

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Testosterone is ubiquitous among males and is the most potent naturally secreted steroidal androgenic hormone. Testosterone deficiency, also known as hypogonadism, occurs in males and generally increases with age; obesity, genetic causes, stress, alcohol consumption, and oxidative stress are known to be physiological factors leading to hypogonadism [12]. Hypogonadism results in several diseases such as sexual dysfunction, depression, difficulty concentrating, and muscle weakness [3456]. Serum testosterone levels gradually and progressively decline in people over 40 years of age. Thus, maintaining physiological testosterone levels, especially in men over 40 years of age, has significant health benefits.
Reactive oxygen species (ROS) play an essential role as a second messenger in many intracellular signaling pathways that maintain the microenvironment and cellular homeostasis. Oxidative stress occurs either due to an enhanced ROS production or the downregulated availability of antioxidants. In contrast, an unbalanced oxidative status between ROS generation and clearance by the intrinsic antioxidant defense system may trigger testosterone production in Leydig cells [7]. Moreover, increased ROS levels can lead to DNA damage, endoplasmic reticulum (ER) stress, and apoptosis [89]. However, various antioxidants are documented to have beneficial effects on diverse diseases, including hypogonadism [10].
Natural products provide a valuable source for new drug discovery candidates. Since the newly created chemical compound libraries have limited structural diversity in the therapeutic domain, many researchers are trying to discover therapeutic agents from natural sources [11]. The fruit of Schisandra chinensis (S. chinensis) Baillon, a member of the Magnoliaceae family, is a traditional herbal medicine known to have five flavors (sour, bitter, sweet, salty, and spicy) [1213]. S. chinensis has been used as a tonic, a sedative, and an antitussive medicine in Korea, China, and Japan for over centuries [14]. Several pharmacological studies have demonstrated that S. chinensis has antioxidant, anti-inflammatory, antiviral, anti-cancer, anti-aging, cardioprotective, and hepatoprotective activities [1516]. S. chinensis contains various bioactive dibenzocyclooctadiene lignans such as gomisins (A, B, C, D, E, F, G, K3, N, and J), schisandrol A, schisandrol B, and schisandrin C [17]. The major lignans, viz., schisandrol A and gomisin N, impart diverse pharmacological activities such as hepatoprotective, antiviral, neuroprotective, anti-cancer, anti-inflammatory, and anti-melanogenic [18].
The modulating effects of lignans have recently been linked to cytochrome P450 enzymatic activities. For example, gomisin compounds (A, C, and G) and schisandrin A suppress CYP3A and CYP2C19, respectively [19]. In addition, gomisin A and schisandrin A inhibit CYP3A4 activity [20]. Although it is becoming increasingly clear that lignans play a vital role as antioxidants and improve testosterone production, the molecular mechanism of attenuating hypogonadism still remains unknown. This study evaluates the effects of lignans of S. chinensis fruits, including schisandrol A and gomisin N, on hypogonadism through attenuation of oxidative stress.
Go to : 
MATERIALS AND METHODS
Chemicals and reagents
Schisandrol A, schisandrol B, schisandrin A, gomisin N, schisandrin C, 2,2-diphenyl-2-picrylhydrazyl (DPPH), L-ascorbic acid, potassium persulfate, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), dimethyl sulfoxide (DMSO), and hydrogen peroxide (H2O2) were procured from Sigma-Aldrich (St. Louis, MO, USA). The 2,2’-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) single solution was obtained from Invitrogen (Camarillo, CA, USA). Dulbecco’s Modified Eagle’s Medium (DMEM) and fetal bovine serum (FBS) were purchased from GenDEPOT (Barker, TX, USA). Penicillin-streptomycin and trypsin-ethylenediaminetetraacetic acid (EDTA) were obtained from Gibco (Grand Island, NY, USA). TRI-Solution™ was purchased from BioScience Technology (Daegu, Korea).
Plant material
S. chinensis fruit, identified and confirmed by Professor Hye Kyung Kim, was obtained from the herbal medicinal market (Munkyung, Korea). The voucher specimen (accession number KS-SC-1) was deposited at the Herbarium, College of Pharmacy at Kyungsung University.
Preparation of ethanol extract
Shade-dried S. chinensis fruit (50 g) was pulverized and extracted 2 times with 200 mL of 60% aqueous ethanol for 3 h under reflux. The extracted solutions were filtered through a filter paper (5-μm pore size). The S. chinensis extract (SCE) was concentrated using a rotary evaporator (EYELA N-1000, Rikakikai Co., Tokyo, Japan) under vacuum and lyophilized with a freeze dryer (FD5512; Ilshin, Dongducheon, Korea) to yield a 60% ethanolic extract of SC fruits (SCE). The yield of SCE was 45.6% and was stored at 4°C until use. Prior to use, the extract was dissolved in DMSO to the required concentration.
HPLC analysis of lignans in SCE
Lignans, the major compounds of SCE, were analyzed by HPLC (Fig. 1). The retention times of major peaks in the standard were compared to peaks obtained in the SCE. The HPLC system used for the analysis was a Hitachi Lachrom Elite® HPLC system (Hitachi Instruments Inc., Danbury, CT, USA) equipped with an autosampler and UV detector. Chromatographic separation was accomplished on a Zorbax Eclipse XDB-C18 (250 mm × 4.6 mm, 5 μm) analytical column. Both acetonitrile (A) and water (B) were used as mobile phases in gradient elution mode, and the gradient elution was as follows: 0–10 min, 50% A; 10–40 min, 50–80% A; 40–45 min, 80% A. The flow rate was set at 1.0 mL/min. The column was equilibrated for 10 min before the next injection. The injection volume was 10 μL, and the UV wavelength was 254 nm.

DPPH radical scavenging assay
To investigate the antioxidant activity of SCE, we performed the DPPH radical scavenging assay. Various concentrations of SCE (62.5, 125, 250, 500, and 1,000 μg/mL) were prepared in 1.5 mM DPPH solution in methanol, and the mixtures were incubated at room temperature for 15 min. After the reaction, absorbance was measured at 517 nm using a microplate spectrophotometer (LTEK, Seongnam, Korea). L-ascorbic acid (AA) was used as the positive control. The DPPH scavenging activity was calculated as follows:
ABTS radical scavenging assay
The antioxidant activity of SCE was measured by performing the ABTS radical scavenging assay. To produce ABTS radical cation (ABTS+), the ABTS solution was incubated with 2.45 mM potassium persulfate in the dark at room temperature for 16 h. The ABTS+ solution was diluted with distilled water till it attained absorbance 0.70 (± 0.03) at 734 nm. This solution was then mixed with SCE to prepare varying concentrations of SCE (62.5, 125, 250, 500, and 1,000 μg/mL). Absorbance was measured within 5 min after the reaction; AA was used as the positive control. The ABTS scavenging activity was calculated as follows:
Cell culture and treatment
TM3 Leydig cells were obtained from the Korea Cell Line Bank (Seoul, Korea) and cultured in T75-flasks with DMEM supplemented with 10% FBS and 100 U/mL penicillin-streptomycin at 37°C under 5% CO2. The cell culture media were replaced every 2 or 3 days, and subculture was performed using 0.05% trypsin-EDTA when the cell reached approximately 80% confluency. Exposure to H2O2 is widely used to stimulate oxidative stress in the cell culture system [21]. Therefore, the TM3 cells were exposed to 400 μM H2O2 for 4 hr to promote oxidative stress for further experiments.
Measurement of cell viability
Cell viability was determined using the MTT assay. Briefly, TM3 cells were seeded in a 96-well plate at a density of 1 × 104 cells/well and incubated for 24 h. The culture media were replaced with fresh media containing varying concentrations of SCE, schisandrol A, or gomisin N, followed by incubation for 24 h. MTT solution (5 mg/mL in phosphate-buffered saline [PBS]) was added to each well and incubated for 1 h to measure the cell viability. The media were subsequently removed, and DMSO was added to dissolve the formazan crystals. The absorbance was then measured at 540 nm using Micro Plate Spectrophotometer.
Testosterone enzyme-linked immunosorbent assay (ELISA) assay
Cells were seeded in a 24-well plate at a density of 6 × 104 cells/well and incubated for 24 h, after which the cells were exposed to 400 μM H2O2 for 4 h to induce oxidative stress. Wells were replaced with fresh media containing varying concentrations of SCE, schisandrol A, or gomisin N, and incubated for 24 h. The supernatants were collected for the testosterone ELISA assay, which was performed as per the manufacturer’s instruction (ALPCO, Salem, NH, USA).
Gene expression analysis
To investigate the effects of SCE, schisandrol A, and gomisin N on mRNA expression levels, total RNA was extracted from TM3 cells. The cells were incubated under the same conditions described above for the testosterone ELISA assay. After 24 h incubation with samples, the media were discarded, and cells were washed with PBS. Total RNA was extracted using TRI-Solution™, and RNA concentration was quantified using nanodrop (MicroDigital Co., Ltd., Seongnam, Korea). Gene expression analysis was performed using the One-Step TB Green® PrimeScript™ RT-PCR kit II (TaKaRa, Shiga, Japan), according to the manufacturer’s instruction. Amplification reactions were carried out with the Thermal Cycler Dice™ Real-Time System III TP950 (TaKaRa). Briefly, the reaction mixture was amplified as follows: 95°C for 30 s, 40 cycles comprising 95°C for 5 s, 58°C for 30 s, and 72°C for 30 s. Primer sequences used were as follows: hydroxysteroid dehydrogenase 4 (3β-HSD4) forward 5′-GTGATATGAAGAGGGAGAGGAC-3′, reverse 5′-AAGACAAGTTGGACAGAGTGT-3′; 17-beta-hydroxysteroid dehydrogenase 3 (17β-HSD3) forward 5′-GTTCTCGCAGCACCTTTTTC-3′, reverse 5′-CAGCTTCCAGTGGTCCTCTC-3′; 17, 20-desmolase forward 5′-ACGCTCATCTTCAAGTCAGTAAT-3′, reverse 5′-GGTCTGTATGGTAGTCAGTATCG-3′; β-actin forward 5′-TACTGCCCTGGCTCCTAGCA-3′, reverse 5′-TGGACAGTGAGGCCAGGATAG-3′. The relative gene expression levels were calculated using the 2−(ΔΔCt) method.
Statistical analysis
All data are presented as mean ± standard error of the mean (SEM). Statistical analysis was performed using the GraphPad Prism (version 3.0; GraphPad Software, Inc., San Diego, CA, USA). Statistically significant values were analyzed using one-way ANOVA followed by Dunnett’s multiple comparisons test and considered P-values less than 0.05.
Go to :
RESULTS
Quantification of five lignans in SCE
The calibration curves for five lignans were calculated as a linear relationship between the peak area (y) and concentration (x, μg/mL) of each standard compound and are presented as regression equations (y = ax + b) in Table 1. All calibration curves for the 5 lignans showed acceptable linearity (r2
= 0.9999). The LOD for the five lignans ranged from 0.97 to 1.17 μg/mL, and LOQ ranged from 2.94 to 3.53 μg/mL. The determined, analytical method was applied for the quantification of schisandrol A, schisandrol B, schisandrin A, gomisin N, and schisandrin C in SCE. The contents of these lignans ranged from 3.0 to 19.2 mg/g. The contents of schisandrol A and gomisin N were 19.2 and 14.2 mg/g, respectively. Among these 5 lignans, schisandrol A was the most abundant compound.
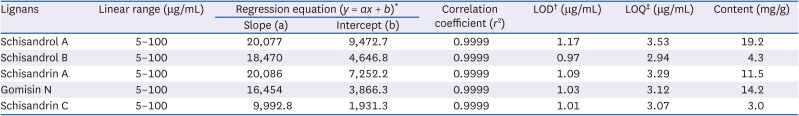
Table 1
Regression equation, linearity, limits of detection, limits of quantification, and content of lignans
LOD, limits of detection; LOQ, limits of quantification; SD, standard deviation.
*y = ax + b, y means peak area and x means concentration (μg/mL).
†LOD: 3.3 × (SD of the response/slope of the calibration curve).
‡LOQ: 10 × (SD of the response/slope of the calibration curve).
![]()
Antioxidant effect of SCE
DPPH and ABTS assays were performed to investigate the antioxidant effect of SCE. DPPH is a stable organic free radical and has been widely used to study the ability of natural products to act as free radical scavengers [22]. As shown in Fig. 2A, SCE (125, 250, 500, and 1,000 μg/mL) dose-dependently scavenges the DPPH radicals, and the scavenging activity was determined to be 19.28%, 45.18%, 78.51%, and 87.88%, respectively. ABTS radical is more reactive than DPPH radical, and can be applied to both the aqueous and organic phases of the extracts [23]. As shown in Fig. 2B, the ABTS radical scavenging activity of SCE (62.5, 125, 250, 500, and 1,000 μg/mL) increased in a dose-dependent manner (14.67%, 25.02%, 46.08%, 75.46%, and 94.79%, respectively). Taken together, these results indicate that SCE has the potential as an antioxidant agent to inhibit oxidative stress.
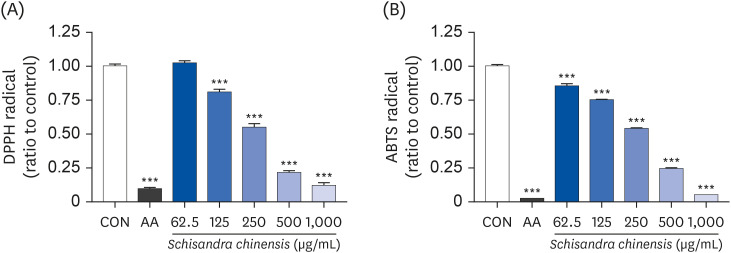 | Fig. 2Antioxidant effect of SCE. (A) DPPH assay and (B) ABTS assay. Data are presented as mean ± SEM, and statistical significance was analyzed by one-way ANOVA followed by Dunnett’s multiple comparisons test.SCE, Schisandra chinensis extract; DPPH, 2,2-diphenyl-2-picrylhydrazyl; ABTS, 2,2’-azino-bis(3-ethylbenzothiazoline-6-sulfonic acid); SEM, standard error of the mean; CON, control; AA, L-ascorbic acid.
***P < 0.001 compared to control.
|
Effects of SCE and its active compounds on cell viability against oxidative stress
We first performed the MTT assay to determine the cytotoxicity of SCE and the active compounds of SCE on TM3 cells. Initially, cytotoxicity of SCE (62.5, 125, 250, 500, and 1000 μg/mL) and the active compounds of SCE (2.5, 10, and 50 μM) were examined on TM3 cells exposed to 400 μM H2O2. As shown in Fig. 3A, the cell viability increased by 7.33% at 250 μg/mL SCE but decreased by 8.66% at 1,000 μg/mL SCE. Although schisandrin A and schisandrin C exerted cell cytotoxicity (data not shown), schisandrol A (Fig. 3B) and schisandrol B (data not shown) did not affect the cell viability. However, exposure to gomisin N increased the cell viability by 2.06% and 7.08% at 10 and 50 μM, respectively (Fig. 3C). Among the three compounds that imparted no cell cytotoxicity, schisandrol A and gomisin N were the most abundant compounds of SCE. Therefore, we selected schisandrol A and gomisin N for further studies to investigate the effect in hypogonadism.
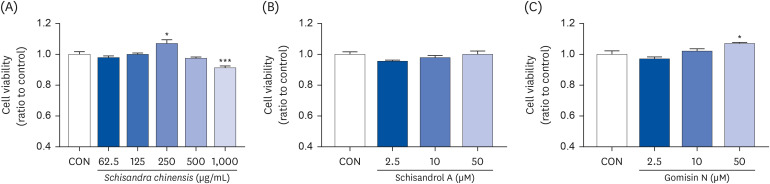 | Fig. 3Cell viability of SCE and its active compounds. (A) SCE, (B) schisandrol A, and (C) gomisin N. Data are presented as mean ± SEM, and statistical significance was analyzed by one-way ANOVA followed by Dunnett’s multiple comparisons test.SCE, Schisandra chinensis extract; SEM, standard error of the mean; CON, control.
*P < 0.05 and ***P < 0.001.
|
Next, TM3 cells were treated with different concentrations of H2O2 for 4 h, and the MTT assay was conducted after 24 h. The cell viability decreased by 4.3%, 22.6%, and 35.28% after exposure to 200, 400, and 600 μM H2O2-induced oxidative stress (data not shown). Following these results, TM3 cells were treated with 400 μM H2O2 to induce oxidative stress for 4 h. The media were replaced with fresh media containing 62.5 or 250 μg/mL SCE and 2.5, 10 or 50 μM schisandrol A or gomisin N for 24 h. The cell viability was observed to recover by 3.02% and 10.22% at 62.5 and 250 μg/mL SCE against H2O2-induced oxidative stress (Fig. 4A). Schisandrol A and gomisin N similarly imparted cell-protective effect of 7.46%, 11.6%, 15.47%, and 9.87%, 14.13%, 11.78%, respectively, as compared to the H2O2 treated group (Fig. 4B and C). These data suggest that SCE and its active compounds, schisandrol A and gomisin N, have the potential to act as antioxidants.
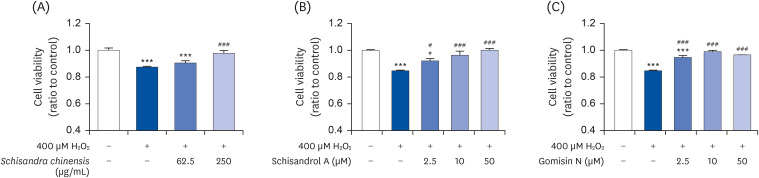 | Fig. 4Effects of SCE and its active compounds on H2O2-induced oxidative stress in Leydig cells. (A) SCE, (B) schisandrol A, and (C) gomisin N. Data are presented as mean ± SEM, and statistical significance was analyzed by one-way ANOVA followed by Dunnett’s multiple comparisons test.SCE, Schisandra chinensis extract; H2O2, hydrogen peroxide; SEM, standard error of the mean.
*P < 0.05 and ***P < 0.001 compared to control. #P < 0.05 and ###P < 0.001 compared to H2O2-treated group.
|
Effects of SCE and its active compounds on testosterone level
To evaluate the effects of SCE and its active compounds on the testosterone level, cells were incubated with 250 μg/mL SCE, 50 μM schisandrol A, or 50 μM gomisin N for 24 h after treatment with 400 μM H2O2 for 4 h. The media were then collected for testosterone ELISA assay. As shown in Fig. 5, the reduced testosterone level after exposure to hydrogen peroxide was recovered by treating with SCE (Fig. 5A), schisandrol A (Fig. 5B), and gomisin N (Fig. 5C). These results indicate the potential of SCE and its active compounds to recover the damage under conditions of oxidative stress.
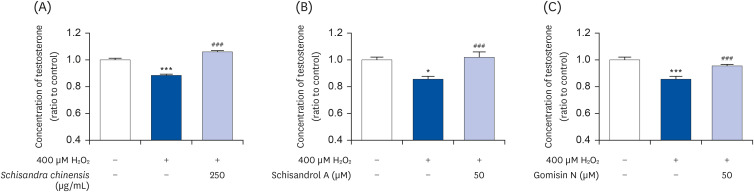 | Fig. 5Effect of SCE and its active compounds on testosterone level under oxidative stress condition. (A) SCE, (B) schisandrol A, and (C) gomisin N. Data are presented as mean ± SEM, and statistical significance was analyzed by one-way ANOVA followed by Dunnett’s multiple comparisons test.SCE, Schisandra chinensis extract; H2O2, hydrogen peroxide; SEM, standard error of the mean.
*P < 0.05 and ***P < 0.001 compared to control. ###P < 0.001 compared to H2O2-treated group.
|
Effects of SCE and its active compounds on gene expression
We investigated the effects of SCE and its active compounds on mRNA expressions of genes involved in testosterone biosynthesis (including 3β-HSD4, 17β-HSD3, and 17, 20-desmolase) under oxidative stress conditions in Leydig cells. As shown in Fig. 6, compared to the H2O2 treated group, exposure to SCE induces the mRNA expressions of 3β-HSD4 (Fig. 6A), 17β-HSD3 (Fig. 6B), and 17, 20-desmolase (Fig. 6C) (49.72%, 70.7%, and 43.03%, respectively). Similarly, compared to the H2O2 treated group, the mRNA expressions of enzymes were significantly induced by 50 μM schisandrol A (41.6%, 41.9%, and 39.45%, respectively; Fig. 6D-F) and gomisin N (49.53%, 32.65%, and 49.11%, respectively; Fig. 6G-I). These data indicate that SCE containing schisandrol A and gomisin N as active compounds have the potential to induce the enzymes involved in testosterone biosynthesis.
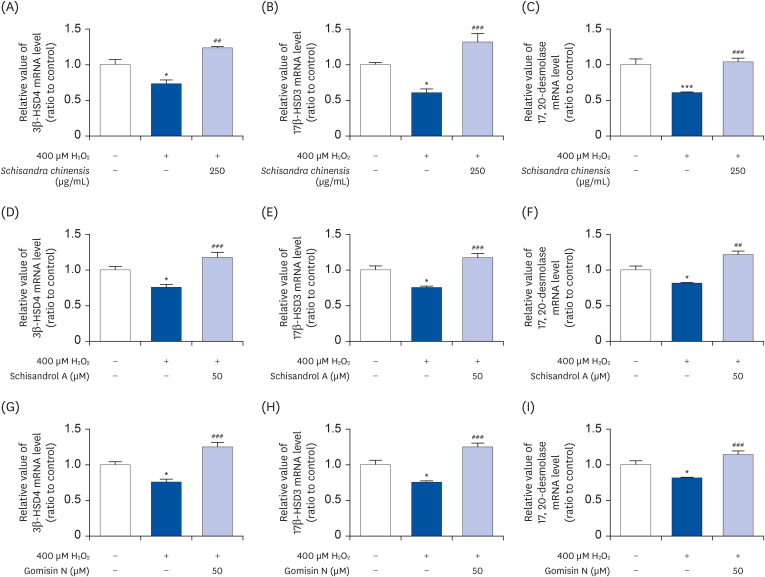 | Fig. 6Effect of SCE and its active compounds on mRNA expressions of 3β-HSD4, 17β-HSD3, and 17, 20-desmolase. (A-C) SCE, (D-F) schisandrol A, and (G-I) gomisin N. Data are presented as mean ± SEM, and statistical significance was analyzed by one-way ANOVA followed by Dunnett’s multiple comparisons test.SCE, Schisandra chinensis extract; SEM, standard error of the mean; H2O2, hydrogen peroxide.
*P < 0.05 and ***P < 0.001 compared to control. ##P < 0.01 and ###P < 0.001 compared to H2O2-treated group.
|
Go to :
DISCUSSION
Testosterone plays an essential role in the quality of life and well-being through controlling the male physiology [24]. Recent evidence has reported the effects of S. chinensis on testicular dysfunction [25]. Numerous natural products with antioxidant properties have been associated with increased testosterone in males, and their compounds have been beneficial in late-onset hypogonadism [2627]. A particular antioxidant, identified as a lignan from S. chinensis, has proven to be a fascinating pharmaceutical or nutraceutical candidate but remains understudied. A recent study identified the major lignans of S. chinensis fruit are schisandrol A, schisandrol B, gomisin N, schisandrin A, and schisandrin C [28]. For the first time, the current study examined the lignans from the S. chinensis fruits, and evaluated whether schisandrol A and gomisin N could be the primary compounds that enhance testosterone production by suppressing oxidative stress. This study also examined the molecular mechanism that alters the expression level of testosterone biosynthesis-related genes such as β-HSD4, 17β-HSD3, and 17, 20-desmolase.
Leydig cells in the testes are the primary source of circulating testosterone levels of the luteinizing hormone produced and secreted by the pituitary gland [29]. Thus, age-associated testosterone deficiency is attributed to impaired function and a reduced number of Leydig cells [30]. Moreover, compelling evidence shows that Leydig cells are sensitive to oxidative stress, which could be a risk factor for decreased testosterone production [3132]. Therefore, the development of compounds capable of inhibiting oxidative stress in Leydig cells is considered a practical approach for treating low testosterone production.
Oxidative stress is involved in severe damage to cells, thereby impairing cellular function and even resulting in cell death through increasing the intracellular ROS or RNS levels to damage cellular microstructures, and activation of the immune and inflammatory systems [3334]. Consistent with previous findings, this study demonstrated that H2O2-induced TM3 Leydig cells resulted in decreased cell viability under oxidative stress. However, following exposure to the total extract of S. chinensis as well as the lignan components of S. chinensis (schisandrol A and gomisin N), cell viability was recovered and oxidative stress levels under H2O2-stimulated conditions were attenuated. In line with previous studies that antioxidants are crucial in cells and protect them against oxidative damage [35], we observed that the total extract, gomisin N, and schisandrol A could directly scavenge ROS, as determined by the DPPH and ABTS assays, thus protecting H2O2-stimulated TM3 Leydig cell death. Recent studies have also supported the finding that SCE suppresses oxidative stress through regulating phase I and phase II detoxification enzymes such as NAD(P)H:quinone oxidoreductase 1 and nuclear factor-like 2 signaling pathway [36]. Therefore, the ability of the SCE to recover cell death seems to be due to its antioxidant effect.
The testosterone production process starts with the oxidative cleavage of the cholesterol side-chain by the cholesterol side-chain cleavage gene (CYP11A) localized in the mitochondrial membrane and converts cholesterol into pregnenolone. The CYP17A1 enzyme in the ER then removes two extra carbon atoms and produces multiple C19 steroids. In addition, the pregnenolone is converted to a weakly active form, such as androstenedione/progesterone, by 3β-HSD4. Finally, testosterone is produced by cleavage of the keto group at the 17th carbon position of androstenedione by 17β-HSD3. Therefore, for a possible therapeutic agent to be a testosterone protector, the compound must have a reverse reaction for attenuation of 3β-HSD4, 17β-HSD3, and 17, 20-desmolase levels. In this study, we observed that the total extract of S. chinensis, gomisin N, and schisandrol A were able to significantly reverse the H2O2-induced suppression of 3β-HSD4, 17β-HSD3, and 17, 20-desmolase expressions. Even though it is already demonstrated that total extract of S. chinensis improved the serum testosterone level in varicocele-induced Sprague-Dawley rats [37], this result established, for the first time, that the major compounds of the fruit (viz., schisandrol A and gomisin N) exert a protective role in the H2O2-stimulated testosterone deficiency by recovering the expression levels of 3β-HSD4, 17β-HSD3, and 17, 20-desmolase.
In conclusion, we demonstrate for the first time that several lignans from S. chinensis, including schisandrol A and gomisin N, have the ability to alter the oxidative stress-induced expression levels of testosterone biosynthesis related-genes such as 3β-HSD4, 17β-HSD3, and 17, 20-desmolase, leading to enhancing the testosterone production. These findings provide the underlying molecular mechanisms of lignans from S. chinensis, and present the possibility of potential therapeutic agents for improving testosterone regulation.
Go to :
 XML Download
XML Download