

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Patients with diabetes mellitus (DM) have a 2- to 5-fold increased prevalence of heart failure (HF),1) and over 40% of acute HF patients have DM.2) Diabetic HF is associated with a higher risk of overall mortality. Although well-controlled DM is associated with a lower risk of long-term mortality than uncontrolled DM, not all hypoglycemic agents are beneficial for HF. In contrast, insulin treatment is associated with an increased mortality compared to those treated with oral antihyperglycemic agents.3) In response to concerns about the increased risk of adverse cardiovascular outcomes associated with certain glucose-lowering therapies, the US Food and Drug Administration and the Committee for Medicinal Products for Human Use of the European Medicines Agency mandated to ensure the cardiovascular safety of new glucose-lowering therapies for treatment of type 2 DM.4) This resulted in a plethora of new cardiovascular outcome trials, presenting major budget issues to develop new drugs. Thus, there is an urgent need for new, simple, rapid, and cost-effective methods to evaluate the effects of hypoglycemic agents on the cardiovascular system, particularly during the early developmental stages.
Recently, zebrafish have been proposed as an excellent vertebrate animal model owing to their high fertility, cost-effectiveness, and physiological similarity to humans.5)6) Particularly, the β-cells of the pancreas and the heart, which are essential organs for DM-HF research, are structurally and genetically similar to those of humans.7)8) Additionally, owing to the rapid development of zebrafish, these organs fully develop within 2–3 days post fertilization (dpf); hence, research can be conducted in a short period. Moreover, compared to other vertebrate models, which die immediately when the oxygen supply is stopped, zebrafish can survive for several days via passive oxygen diffusion, even when blood circulation is blocked. Therefore, zebrafish larvae are widely used in cardiovascular defect studies.9) In zebrafish larvae, which have transparent bodies, cells and organs can be directly observed under a microscope. As heart is a continuously beating organ, it is necessary to observe its contraction to accurately evaluate its function. The beating hearts of live zebrafish larvae can be directly observed via fluorescence microscopy using a transgenic (Tg) zebrafish line expressing the fluorescent protein in the myocardium.10) Therefore, research using zebrafish larvae can provide information that can be easily missed while using the existing vertebrate models.
We have previously reported a zebrafish model of dilated cardiomyopathy induced by brief treatment with terfenadine (TER) and demonstrated its feasibility in screening of new drugs.11)12) Here, we developed and validated a zebrafish model for diabetic HF. We optimized a simple method to induce a DM-like phenotype in zebrafish by treating them with a combination of D-glucose (GLU) and streptozotocin (STZ). Furthermore, we established a novel method and different parameters to evaluate cardiac function after the induction of HF with TER in a DM zebrafish model.
METHODS
Ethical statement
All animal experiments and husbandry procedures were approved by the Institutional Animal Care and Use Committee of the Seoul National University (SNU-200310-1).
Zebrafish maintenance
Adult zebrafish (Danio rerio) were maintained at 26–28°C on a 14/10 hours light-dark cycle in an automatic circulating tank system (Genomic Design, Daejeon, Korea). Zebrafish embryos were raised in egg water prepared by dissolving 60 μg/mL ocean salts (Sigma-Aldrich, St. Louis, MO, USA) in autoclaved deionized distilled water (DDW) at pH 7.0. Zebrafish embryos were euthanized via the low-temperature shock method when the survival rate at 24 hours post fertilization was less than 80%.13) All experiments were performed from 3 dpf of hatched zebrafish embryos to 9 dpf. During treatment with GLU (Sigma-Aldrich), the environmental GLU solution was replaced daily to avoid contamination. We used Tg (myl7:EGFP) zebrafish expressing an enhanced green fluorescent protein (EGFP) in cardiac myosin light chain 7 (myl7)10) and Tg (ins:EGFP) zebrafish expressing EGFP in pancreatic β-cells.14) The Zebrafish Center for Disease Modeling (ZCDM), Korea, provided all Tg zebrafish.
Glucose colorimetric assay
As zebrafish larvae are small, the samples not sufficient to measure blood glucose levels using conventional methods. Therefore, we used an alternative method of measuring glucose in whole-body fluids.7)15) Briefly, 10 larvae per sample were transferred to a 1.5 mL tube and euthanized using the low-temperature shock method. Zebrafish larvae were gently rinsed twice with phosphate-buffered saline (PBS) and homogenized in 100 μL DDW using TissueLyser II (Qiagen, Hilden, Germany). The homogenate was then centrifuged at 13,000 rpm and 4°C for 15 minutes to obtain an eluate containing free glucose. Then, 50 μL of the eluted solution was placed in a 96-well plate and allowed to react with 50 μL of the Amplex™ Red reagent/horseradish peroxidase/glucose oxidase mixed solution, provided in the Amplex™ Red Glucose/Glucose Oxidase Assay Kit (Thermo Fisher Scientific, Waltham, MA, USA), for 30 minutes at room temperature. The absorbance was measured using a microplate reader (VERSAmax; Molecular Devices, San Jose, CA, USA) at an excitation wavelength of 530–560 nm.
Quantitative real-time polymerase chain reaction
For quantitative real-time polymerase chain reaction (qRT-PCR) analysis, total cellular RNA was extracted from ten zebrafish larvae per sample using QIAzol Lysis Reagent (Qiagen). The extracted RNA was reverse-transcribed using the amfiRivert cDNA Synthesis Premix (GenDEPOT, Katy, TX, USA) according to the manufacturer’s instructions. qRT-PCR was performed using cDNA as the template, a SYBR Green PCR kit (GenDEPOT), and the StepOnePlus Real-Time PCR System (Applied Biosystems, Waltham, MA, USA). The expression of all genes was normalized to that of the housekeeping gene 18S ribosomal RNA (18s rRNA) using the 2−ddCt method. The primers used were as follows: ins (forward: 5′-AGT GTA AGC ACT AAC CCA GGC ACA-3′, reverse: 5′-TGC AAA GTC AGC CAC CTC AGT TTC-3′), pck1 (forward: 5′-GAG AAT TCT CAC ACA CAC ACA CGT GAG CAG TA-3′, reverse: 5′-GTA AAA GCT TTC CGC CAT AAC ATC TCC AGC AGA A-3′), nppb (forward: 5′-CAT GGG TGT TTT AAA GTT TCT CC-3′, reverse: 5′-CTT CAA TAT TTG CCG CCT TTA C-3′), Ucn3 (forward: 5′-GAG TGC AGG GCA GAA CAA TGT-3′, reverse: 5′-GAA ACT GGT TGC GCA AAG GA-3′), Slc30a8 (forward: 5′-ATC GTC TTG ATG GAA GGC AC-3′, reverse: 5′-TTT CTC GAA GCA CCT CCT GT-3′), and 18s rRNA primers (forward: 5′-TCG CTA GTT GGC ATC GTT TAT G-3′, reverse: 5′-CGG AGG TTC GAA GAC GAT CA-3′).
Insulin enzyme-linked immunosorbent assay
As with the glucose colorimetric assay above, zebrafish larvae are small and contain very little body fluid, so there are not enough samples to measure circulating or blood insulin levels. Therefore, we measured insulin by homogenizing the whole body of 30 zebrafish larvae per sample. Insulin contents were measured with a Fish Insulin (INS) ELISA kit (MyBioSource, San Diego, CA, USA) according to the manufacturer’s protocol. The absorbance was measured using a microplate reader (Infinite 200 pro; Tecan, Männedorf, Switzerland) at an excitation wavelength of 450 nm.
Measurement of ventricular contractility
Cardiac contractility was estimated as the ventricular fractional shortening (vFS). The Tg (myl7:EGFP) zebrafish larvae were anesthetized with 0.02% tricaine (MS222; Sigma-Aldrich) and then embedded in 3% methylcellulose (Sigma-Aldrich). The beating heart was observed using an automated inverted fluorescence microscope (Leica DMI6000B; Leica Microsystems, Wetzlar, Germany). The hearts of individual zebrafish larvae were imaged continuously for 30 seconds. The vFS was calculated using the ventricular dimension at the end-systole (VDs) and end-diastole (VDd). Calculation formula is as follows:
Measurement of contraction irregularity
Cardiac contraction irregularity was estimated as the standard deviation (SD) of the beat-to-beat interval. Zebrafish larvae anesthetized with 0.02% tricaine were embedded in 3% methylcellulose and positioned such that the dorsal aorta was visible. Blood flow was recorded for 30 seconds using a high-speed camera and the ZebraBlood of the MicroZebraLab system (ViewPoint, Civrieux, France). Subsequently, real-time blood flow pulses were analyzed using ZebraBlood. The blood flow data were used to identify peaks using the findpeaks function of MATLAB (MathWorks, Natick, MA, USA), and the time between the peaks was calculated to determine the beat-to-beat interval. MATLAB was used to set an appropriate threshold to identify the correct peaks.
Motility test and survival analysis
Zebrafish larvae motility was analyzed using DanioVision and EthoVision XT (Noldus, Wageningen, Netherlands). Zebrafish larvae were individually placed in square 96-well plates containing 200 μL egg water, and movements were tracked and recorded for 5 minutes using DanioVision. While monitoring zebrafish larvae motility, they were stimulated with a tapping device once every 30 seconds. Zebrafish motility analysis using EthoVision XT included the assessment of movement distance, velocity, acceleration, and duration.
Kaplan-Meier survival analysis was used for survival analysis. Each zebrafish was transferred to a 96-well plate containing 200 μL egg water. Survival was observed using a microscope every 12 hours until 9 dpf.
Statistical analysis
All data are presented as mean ± SD. Statistical analyses were performed using GraphPad Prism (GraphPad Software, San Diego, CA, USA). Mann–Whitney U-test or Student’s t-test was used for comparing 2 groups or a one-way analysis of variance (ANOVA) followed by the Tukey post-hoc test for compare more than 2 groups. The Kaplan–Meier method with the Log-rank test was used for survival analysis. A p-value of less than 0.05 indicated a significant statistical difference.
RESULTS
Effects of concentrations of D-glucose and/or streptozotocin on the survival rates of zebrafish larvae
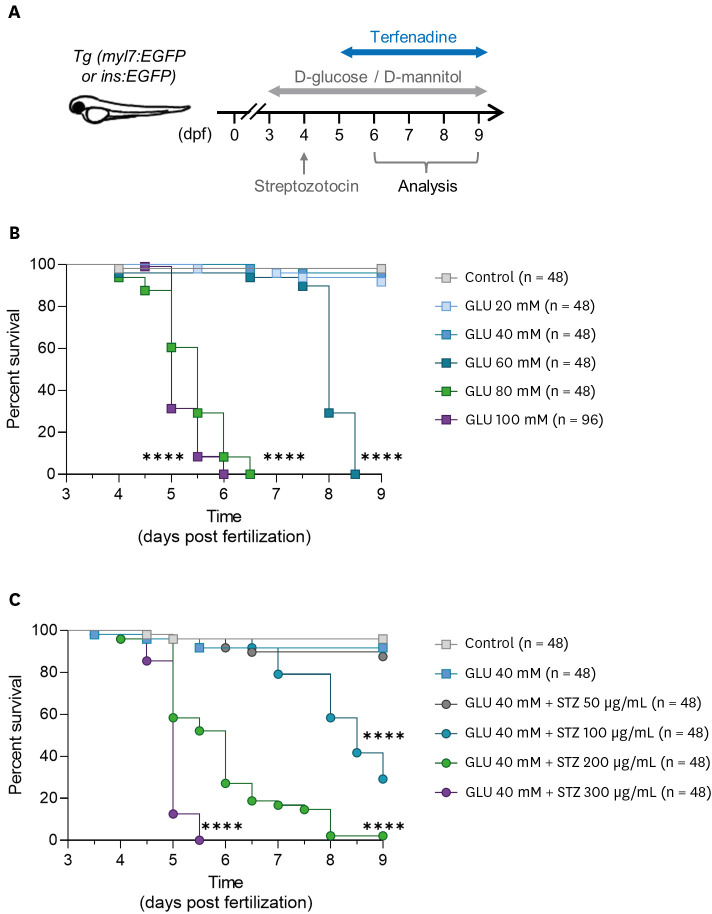
To use the DM zebrafish model for experiments, the baseline survival rate must be stabilized; however, GLU is toxic and can affect survival. Therefore, we evaluated the viability of zebrafish larvae treated with different concentrations of GLU. Because of GLU treatment from 3 to 9 dpf (Figure 1A), immersion in 60–100 mM GLU for over 48 hours had lethal effects on zebrafish larvae, resulting in a sharp decrease in their survival rates (Figure 1B). However, no significant reduction in viability was observed after immersion in GLU concentrations <40 mM (Figure 1B). Viability was assessed in the presence of 40 mM GLU and various concentrations of STZ (Figure 1A). A high STZ concentration significantly reduced the survival rate, whereas 50 μg/mL STZ treatment did not affect the survival rate (Figure 1C).
Figure 1
Effects of various concentrations of GLU and/or STZ on the survival rates and motility of zebrafish larvae. (A) Schematic of the study design. (B) Kaplan-Meier survival analysis of zebrafish larvae after treatment with various concentrations of GLU (n = 48 per group) and (C) a combination of 40 mM GLU and various concentrations of STZ. (n = 48 per group).
GLU = D-glucose; STZ = streptozotocin.
****p<0.0001 vs. (B) control group or (C) GLU 40 mM group.
Induction of hyperglycemia and impairment of glucose homeostasis in zebrafish model with the combination of D-glucose and streptozotocin
DM is characterized by hyperglycemia and impaired glucose homeostasis owing to abnormal insulin expression or function. Therefore, we confirmed the expression of insulin and phosphoenolpyruvate carboxykinase (PEPCK), an essential enzyme involved in gluconeogenesis, in zebrafish treated with GLU/STZ. The same concentration D-mannitol (MAN) was used as an osmotic control for GLU. The expression of ins, a preproinsulin gene, was significantly higher in the GLU-only group than in the control and MAN groups. In contrast, the expression of ins in the GLU/STZ-treated zebrafish was completely suppressed (Figure 2A, Supplementary Table 1). This has also been observed in whole-body insulin contents (Figure 2B). Additionally, the expression of pck1, a PEPCK gene, showed a statistically significant increase in the GLU/STZ group compared to that in other groups, but there was no change in its expression in the GLU or MAN groups (Figure 2C, Supplementary Table 1).
Figure 2
Induction of diabetes mellitus-like phenotypes in zebrafish larvae via combined treatment with GLU and STZ. (A) Relative mRNA expression of ins (n = 8 per group) and (B) Whole-body insulin level (n = 6 per group). (C) Relative mRNA expression of pck1 (n = 8 per group). (D) Whole-body glucose concentration (n = 6–8 per group). (E) Changes in whole-body glucose concentration after high-GLU challenge and (F) the corresponding AUC (n = 5 per group). (G) Relative gene expression of Ucn3 and (H) Slc30a8 (n = 8–11 per group). (I) Representative fluorescence microscopic image of pancreatic β-cells of the Tg (ins:EGFP) zebrafish larvae. Each group had 6–11 samples, with (A, B, and D-H) 10 or (C) 30 larvae per sample. Data are presented as mean ± standard deviation and each dot represents the value of each sample.
AUC = area under the curve; GLU = D-glucose; MAN = D-mannitol; STZ = streptozotocin; Tg = transgenic; EGFP = enhanced green fluorescent protein.
*p<0.05, **p<0.01, ***p<0.001, ****p<0.0001 vs. indicated group.
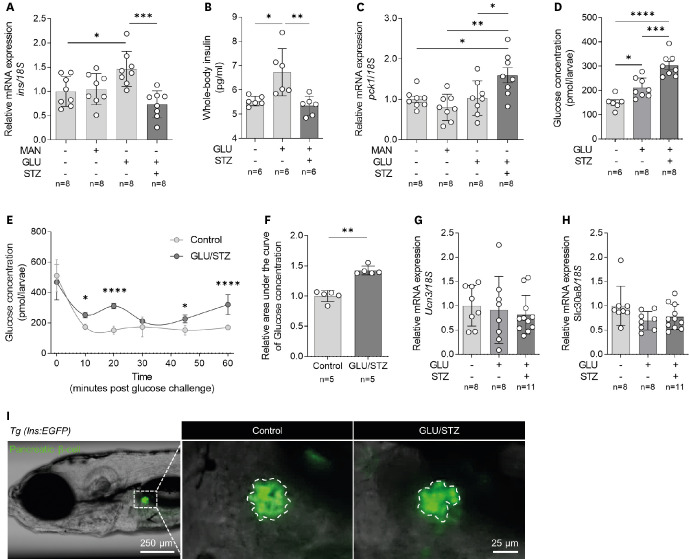
We then measured whole-body glucose concentration as an alternative blood glucose measurement parameter in zebrafish larvae. The GLU/STZ treatment resulted in a significant increase in whole-body glucose concentration compared with that in the control (Figure 2D). The GLU-only group also increased whole-body glucose concentrations, but not as high as GLU/STZ (Figure 2D). Additionally, after the high-glucose challenge, whole-body glucose levels in the GLU/STZ group stabilized more slowly than those in the control group and showed a tendency to fluctuate (Figure 2E and F). Despite the GLU/STZ-induced hyperglycemia and glucose homeostasis abnormalities, there was no significant change in the gene expression of ucn3 and slc30a8, markers of mature pancreatic β-cells (Figure 2G and H, Supplementary Table 1). Also, no change was observed in the morphology of body and pancreatic β-cells of zebrafish larvae (Figure 2I).
Establishment of heart failure with reduced contractility via consecutive treatment with terfenadine post diabetes mellitus induction
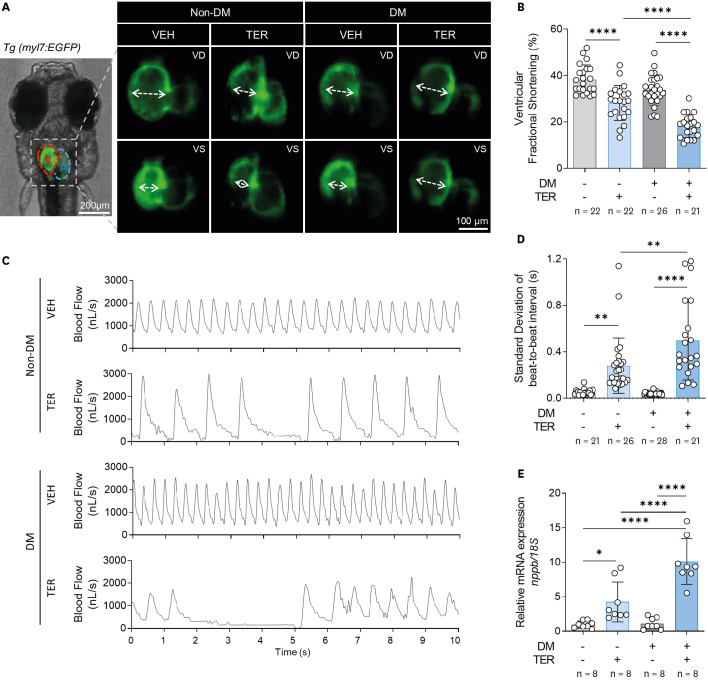
HF with reduced ejection fraction is mainly characterized by reduced cardiac contractility and increased irregular contractions. We previously reported that TER induces HF in zebrafish larvae.11)12) Therefore, we evaluated cardiac contractility and irregular contraction after TER treatment in DM and non-DM zebrafish. First, the hearts of Tg (myl7:EGFP) zebrafish were observed under a fluorescence microscope, and then the ventricle contraction was evaluated (Figure 3A). The vFS was not reduced in the DM zebrafish model; however, TER treatment in non-DM zebrafish slightly reduced the vFS compared with that in the control. Importantly, TER-treated DM zebrafish showed a remarkable reduction in the vFS compared with non-DM zebrafish treated with the same TER concentration (Figure 3A and B). Next, contraction irregularity was evaluated via blood flow analysis. Irregular contractions were observed in both the groups treated with TER but not in the non-DM and DM groups (Figure 3C). Particularly, severe irregular contractions were observed in the TER-treated DM zebrafish (Figure 3C). The SD of the beat-to-beat interval, indicating irregular cardiac contractions, was significantly higher in the TER-treated DM zebrafish than in the TER-treated non-DM zebrafish (Figure 3D). Additionally, the expression of nppb, the gene encoding natriuretic peptide B, an HF biomarker, was also significantly increased in TER-treated DM zebrafish compared to that in other groups (Figure 3E, Supplementary Table 1).
Figure 3
HF phenotype induced by TER in the DM zebrafish model. (A) Representative fluorescent microscopic images of the Tg (myl7:EGFP) zebrafish heart with a GFP at 7 dpf. (B) vFS (n = 21–26 per group). (C) Representative blood pulse graphs. (D) SD of the beat-to-beat interval (n = 21–28 per group). (E) Relative mRNA expression of nppb (n = 8 per group). Each group had 8 samples, with 10 larvae per sample. Data are presented as mean ± SD and each dot represents the value of each sample.
DM = diabetes mellitus; dpf = days post-fertilization; EGFP = enhanced green fluorescent protein; HF = heart failure; myl7 = myosin light chain 7; SD = standard deviation; TER = terfenadine; Tg = transgenic; VD = ventricular diastole; VEH = vehicle; VS = ventricular systole.
*p<0.05, **p<0.01, ****p<0.0001 vs. indicated group.
Evaluation of global motility and viability in the diabetes mellitus-heart failure zebrafish model
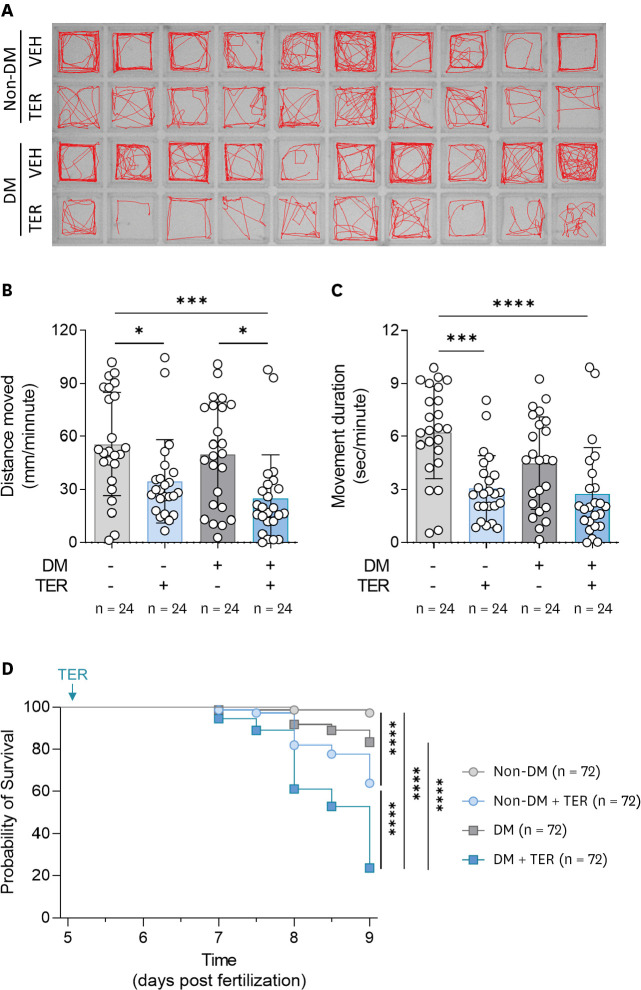
After 24 hours of treatment of non-DM and DM zebrafish with TER, each zebrafish was tracked to evaluate motility (Figure 4A). TER treatment reduced the distance moved and movement duration in both the non-DM and DM groups (Figure 4A-C). Particularly, the TER-treated DM zebrafish demonstrated markedly reduced driving distance compared to the other groups, and the distance moved of TER-treated DM zebrafish was lower than that covered by the TER-treated non-DM zebrafish, but not significant (Figure 4B). Also, no statistical difference was observed between TER-treated non-DM and TER-treated DM zebrafish in terms of the movement duration. (Figure 4C). The survival rate showed a tendency similar to that of motility. Furthermore, a reduction in the survival rate was observed in the TER-treated non-DM zebrafish. However, the survival rates of TER-treated DM zebrafish decreased more remarkable than those of other groups (Figure 4D). In the DM-HF zebrafish model, no morphological changes were observed despite significant reduction in motility and the survival rates (Supplementary Figure 1).
Figure 4
Reduced motility and viability of the diabetic HF zebrafish model. (A) Representative images of motility tracking for 5 minutes in zebrafish larvae. (B) Average moved distance and (C) movement duration per minute (n = 24 per group). Data are presented as mean ± SD and each dot represents the value of each sample. (D) Kaplan-Meier survival analysis (n = 72 per group).
DM = diabetes mellitus; TER = terfenadine; VEH = vehicle.
*p<0.05, ***p<0.001, ****p<0.0001 vs. indicated group.
DISCUSSION
This study presents a method for establishing and evaluating a new animal model for in vivo experiments of DM-HF. The GLU/STZ treatment induced hyperglycemia and abnormal glucose homeostasis in zebrafish larvae, which showed a DM-like phenotype. In addition, TER treatment induced more severe HF in DM zebrafish larvae than that in non-DM zebrafish, as revealed by a remarkable reduction in cardiac contractility and an increase in irregular contraction.
In the previous DM zebrafish larvae models, hyperglycemia was induced by immersion in only GLU.16)17) However, according to the results of our study, immersion in high GLU concentrations resulted in a sharp decrease in the survival rates of zebrafish larvae. This finding indicates that a high-concentration GLU immersion-induced hyperglycemia model is challenging to use in a study requiring a period of 3 days or more and may lead to serious bias in the results. We examined the survival rates of zebrafish at various GLU concentrations and determined an optimum concentration that did not affect viability. STZ destroys pancreatic β-cells through DNA alkylation.18) Our experimental results show that insulin expression is decreased in GLU/STZ-treated zebrafish larvae without affecting the morphology and maturation of pancreatic β cells. It seems that continuous stress caused by GLU and mild damage caused by STZ caused partial dysfunction without destroying β-cells. Here, we determined conditions that partially inhibited pancreatic β-cell function without affecting survival. The GLU/STZ induced-DM-like characteristics did not result in a sharp decrease in the survival rate until at least 9 dpf. In addition, GLU/STZ treatment considerably increased the levels of PEPCK, an essential enzyme in gluconeogenesis, compared with GLU alone. The increase in PEPCK expression suggests that gluconeogenesis was increased due to the lack of glucose required for tissue metabolism despite the high blood glucose levels. Additionally, the GLU challenge experiment revealed that GLU/STZ treatment reduced glucose homeostasis. These results validated the establishment of a new DM zebrafish model with hyperglycemia and abnormal glucose homeostasis.
In this study, DM zebrafish were treated with TER to induce HF. We previously reported a zebrafish larvae HF model induced using TER.11)12) In that study, TER inhibited normal myocardium contraction by blocking the potassium channel, induced cardiomyocyte apoptosis, and consequently reduced cardiac function in zebrafish larvae.11)12) The present study showed that despite treatment with the same TER concentration, more severe cardiac dysfunction was induced in DM zebrafish than in non-DM zebrafish. In DM zebrafish treated with TER, cardiac contractility significantly decreased, irregular contraction remarkably increased; additionally, a sharp increase in the expression of nppb, an HF biomarker, and significant reduction in motility and survival were observed. These results support the results of clinical studies in which patients with DM showed significantly increased HF prevalence and mortality.1)2) Zebrafish larvae models of HF induced by various methods have been reported. Aristolochic acid (AA), a toxin that induces inflammation-mediated HF, is commonly used in zebrafish HF studies;19) additionally, laser-induced HF zebrafish models suitable for cardiac injury and recovery studies are attracting increasing attention.20) In this study, we induced HF using only TER in DM zebrafish. However, depending on the purpose of the study, various methods of HF induction, such as AA or laser injury, can be used.
As research on the prevention and treatment of DM-HF is strongly required, various animal models are being used.21)22)23) However, existing animal models for studying DM-HF have several limitations. As it takes a lot of time to induce DM and HF, induction of DM-HF is more time-consuming. DM induction is mainly induced by a high-fat diet or the β-cell ablation method.24)25)26) However, a high-fat diet requires several weeks to months to induce DM, and symptoms may differ among individuals depending on the feeding state. The β-cell ablation method can induce only the characteristics of type 1 DM. Additionally, conventional induction of HF uses an invasive, technically demanding procedure, and it is difficult to control the symptoms and mortality suitable for the experiment.27) The DM-HF zebrafish model established in this study has a short induction time; can minimize differences among individuals because it does not require a diet; induces hyperglycemia without destroying pancreatic β-cells; and is non-invasive. Therefore, this easy-to-use approach would complement the animal models used in DM-HF studies.
The main limitation of zebrafish DM-HF models is that they are not mammals and have different cardiac circulatory systems than rodents or humans. Therefore, the study results using our DM-HF zebrafish larvae model must be confirmed in rodents and humans. Moreover, chemically induced zebrafish larvae disease models are less stable than genetically modified models. To establish consistent chemically induced models and obtain experimental results, it is necessary to thoroughly control environmental factors, such as circadian rhythm, temperature, population density, and water contamination. Lastly, our DM-HF zebrafish models pertain to HF with reduced ejection fraction and do not apply to HF with preserved ejection fraction, which constitutes approximately 40% of the overall HF cases.
In conclusion, this study established a novel animal model for DM-HF by sequentially immersing zebrafish larvae in GLU/STZ and TER. Additionally, methods for evaluating DM-HF in zebrafish larvae were presented. The results of this study can be used for in vivo studies of DM-HF and utilized as a platform to assess the effects of new medications for DM and HF.
 XML Download
XML Download