

 PDF
PDF Citation
Citation Print
Print


Introduction
Induced pluripotent stem cells (iPSCs) can be generated from different types of somatic cells including fibroblasts and keratinocytes by ectopic expression of four reprogramming factors OCT4, SOX2, KLF4, and MYC (hereafter referred to as OSKM) (1). iPSCs hold great promise for regenerative medicine by virtue of their unlimited proliferation and multilineage differentiation capacity (1, 2). More specifically, iPSCs generated from isolated patient cells and their derivatives provide powerful in vitro models with which elucidation of cellular and molecular mechanisms underlying disease pathogenesis and development of new drugs to alleviate disease phenotypes are possible (3). Furthermore, genetically and phenotypically corrected patient-specific iPSCs can be used for preclinical and clinical trials to investigate their potential for replacing damaged cells/tissues (2).
Pelizaeus-Merzbacher disease (PMD) is an X-linked recessive hypomyelination disorder that is caused by mutations in the proteolipid protein 1 (PLP1) (4). PLP1 is specifically expressed in oligodendrocyte precursor cells (OPCs) and oligodendrocytes (OLs) and plays decisive roles in compaction, stabilization, and maintenance of myelin sheaths (5). Dysregulation of these processes that are elicited by mutations in PLP1 can lead to death of OLs, consequently resulting in the loss of myelin sheaths (6, 7). More than 100 different mutations in PLP1 have been reported in PMD, including duplications, point mutations and null mutations (4). Duplications of PLP1 are the most common mutations, which have been detected in over 60% of PMD patients. Missense and nonsense PLP1 mutations have been reported in over 20% PDM patients. Null mutations are relatively rare. Clinical symptoms (phenotypes) of PMD patients are diverse (4), but whether/how these phenotypic diversities between PMD patents are actually linked to different types of genotypic mutations remains unknown. Furthermore, cellular and molecular mechanisms underlying the pathogenesis of PMD are not yet fully defined. Hence, establishment of iPSC lines from individuals with mutations spanning genetic and clinical diversities of PMD may enhance our understanding of PMD pathogenesis and genotype-phenotype relationship.
Lymphoblastoid cell lines (LCLs) have been generated by infection of B cells from peripheral blood of healthy donors and patients with Epstein-Barr Virus (EBV) (8). These EBV-immortalized LCLs are a convenient and useful model to study a variety of human genetic disorders. In fact, they have been used for various biomedical studies including disease-related gene discovery and mutational analyses (9, 10). Importantly, over tens thousands of LCLs have been deposited internationally from a variety of individuals with various ethnic backgrounds and disease states (11-13). For instance, the Coriell Institute for Medical Re-search currently stores twenty-three LCLs which are deposited from ten PMD patients and thirteen related heal-thy family members. iPSCs generated from these LCLs and their derivatives (OPCs and OLs) could provide a valuable model for studying demyelination diseases and discovering therapeutic options for PMD.
In this study, we attempted to develop an efficient gene delivery method for LCLs. We found that electroporation is an efficient means to deliver genes into LCLs. With this strategy, we successfully generated PMD-specific iPSCs from LCLs. These iPSCs expressed pluripotency markers and could form embryoid bodies. Our findings show that electroporation is an effective method with which disease-specific iPSCs can be generated from LCLs.
Materials and Methods
Cell culture
Epstein-Barr virus immortalized lymphoblastoid cell lines (LCLs) were obtained from the Coriell Institute. The Coriell cell repository maintains the consent and privacy of the donor LCLs. LCLs were cultured in RPMI 1640 (Gibco) supplemented with 15% fetal bovine serum (FBS; Biochrom) and 2 mM L-glutamine (Sigma-Aldrich). CRL-2097 neonatal skin fibroblasts and HEK293 cells were purchased from ATCC and cultured in Dulbecco’s Modified Eagle’s medium high glucose (DMEM; Welgene) supplemented with 10% FBS, 1×nonessential amino acids (NEAAs; Sigma-Aldrich), 1×GlutaMAX (Gibco), and 1×penicillin/treptomycin (P/S; 100 U/ml each, Sigma-Aldrich). Human iPSCs were cultured on the Matrigel-coated plate with MEF-conditioned medium (MEF-CM). The MEF-CM medium was prepared as described previously (ref). The cells described in this study were cultured at 37℃ and 5% CO2 in a humidified Heraeus BB15 incubator (Thermo Fisher Scientific).
Plasmid
pLEX307-GFP and pCXLE-mCherry were cloned in house. pMXs-OCT4, pMXs-SOX2, pMXs-KLF4, pMXs-c-MYC, pMXs-GFP and pRRL-hOSKM-tdTomato were previously described (14-16). Other plasmids including pMD2.G (#12259), psPAX2 (#12260), pUMVC (#8449), pCMV-VSV-G (#8454), pCXLE-hOCT3/4 (#27076), pCXLE-hSK (#27078), pCXLE-hUL (#27080), and pCXLE-mp5 3DD (#41859) were purchased from Addgene. Plasmid DNA was isolated from E. coli and purified using Nucleo-Bond Xtra Kit (Macherey-Nagel).
Virus production and infection
To produce retroviruses, 4.5 μg of pMXs-GFP, 3 μg of pUMVC, and 1.5 μg of pCMV-VSV-G were transfected into HEK293 cells using 27 μl of Polyethylenimine (PEI; Polysciences) in 600 μl Opti-MEM (Invitrogen). At 48 h post-transfection, virus-containing supernatants were collected and filtered through a 0.45-μm PVDF filter (Mil-lipore), concentrated at 23,000 rpm for 2 h using an Optima XL-100 K Ultracentrifuge (Beckman), and resuspended in 1 ml of DMEM. Retroviral suspensions were stored at −80℃ until use. To produce lentiviruses, 1.5 μg of pMD2.G, 3 μg of psPAX2 and 4.5 μg of the pRRL-hOSKM-tdTomato were transfected into HEK293 cells using 27 μl of PEI in 600 μl of Opti-MEM. At 48 h post-tra-nsfection, virus-containing supernatants were collected and filtered through a 0.45-μm PVDF filter, concentrated at 23,000 rpm for 2 h using an Optima XL-100 K Ultrace-ntrifuge and resuspended in 1 ml of DMEM. Lentiviral suspensions were stored at −80℃ until use. The LCLs and CRL2097 fibroblasts were then infected with viruses twice over the course of three days in the presence of 8 μg/ml protamine sulfate (Sigma-Aldrich).
Compounds
Sodium butyrate (NaB; Sigma-Aldrich) was used at a final concentration of 250 μM. SB431542 (purity: ≥98%; Cayman Chemical) was used at a final concentration of 2 μM. SGC0946 (purity: ≥98%; Cayman Chemical) was used at a final concentration of 3 μM.
iPSC generation
1×106 of LCLs were electroporated with total 2 μg of pCXLE-hOCT3/4, pCXLE-hSK, pCXLE-hUL, and pCXLE-mp53DD (0.5 μg each) using the NeonTM Transfection System 10 μl Kit (MPK10096; Thermo Fisher Scientific). The transfected LCLs were then transferred to 35 mm dishes pre-coated with hESC-qualified Matrigel (BD Bios-ciences) and cultured in MEF-CM supplemented with NaB, SB431542, and SGC0946. The medium was changed every other day. Chemicals were added until colonies reached the size of over 200 μm in diameter.
Flow cytometry analysis
The cells were mechanically or enzymatically disso-ciated and resuspended in 300 μl of 3% FBS/PBS. The cell suspension was transferred to 5 ml polystyrene round-bottom tubes through a cell strainer cap (Falcon) and subjected to flow cytometry analysis. The cells were then separated from debris and aggregates by forward scatter/side scatter (FSC/SSC) gating. Single cells were identified by plotting FSC area versus FSC width. Dead cells were excluded by staining with DAPI and gating on DAPI+ cells. Cells that had not been infected with viruses or had been electroporated with empty plasmids were used as negative controls for gating. Fluorescence was measured using a FACSCanto II (BD Biosciences) and flow cytometry data were analyzed using FlowJo software (Tree Star Inc.).
Immunofluorescence
The cells were fixed with 10% formalin (Cellnest) for 20 mins, incubated with 0.1% Triton X-100/PBS for 30 mins, and blocked in 5% BSA/PBS for 1 h. The cells were incubated with appropriate primary antibodies overnight at 4℃. The primary antibodies used were following: anti-SSEA-4 (1:100, 90231; Millipore), anti-NANOG (1:1,000, 5232S, Cell Signaling Technology), anti-OCT4 (1:1,000, 5677S; Cell Signaling Technology), anti-SALL4 (1:1,000, ab29112; Abcam). The cells were then washed three times with PBS and incubated with appropriate fluorescently labeled Alexa-Fluor secondary antibodies (1:1,000, Invitrogen) for 1 hr. The cells were then washed three times with PBS, incubated with 0.5 μg/ml 4’,6-dia-midino-2-phenylindole (DAPI, Molecular Probes) for 10 mins, and washed once with PBS. The cells were then subjected to immunofluorescence microscopy analysis. Images were acquired using a fluorescence inverted microscope (IX71; Olympus) equipped with the CCD camera (DP30BW; Olympus) and analyzed with the DP-BSW (Olympus) and Fiji software (17).
Results
Retroviral transduction in LCLs
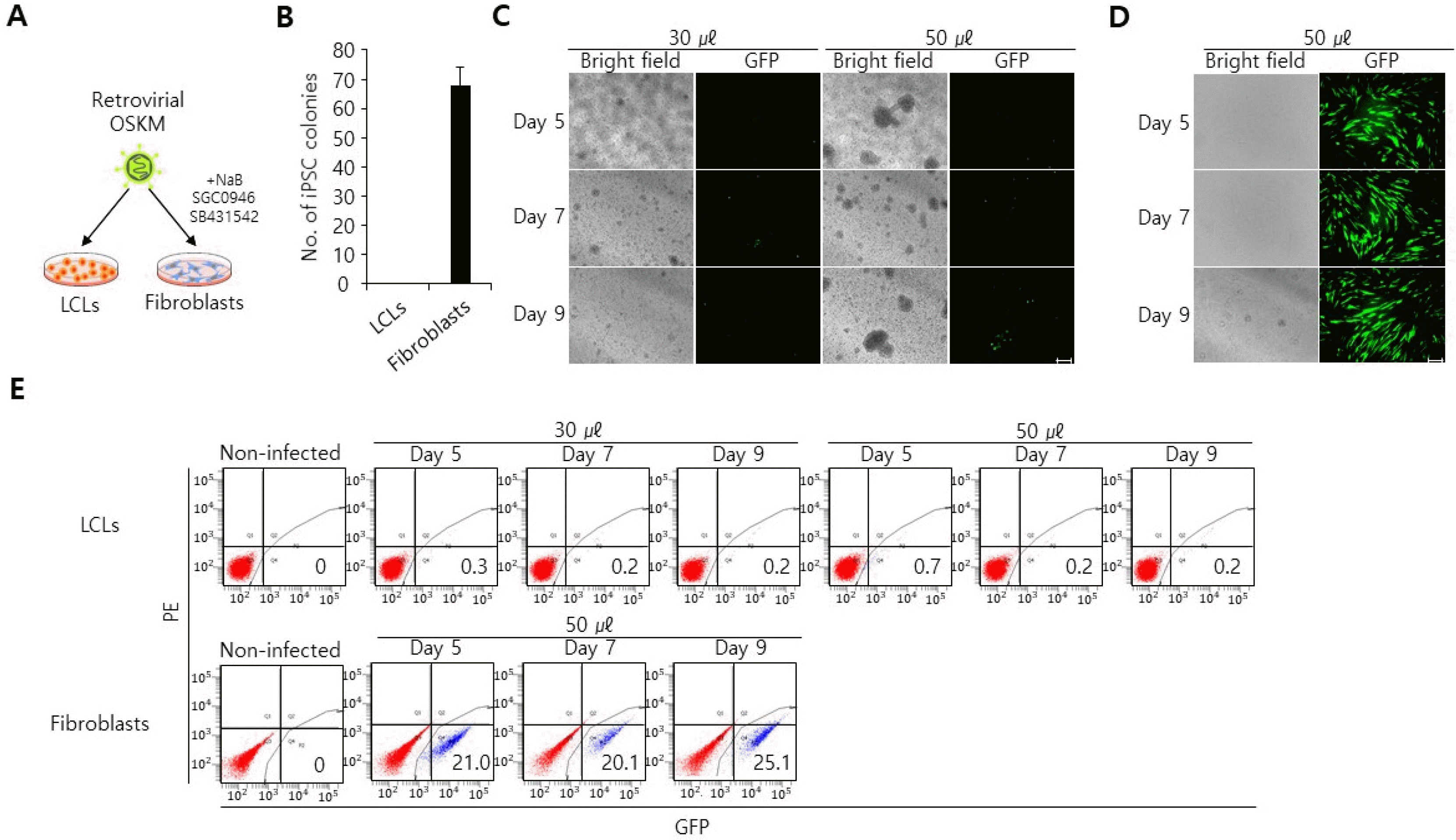
Retroviral transduction of OSKM has been widely used for generation of iPSCs from different types of human cells (15, 18). Since retroviruses can only infect actively dividing cells (19), our first choice for gene delivery into LCLs was retroviral transduction. We transduced LCLs with retroviruses containing OSKM and cultured in MEF-CM medium supplemented with sodium butyrate (a HDAC inhibitor), SB431542 (a TGFb inhibitor) and SGC0946 (a DOT1L inhibitor) (Fig. 1A). These chemicals positively influence iPSC generation (20). Surprisingly, retroviral transduction of OSKM in LCLs did not yield any iPSC colonies in contrast to concurrently transduced fibroblasts (Fig. 1B). We then hypothesized that the failure of generating iPSCs from LCLs might be due the fact that retrovirus might not effectively transduce LCLs. To test this hypothesis, we transduced LCLs with retroviruses encoding green fluorescent protein (GFP) and evaluated GFP expression by a fluorescence microscope. At five days post-infection, we found that GFP was not expressed in retroviral GFP-transduced LCLs (Fig. 1C). Neither increased titers of GFP virus or longer culturing periods (days 7 and 9) changed GFP negativity (Fig. 1C). In contrast, GFP was expressed in retroviral GFP-transduced fibroblasts (Fig. 1D). Flow cytometry analysis further confirmed that GFP was not expressed in retrovirally GFP-transduced LCLs, in contrast to concurrently transduced fibroblasts which yield over 20% GFP+ cells at day 5 post-infection (Fig. 1E). Together, these data demonstrate a decisive difference in retroviral transduction between two cell types and show that retrovirus-mediated gene delivery is not effective for LCLs.
Lentiviral transduction in LCLs
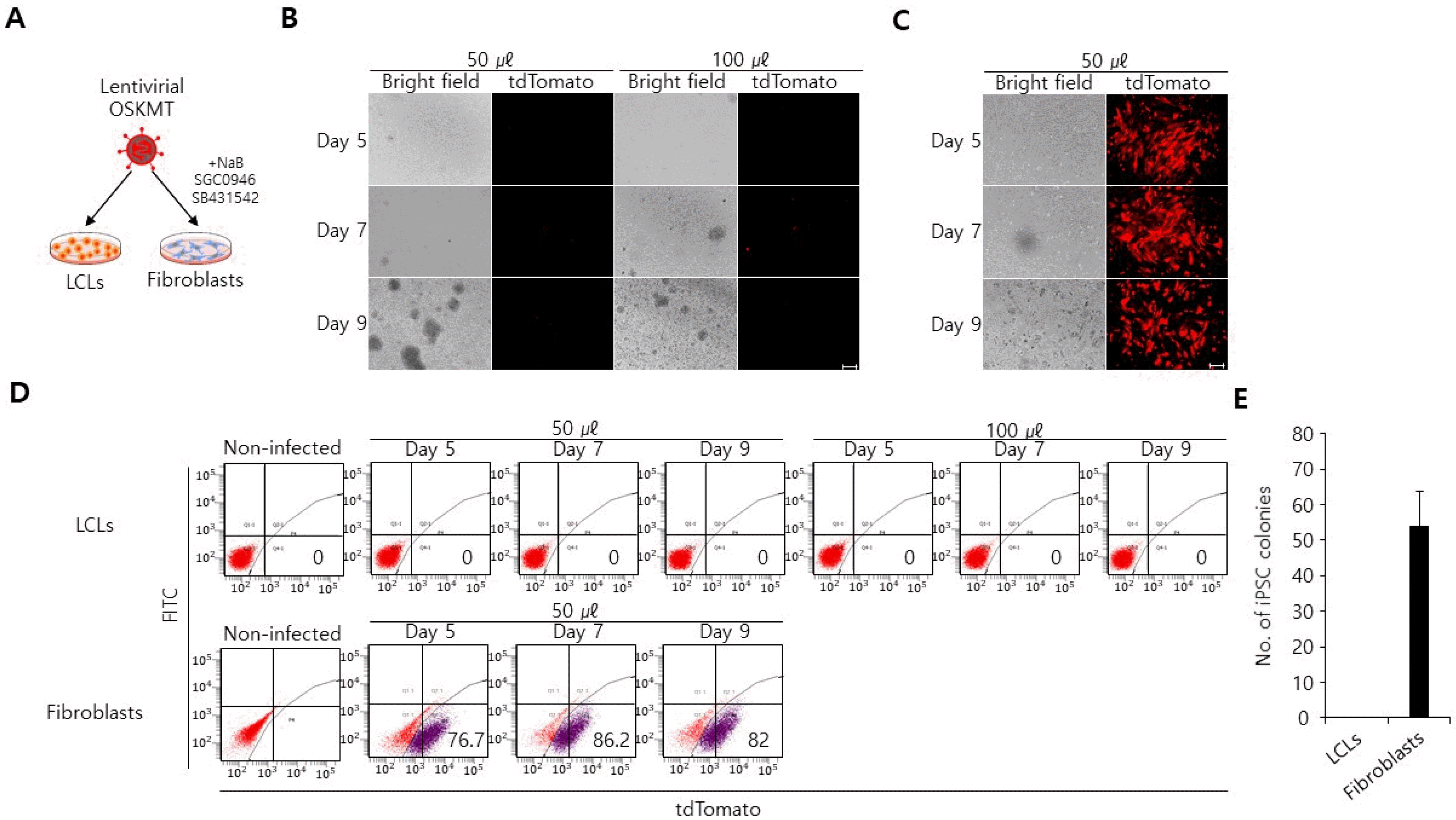
Lentiviruses can infect a wider range of cell types than retroviruses (21). We thus postulated that lentiviral transduction might be in fact an effective way to deliver genes into LCLs. To test this, we produced lentiviruses with a polycistronic lentiviral vector encoding OCT4, SOX2, KLF4, c-MYC, and tdTomato (hereafter referred as OSKMT) and transduced LCLs (Fig. 2A). In this system, tdTomato was linked to OSKM by an internal ribosomal entry site (IRES) and thus transgene expression can be monitored by tdTomato expression. Surprisingly, at forty-eight hours post-infection, we found that lentiviral OSKMT-transduced LCLs did not yield any tdTomato+ cells, in contrast to concurrently transduced fibroblasts which yielded over 70% tdTomato+ cells (Fig. 2B∼D). Consequently, le-ntiviral OSKMT-transduced LCLs failed to produce iPSC colonies, whereas concurrently transduced fibroblasts produced iPSC colonies (Fig. 2E). Together these data demonstrate lentiviral transduction is also not an effective way to deliver genes into LCLs and is not suitable for generating iPSCs from LCLs.
Electroporation in LCLs
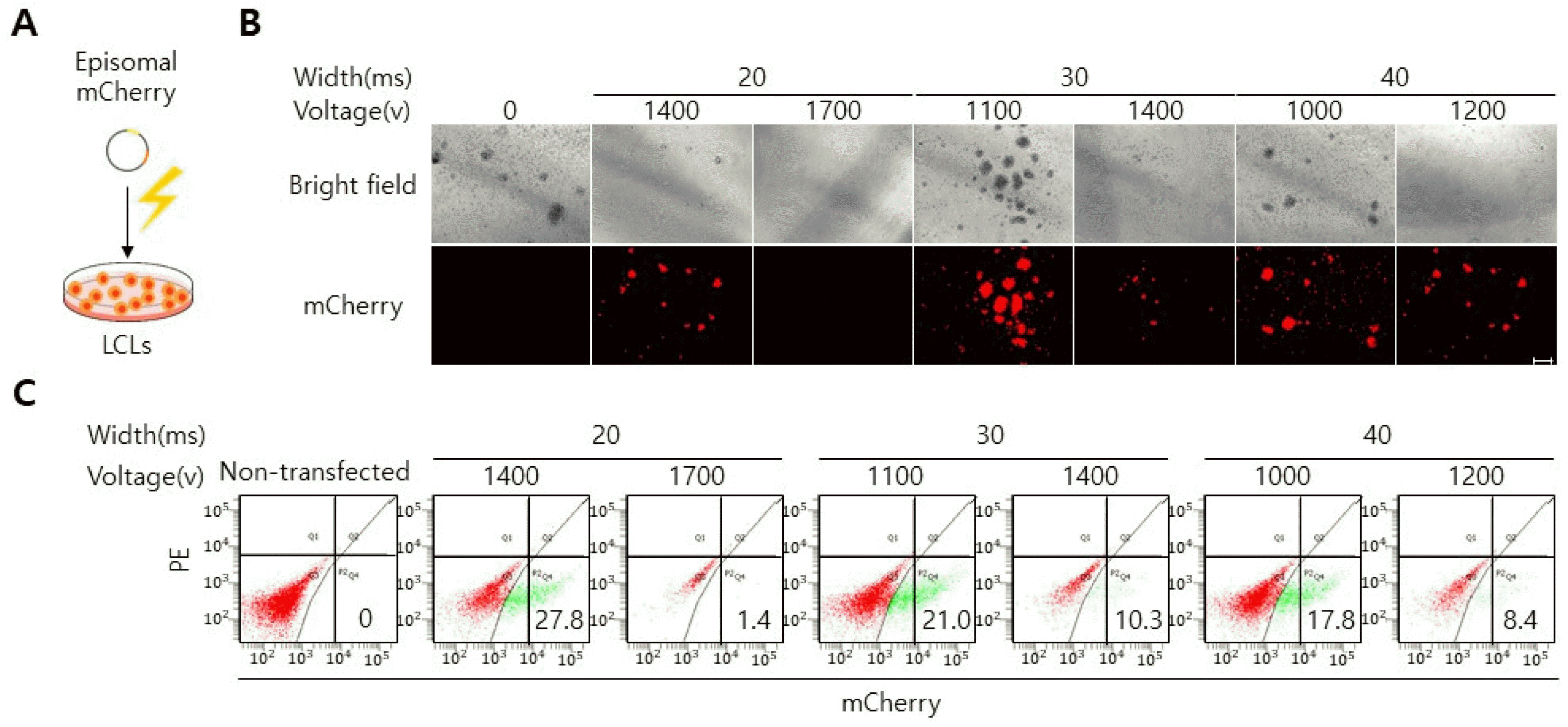
Beside retroviral and lentiviral transduction, electroporation is an efficient gene delivery method which can introduce genes into a wide range of cell types, especially hard-to-transfect cells, like primary cells and cells that are cultured in suspension (22, 23). Electroporation uses an electrical pulse to create temporary pores in cell membranes through which substances like plasmids can enter into the cells. Different cell types require distinct electroporation settings (i.e. pulse time, voltage strength) to achieve high viability and reproducible transfection efficiency (22, 23). To identify an optimal electroporation condition for LCLs, we electroporated LCLs with episomal vector encoding mCherry with different ranges of voltage stren-gth and pulse width (Fig. 3A). We selected an OriP/EBNA1-based episomal vector particularly as a vehicle to deliver mCherry into LCLs, since two elements (oriP and EBNA-1) allow for replication and long-term retainment of vectors in mammalian cells (24). At forty-eight hours post-transfection, mCherry expression was analyzed by a fluorescence microscope. Intriguingly, of six condition we tested, two conditions (20 ms/1,400 v, 30 ms/1,100 v) yie-lded over 20% mCherry+ cells (Fig. 3B and 3C). Of note, cell death rates increased with higher voltage conditions (20 width: 1,700 v, 30 width: 1,400 v, 40 width: 1,200 v), indicating that voltage strengths influence the cell viability (Fig. 3C). Together these data reveal defined electroporation conditions that enable efficient gene delivery into LCLs.
Reprogramming of LCLs into iPSCs
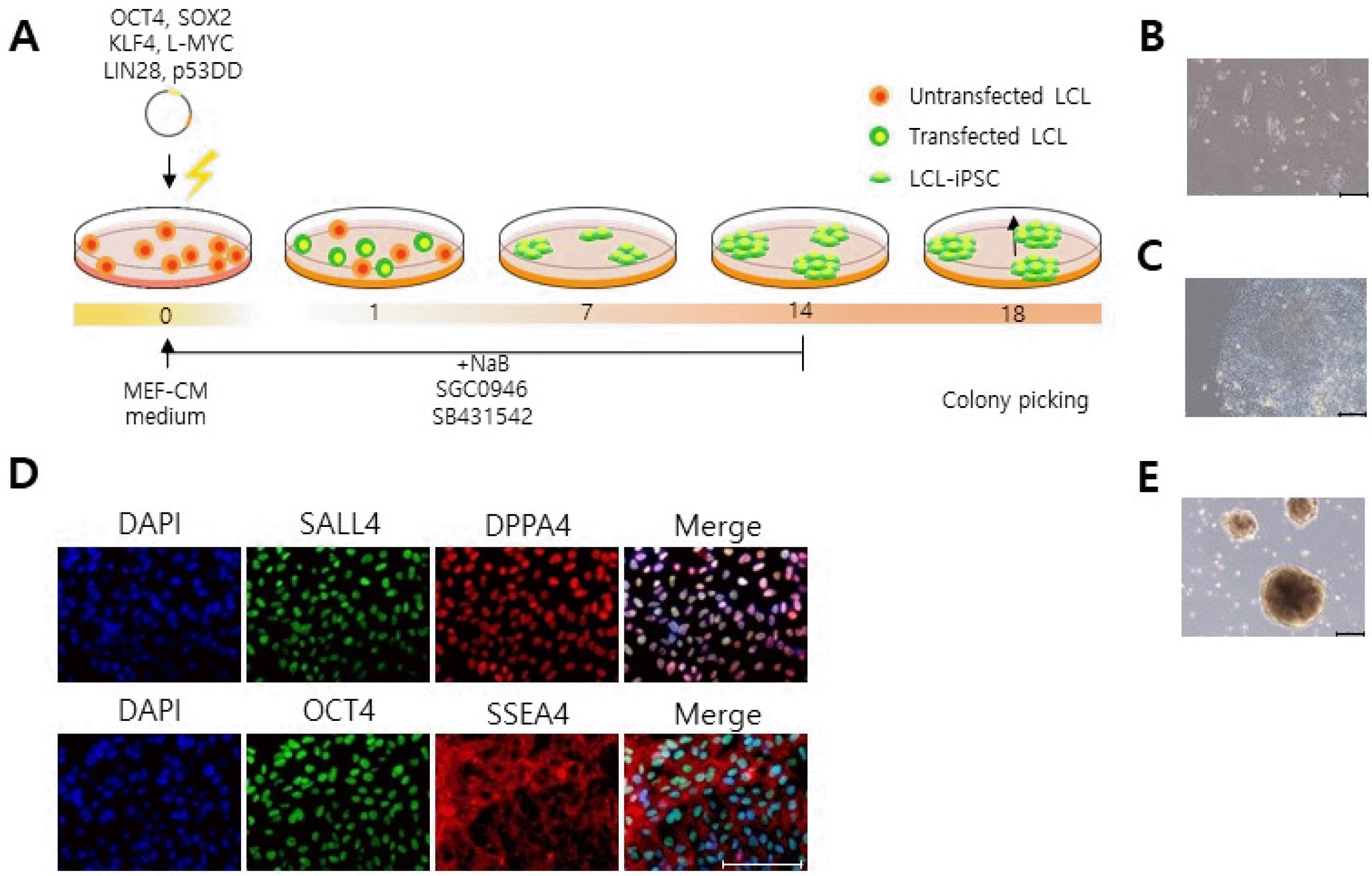
Having established that electroporation is a powerful and versatile method for delivery genes into LCLs, we next attempted to use this method to reprogram LCLs into iPSCs. For this, we electroporated LCLs, which were derived from a PDM patient, with episomal vectors encoding repro-graming factors [OCT4, SOX2, KLF4, L-MYC, LIN28 and p53DD (a dominant-negative inhibitor of p53 function)] and subsequently cultured them in MEF-CM medium supplemented with sodium butyrate, SB431542, and SGC0946 (Fig. 4A). At one week after transfection, we observed a number of small iPSC colonies firmly attached to the bottom of the culture plate (Fig. 4B). After the colonies reached over 800 μm in diameter (Fig. 4C), they were mechanically isolated and cultured independently. To confirm if the LCLs had transformed into iPSCs, we stained them with the number of pluripotency makers (Fig. 4D). We found that the LCL-derived iPSCs indeed expressed OCT4, SSEA4, DPPA4, and SALL4 (Fig. 4D). Further-more, they could form embryoid bodies when the cells were cultured in ultra-low attachment culture plates (Fig. 4E). Together these data demonstrate that electroporation of episomal vectors enables iPSC generation from LCLs.
Discussion
Lymphoblastoid cell lines (LCLs) can grow indefinitely in culture and derive from B lymphocytes infected by Epstein-Barr virus (EBV) (8). Because of their non-adherent nature in culture, LCLs are relatively easy to handle by researchers. Over tens thousands of LCLs have been deposited from a variety of patients with various ethnic backgrounds and disease states (11-13). Disease-specific iPSCs generated from these LCLs and their derivatives would provide a powerful in vitro model for elucidating disease pathogenesis and developing new therapeutic options.
Although LCLs are actively dividing cells (8), we found that retroviral transduction could not infect LCLs. Why LCLs appear to be refractory towards retroviral transduction remains unknown. However, one could readily expect that certain retroviral receptors are not present in LCLs such that retroviruses cannot dock to the surface of LSCs to initiate infection. Thus, it will be interesting to determine whether ectopic expression of some or any retroviral receptors might render LCLs susceptible to retrovirus infection.
Lentiviruses are capable of infecting a wider range of cell type and also it can infect even non-dividing cells (21). As such, our observation that LCLs cannot be transduced by lentiviruses was unexpected. For lentivirus-mediated reprogramming, we used a polycistronic lentiviral vector encoding OSKMT (pRRL-OSKMT) (16). In this vector, a retroviral spleen focus-forming virus U3 (SFFV) promoter regulates OSKMT expression. This promoter is known to mediate efficient expression of genes in only certain types of cells, but not in all cell types (16). Furthermore, it has been shown that this promoter can be rapidly silenced in some cell types, like hematopoietic cells and pluripotent stem cells (16, 25). Thus, the failure of lentiviral transduction in LCLs might be due to the activity of SFFV promoter present in pRRL-OSKMT. We found that elongation factor 1 alpha (EF1α) promoter-derived lentiviral GFP was effectively expressed in LCLs, which supports this idea (Supplementary Fig. S1A∼C).
Given the tremendous potential of iPSCs for regene-rative medicine and disease modeling, generation of disease-specific iPSCs have become important (2). In this study, we generated PMD-specific iPSCs from LCLs which derived from a PMD patient. PMD is a demyelinating disease that eventually results in developmental delays and neurological dysfunction, and death (4). Current pharmaceutical treatments cannot cure PMD. With PMD-iPSCs and their differentiated derivatives (OPCs and OLs), future studies will aim to develop new therapeutic perspectives and investigate PMD pathogenesis.
Supplementary Materials
Supplementary data including one figure can be found with this article online at https://doi.org/10.15283/ijsc22177.
 XML Download
XML Download