigma receptor-1 of the lymphatic EC is responsible for the regulation of the tone and barrier function as well as the permeability of the lymphatic tree through regulation of nitroxide biosynthesis signaling pathway [49].
Autophagy in Endothelial Cell Aging as a Marker
Recent meta-analysis showed approximately, 400 differentially expressed genes, 36 core-features, and 57 core-pathways have been found to be associated with ECs senescence [50]. Moreover, the phosphoglycerate dehydrogenase and the serine biosynthesis pathway are the predominant in EC senescence [50]. Impaired autophagy is a hallmark during aging of EC. Early evaluation of the level of the autophagy genes suggests the biological age of the cells, in addition with using the epigenome as a reliable tool to identify the biological age of the cells and particularly the endothelial cell (Fig. 2).
Current Approaches for Future Therapeutic Strategy
Endothelial cell dysfunction is a global issue requires massive attention. Current therapeutic strategies aimed to reverse the pathogenetic pathway and return the physio-anatomy of the EC. However, targeting autophagy as a therapeutic strategy remains in the early stages of study.
The therapeutic strategy includes lifestyle modification, antioxidant, autophagy inducers, control component of metabolic syndrome (dyslipidemia, hypertension, hyperglycemia), epigenetic modification, genetic modifications, transcription factors, DNA methylation or demethylation, microenvironment correction, cell-cell adhesion molecules correction.
Pharmacotherapy such as Resveratrol reduces endothelial cell dysfunction by modulating Bnip3 associated mitophagy [52]. Several other medications are in preclinical and clinical trials to assess their therapeutic value in the regulation of autophagy, such as Metformin, Rapamycin, Sirtuin, Velcade, Artenimol, and Longevinex [8].
Transcription factors has been seen to induce endothelial cell regeneration in animal models particularly pulmonary vessels EC by FoxM1 transcription factor [52]. The ability of the transcription factors (Oct4 and Klf4) to transdifferentiate other non-EC such as fibroblast into EC has shown promising results [53]. Moreover, upregulation of autophagy can be achieved by modification of autophagy gene regulation through transcription factors. Hypoxia-inducible factors-α induces endothelial cell regeneration in animal models through the FoxM1 transcription factor [54]. Sox17 is another specific TF for EC proliferation shows stimulatory effect when its expression upregulated [55]. Atf3 is another key transcription factor in the regeneration strategy of damaged EC [56]. These transcription factors induce common signaling pathways between autophagy and cell cycle regulation encoding genes. Therefore, autophagy induction induces cell regeneration and improves the functionality of impaired EC. Regeneration of EC by the transcription factors culminates in one common result. FoxM1 induces regeneration through upregulation the signaling G-coupled protein receptor–dependent p110γ isoform of phosphoinositide 3-kinase (PI3K). Coexistence of FoxM1 and Pik3cg expression enhances the regeneration capacity of the EC [57]. Several other signaling pathways involved in the regeneration of EC such as Apelin signaling, Notch1 signaling, miRNA-126P signaling, VEGF signaling, and insulin-like growth factor (IGF)-1 receptor (IGF1R) [14, 58-61].
Discussion
Autophagy plays a pivotal role in the pathogenesis of endothelial cell dysfunction. Where basal autophagy is required to prevent endothelial cell damage and survive the stress conditions as well as prevent endothelial cell reprogramming. However, reduced basal autophagy level is indirect cause of endothelial dysfunction and cell death. Majorly, mitophagy is impaired during endothelial cell dysfunction which is responsible for oxidative stress alleviating and recycling impaired mitochondria [62, 63].
Currently, several invasive and non-invasive methods are known to evaluate the functionality of EC in different parts of the vascular tree. Of these methods are venous occlusion plethysmography, flow-mediated dilatation, pulse wave analysis, peripheral arterial tonometry, laser Doppler flowmetry, biochemical markers and bioassays, biochemical markers and bioassays, endothelial microparticles, endothelial progenitor cells, endothelial progenitor cells, and endothelial glycocalyx [64]. Autophagy interferes with apoptosis signaling pathway regulation and several molecules can interact with autophagy and prevent apoptosis and induces necrosis of EC [65-67]. Apoptosis involved common caspase signaling pathway, blocking of caspases activity induces regulated necrosis such as in endothelial cell; pyroptosis [68].
Conclusion
The impairment of endothelial cell autophagy is the basis for endothelial dysfunction. Autophagy does not only self-eating the intracellular component; autophagy effectively participates in maintaining endothelial cell homeostasis. Autophagy prevents cell linage transfer, stabilizes mitochondrial function subsequent protection from oxidative stress, improves cellular antioxidant defense system, prevents endoplasmic reticulum stress, maintains intracellular cell signaling, regulation cell-cell adhesion, enhances proliferation and further angiogenesis, promotes genomic stability of mitochondria and the nucleus, improves transmembrane transportation including mitochondria permeability transition, improves endothelial cell adaptation to macro and microenvironment changes, and improves the synthetic and release function of impaired endothelial cell [68].
 XML Download
XML Download