

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Inflammatory bowel disease (IBD) comprising CD and UC is believed to reflect complex interplay between genetic and environmental factors. CD is one of the major type of IBD. CD and UC exhibit etiologically distinct characters. CD often involves in the ileum and colon mostly but also can involves in any region of the gut. UC mostly common involves the rectum, and it has been known that the inflammation may extend the cecum in a contiguous pattern [1]. Inflammatory diseases such as CD are usually affected due to genetic factors and environmental factors [2]. A recent genome-wide association study successfully identified more than 200 CD-associated loci at the level of genome-wide significance [3,4]. However, these findings could only address a few part of the IBD pathogenesis. Numerous genetic, clinical, and multiple experimental studies should have been proved to our understanding of the etiology of IBD.
DNA methylation, predominantly occurring in the CpG dinucleotide of the genome, is a key epigenetic mechanism that can regulate gene expression and therefore can affect to the development and progression of complex diseases including cancer [5]. Aberrant DNA methylation frequently involves in inactivation of tumor suppressor genes (TSGs) in human cancers [5]. Among others, the transcriptional silencing of tumor suppressor candidate genes is regulated by promoter hypermethylation in colorectal cancer (CRC) by our group and other groups as well [6,7]. High-throughput genome-wide methylation studies provide a brand-new approach to understand the importance of DNA methylation at the whole genome level and its impact on gene regulation [6,8]. Therefore, the role of DNA methylation has been widely studied in many different cancer types and other human diseases, such as IBD [9]. For example, abnormal DNA methylation of the estrogen receptor, p14ARF, and E-cadherin genes has been detected in UC patients. In addition, there is growing evidence that epigenetic changes in the regulation of gene expression are critical factors in the pathogenesis of IBD [10,11]. Many studies have used these wellknown TSG candidates to investigate other cancers, such as lung, breast, and pancreatic cancer [7,12]. We previously have reported that CpG islands in the promoter region of TCERG1L is abnormally hypermethylated at high frequency not only in colon cancer patient tissues [13], but also in patients with UC [14]. However, few studies have been conducted on the DNA methylation profiles of these TSGs in patient with CD.
We tested the promoter hypermethylation of 18 tumor suppressor candidates (TFPI2, GATA4, p16, sFRP1, sFRP4, MGMT, Sox17, HIC1, sFRP2, sFRP5, p14, p15, GATA5, E-cadherin, and Timp3) in a limited number of tissue samples from Korean patients with CD (n = 15). Interestingly, we detected the abnormal promoter hypermethylation of several TSGs in patients with CD: SFRP1, SFRP2, SFRP5, TFPI2, Sox17, and GATA4 were aberrantly hypermethylated with high frequency in most of the CD patient tissues. Our data suggest that the methylation status of 6 genes in patients with CD has the potential to become a marker of risk for the progression of severe disease.
METHODS
1. Patients and Samples
The CD and normal colon biospecimens for this study were provided by the Inje Biobank (Inje University School of Medicine), a member of the National Biobank of Korea, which is supported by the Ministry of Health and Welfare. Tissue samples were collected from the ascending colon at the time of colonoscopy. Of the 15 subjects, 10 were male and 5 female, yielding a male to female ratio of 2:1. The median age at diagnosis of CD was 28.4 years (range, 18–45 years). CD was diagnosed on the basis of conventional clinical, radiologic, endoscopic, and histopathologic criteria. At the time of diagnosis, 3 patients (20%) had disease in the colon alone (L2), and 12 (80%) had disease in the small bowel and colon (L3). Disease behavior at diagnosis was inflammatory (B1) in 10 patients (66.7%), stricturing (B2) in 4 (26.7%), and penetrating (B3) in 1 (6.6%). Clinical characteristics of these patients are shown in Table 1. This study was approved by the respective institutional review board (IRB No. 129792-2015-056) of the participating institutions of the National Biobank of Korea, and written informed consent was obtained for all study participants prior to data collection.
2. Genomic DNA Extraction and DNA Methylation Analyses
Genomic DNA was extracted following a standard phenolchloroform extraction protocol. Bisulfite modification of DNA was performed using the EZ DNA Methylation Kit (Zymo Research, Irvine, CA, USA) according to the manufacturer’s instructions. Methylation analysis by methylation-specific PCR (MSP) was carried out in a 25-μL reaction of 10 × PCR buffer, 10 mM dNTPs (deoxyribonucleotide triphosphates), 10 pmol of each of the methylated or unmethylated primers, 1 unit of JumpStartTM REDTaq® DNA polymerase (Sigma, St. Louis, MO, USA) and 4 μL of bisulfite-treated DNA. The detailed information of primer was listed in in Table 2. Cycling condition were: 5 minutes at 95°C followed by 35 repeats of the following cycle: 95°C for 30 seconds, annealing at the appropriate temperature for 30 seconds. In vitro methylated DNA was used as a positive control.
3. Bisulfite Sequencing Analysis
Bisulfite modification of DNA was used for PCR reaction to clone alleles of target amplicons. The PCR amplicons were gelpurified and subcloned into the pCRII-TOPO vector (Invitrogen, Carlsbad CA, USA). At least 7 clones were randomly selected and sequenced on an ABI3730xl DNA analyzer to ascertain the methylation patterns of each locus. All primers for the bisulfite sequencing analysis are listed in Table 2.
4. Statistical Analysis
All statistical analyses were performed using the STATA 9.2 software package (StataCorp., College Station, TX, USA). The data from most analyses were compared using a t-test, while continuous variables were analyzed using the Mann-Whitney U-test. P-values of less than 0.05 were considered significant.
RESULTS
1. Abnormal Hypermethylation of TSGs in CD Patients
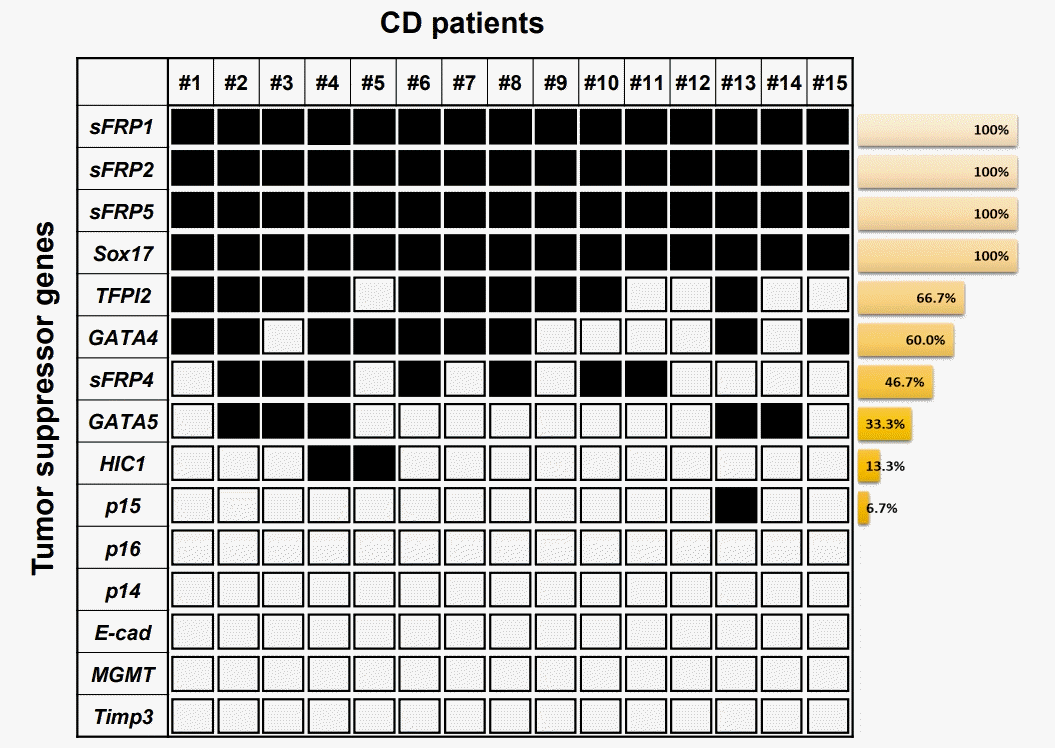
In this study, we investigated transcriptional silencing by promoter hypermethylation of 15 well-known TSG candidates (TFPI2, GATA4, p16, sFRP1, sFRP4, MGMT, Sox17, HIC1, sFRP2, sFRP5, p14, p15, GATA5, E-cadherin, and Timp3) in samples from patients with CD. These genes were previously reported on the basis of comprehensive studies of candidate TSGs in colon cancer, and the transcriptional silencing of these suppressors is regulated by promoter hypermethylation in many cancer types [7]. In addition, highly sensitive DNA methylation markers in colon cancer could be used to detect the risk level of patients with inflammatory diseases, such as UC and CD. Motivated by this possibility, we previously analyzed DNA methylation in UC samples [15,16] and serum samples of CD patients [17]. Based on previous studies, we were aware of that unknown TSGs could be abnormally hypermethylated in patients with CD. Here, we extracted genomic DNA from patient tissues with CD (n = 15) (Table 1). First, using MSP analysis, we assessed the methylation level of the promoter regions of 15 genes (TFPI2, GATA4, p16, sFRP1, sFRP4, MGMT, Sox17, HIC1, sFRP2, sFRP5, p14, p15, GATA5, E-cadherin, and Timp3) (Table 2) in tissue samples from patient with CD. Conventional MSP analysis was performed successfully with most samples. Fig. 1 displays the frequency of hypermethylation for the 15 gene promoter regions in patient tissues with CD. For most of the CD samples we tested, sFRP1, sFRP2, sFRP5, and Sox17 showed the highest hypermethylation levels (100% frequencies), and TFPI2, GATA4, sFRP4, GATA5, HIC1, and p15 showed methylation frequencies of 66.7%, 60.0%, 46.7%, 33.3%, 13.3%, and 6.7%, respectively. The other 5 genes (p16, p14, E-cadherin, MGMT, and Timp3) showed no methylation in the CD samples.
2. Six TSGs Are Highly Methylated in CD Patients
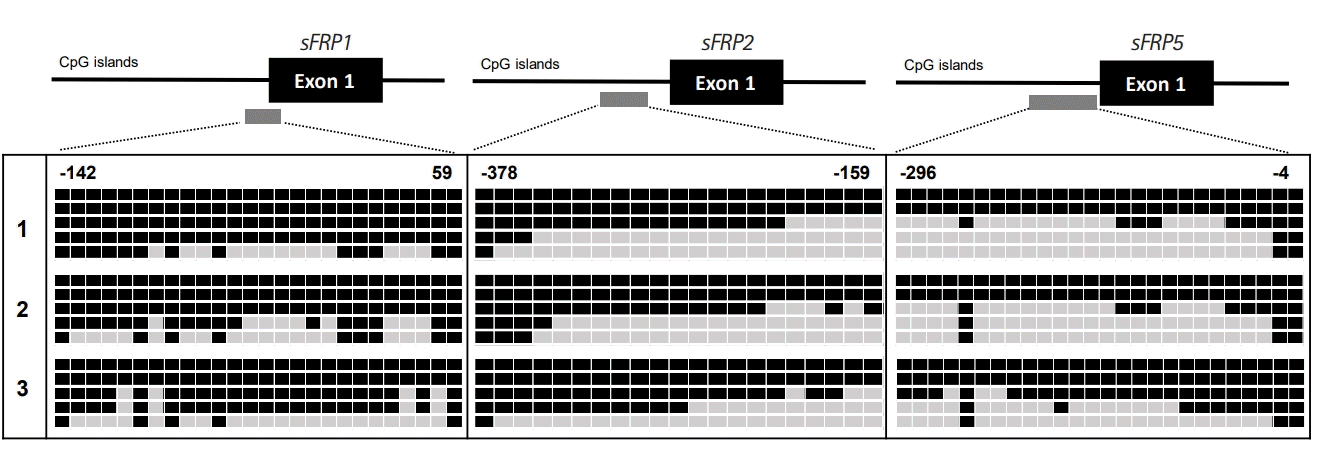
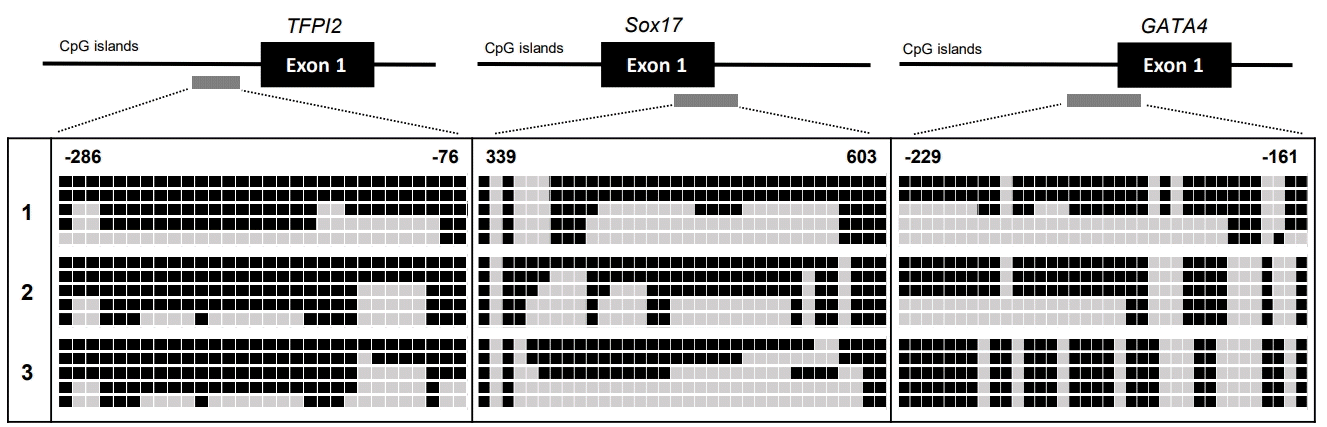
On the basis of the high frequency of methylation of genes (sFRP1, sFRP2, sFRP5, TFPI2, Sox17, and GATA4; > 60% methylation frequencies) in the samples from patients with CD, we next texted the changes in DNA methylation status in the promoter region of the 6 genes by bisulfite genomic sequencing in representative patient tissues with CD (n = 3). All the genes we tested had previously been shown to be hypermethylated in a cancer-specific manner but are not methylated in normal colon tissues [7]. The location and extent of the methylated promoter regions used for bisulfite sequencing, relative to the transcription start sites of exon 1, are as follows: sFRP1: upstream region from –142 to 59, 26 CG sites; sFRP2: upstream region from –378 to –159, 21 CG sites; sFRP5: upstream region from –296 to –4, 26 CG sites; TFPI2: upstream region from –286 to –76, 30 CG sites; Sox17: region from 339 to 603, 34 CG sites; and GATA4: upstream region from –229 to –161, 46 CG sites. The bisulfite sequencing of the individual clones of 5 PCR products from 3 different individual CD tissue samples revealed densely methylated CG sites within the promoter regions of all the clones (Figs 2, 3). The methylation status in the promoter regions of sFRP1, TFPI2, and sFRP2 in the CD tissues showed 82.5%, 70%, and 63.4% methylation at the CG sites, respectively. The CG sites of the Sox17, sFRP4, and GATA4 genes in the CD tissues were methylated in a range from 55% to 57%. Accordingly, the bisulfite sequencing data confirmed the complete methylation of the 6 gene promoters in patients with CD as had been determined by the MSP analysis (Fig. 1); that is, 6 genes were significantly hypermethylated in patient samples with CD. Overall, these results strongly suggest that we found increased DNA methylation levels of the sFRP1, sFRP2, sFRP5, TFPI2, Sox17, and GATA4 genes in inflammatory diseases, as represented by CD.
DISCUSSION
IBD is identified by chronic uncontrolled inflammation of the intestinal mucosa due to abnormal immune response, leading the disruption of the intestinal epithelium [18]. Patients with IBD can be grouped as lifetime risk (2- to 3-fold greater) of developing CRC, and the most important risk factors for colitis-associated cancer are duration, severity and extent of the IBD [13,14]. There is a growing body of evidence suggesting that inflammation is the critical cause of many cancers, and colitis-associated cancer serves as a great model system to study the general principles understanding for the etiology of inflammation-associated cancer [19]. Prevalent evidence supports the theory that IBD is caused by a complex interplay between the predispositions of multiple genes and abnormal interactions with environmental factors. It is becoming increasingly apparent that epigenetic factors can significantly contribute to the pathogenesis of disease.
We have previously reported that the TCERG1L, sFRP1, FBN2, and TFPI2 genes were hypermethylated at high frequency in patients with UC, which is another type of IBD [16]. In the present study, we expanded a gene panel consisting mostly of tumor suppressor candidates identified from the colon cancer and tested the methylation level of these genes in patients with CD. According to our previous data and present study, these TSGs are transcriptionally regulated by promoter hypermethylation in patients with IBD. We do not have direct evidences the biological role of these TSGs during IBD development but our data suggest that epigenetically regulated TSGs could contribute to the pathogenesis of IBD. Although lack of studies have been reported that the methylation of TSGs in CD, here we found the promoter methylation changes of sFRP families, TFPI2, Sox17, and GATA4 genes in patients with CD.
The tissue factor pathway inhibitor 2 (TFPI2) gene is located on chromosome 7q22 and encodes a Kunitz-type serine proteinase inhibitor [20]. TFPI2 can play as a critical roles involved in protecting the extracellular matrix of cancer cells from degradation as well as regulating tumor invasion [21]. Abnormal promoter hypermethylation of TFPI2 has been observed in CRC [22], gastric cancer [23], hepatocellular carcinoma [24], pancreatic cancer [25], and cervical cancer [26]. Moreover, promoter hypermethylation of TFPI2 has been detected in recurrence and early-stage CRC [27], and in sera from CRC patients with large, poorly differentiated carcinomas, invasion, and several metastasis such as lymph node or distant [28]. Overall, our study provides the evidences that promoter hypermethylation of TFPI2 is also detected in most of the CD patient samples we tested. This data suggests that TFPI2 may be a useful biomarker during colonoscopic surveillance screening. Therefore, further studies using a large series of samples are necessary to confirm the hypothesis on the TFPI2 gene could be a promising methylation biomarker for screening patients with CD.
Sox17 encodes an HMG box transcription factor and its biological roles in many different stage of developmental process such as in vascular development and hematopoiesis [29-32]. The Sox17 promoter is hypermethylated in cholangiocarcinoma tissues, lung cancer [33], gastric cancer [34], liver cancer [35], and breast cancer [36]. In addition, inactivated Sox17 by promoter hypermethylation could contribute to the dysregulaion of the Wnt signaling pathway in human cancer [37]. These accumulating data strongly suggests that epigenetically silenced Sox17 can affect to the aberrant activation of Wnt signaling during tumorigenesis in multiple cancer types [38]. We first report here that Sox17 is hypermethylated in patients with CD, suggesting that epigenetically inactivated Sox17 may contribute to the pathogenesis of IBD.
The transcription factor GATA4 plays an essential role in the development and differentiation of the GI tract and is suggested to be involved in CRC development [39,40]. The lack of GATA4 expression regulated by promoter hypermethylation has been reported in primary CRC, gastric, esophageal, lung, and ovarian cancers [40-43]. Although accumulating studies on the GATA4 gene is epigenetically regulated in many cancer types, there is lack of information whether GATA4 gene is hypermethylated in patients with CD.
SFRPs belong to the family of Wnt pathway antagonists, and the transcriptional expression of SFRPs are regulated by gene promoter hypermethylation in colon, gastric, ovarian, and lung cancers [44-47]. Epigenetic regulation of the Wnt pathway was first characterized in colon cancer, which is known to be related to chronic inflammation. Recently, it was shown that the downregulation of SFRP gene family members in gastric and CRC is commonly regulated by promoter hypermethylation [45,48]. Thus, transcriptional silencing of SFRP gene may affect to leading a mechanism of aberrant Wnt signaling activation [45,48]. It has been reported that the frequent inactivation of SFRP1 and SFRP2 in IBD-associated neoplasia further highlights the importance of the canonical Wnt pathway to understand the pathogenesis of IBD-associated cancer. Our study has some limiting issues. We lacked control samples for CD, such that we could not determine the TSG methylation with specificity in CD patient samples. We also lacked clinical information on the CD samples, such as the disease duration or severity that; therefore, we could not implicate the clinical importance of the promoter methylation of TSGs for the CD patients. Therefore, further study is necessary to define the DNA methylation markers in a large series of clinical data from CD patients, including control samples.
In summary, we investigated the promoter DNA methylation pattern of 15 TSGs in samples from patients with CD. We found that samples from 10 of the 15 patients had TSG promoters that were methylated as determined by conventional MSP analysis. Six TSGs (sFRP1, sFRP2, sFRP5, TFPI2, Sox17, and GATA4) showed a high frequency of hypermethylation in the CD patients, a finding that we confirmed by conducting bisulfite sequencing analysis to determine the methylation levels of the genes in the CD patient tissues. To our knowledge, our study demonstrates for the first time that both the Sox17 and GATA4 genes are epigenetically regulated in CD patients and the methylation levels of these genes in samples from CD patients. Although we found that promoter of TSGs is frequently hypermethylated in CD patients, we could not implicate the clinical relevance between promoter methylation of TSGs and CD progression.
 XML Download
XML Download