

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Sarcopenia, a generalized disorder of skeletal muscle, is commonly found in the elderly due to a decline in muscle mass.1 It is known that muscle mass shows decline every year after the age of 30. Such decline is associated with an increased risk of other disorders such as decreased activity, hormonal changes, and digestive disability.2345 Previous research studies have revealed that sarcopenia has multiple contributing factors, including inflammatory pathway, adiposity, and chronic diseases.678 In addition, sarcopenia is heavily influenced by external variables such as diet, exercise, and lifestyle.91011
Recently, scientists are interested in genetic factors that influence skeletal muscle traits for evaluating the heritability of sarcopenia. Several detailed gene-targeted linkage analyses have been performed for insulin like growth factor 1 (IGF-1),12 myostatin (MSTN),13 activin A receptor type 1B (ACVR1B),14 and so on. Variants contributing to the genetic influence on skeletal muscle traits have also been studied. For example, angiotensin-converting enzyme (ACE) has been investigated in many polymorphism studies,151617 although several studies have reported that there are no significant on muscle traits.181920 Alpha-actinin-3 (ACTN3)2122 and vitamin D receptor (VDR)232425 are well known factors associated with muscle strength for possessing multiple polymorphisms.
Many researchers have performed genome-wide association studies (GWAS) to find genetic risk factors of a specific disease. Various single nucleotide polymorphisms (SNPs) have already been found in sarcopenia.2627 SNP is a single base pair mutation in a DNA sequence that can affect gene function and regulation. Although many sarcopenia associated SNPs have been discovered in Caucasians,627 candidate SNPs and related genes in Asians have not been well established yet.
Therefore, the purpose of this study was to evaluate genetic correlation according to muscle mass in middle aged Koreans using data from the Korean Genome and Epidemiology Study (KOGES), a large population-based genomic cohort study.
METHODS
Study subjects
Epidemiological and genomic data sets in this study were from Ansan and Anseong cohorts of the KOGES conducted by National Research Institute of Health, Centers for Disease Control and Prevention, Ministry for Health and Welfare, Republic of Korea. Baseline participants were 10,030 Koreas aged 40 to 69 years who were from Ansan or Anseong of Gyeonggi-do (province), South Korea. Among them, 9,351 subjects with laboratory data available were included in this study.
To identify sarcopenia associated variants, those in the top 30% and bottom 30% of muscle mass index (MMI) were compared. A total of 7,452 people with an MMI of 30–70% were excluded. A total of 1,004 people were also excluded due to missing data. Finally, 895 people were selected for the demographic study. Of these 895 people, 48 were not included in the SNP survey and 67 showed outlier in GWAS analysis. Finally, 780 subjects were included for the GWAS analysis.
Measurements of lifestyle and comorbidities
All participants attended a community clinic for clinical assessments at each follow-up visit. Body mass index (BMI) was calculated as weight in kg divided by the square of the height in meters. Weight was obtained for participants in light clothing and barefoot. Waist and hip circumference were also measured. The remaining survey items consisted of drinking & smoking status, the level of education, and monthly income. We also obtained their history of hypertension, diabetes, gastritis/stomach ulcer, allergy, myocardial infarction, thyroid disorder, congestive heart failure, coronary artery disease, hyperlipidemia, asthma, chronic lung disorder, peripheral vascular disease, kidney disease, various tumors, cerebrovascular disease, head trauma, urinary tract infection, gout, degenerative arthritis, and rheumatoid arthritis.
Study genotypes
Genotyping of the cohort population was performed using Affymetrix Genome-Wide Human SNP array 5.0 (Affymetix Inc., Santa Clara, CA, USA). The Korea Biobank Array generated 500,568 SNPs for this dataset. We used P values for selecting significant SNPs between people in the top 30% and people in the bottom 30% of MMI. We additionally queried alleles, minor allele frequencies (MAF), and associated genes via Single-Nucleotide Polymorphism database (dbSNP) at the National Center for Biotechnology information (www.ncbi.nlm.nih.gov/projects/SNP/). We used ALFA data for MAF containing data of subjects from 12 diverse populations, including Asian, African, European, and others.
We assessed whether candidate variants were related to specific gene expression in other various tissues by expression quantitative trait loci (eQTL) analyses using a database of the genotype-tissue expression (GTEx) project. Among them, genes associated with sarcopenia or muscle-related diseases by eQTL analysis were identified. Other variants known to be polymorphism or disease associated variants of identified genes were found through dbSNP. We then performed an interactive heatmap matrix of pairwise linkage disequilibrium (LD matrix) statistics to find out if variants we found were related to known variants using LD link (https://ldlink.nci.nih.gov/).
Statistical analysis
Continuous data are reported as mean ± standard deviation. Categorical data are presented as number (%). To find any significant differences in baseline characteristics and clinical factors between people in the top 30% and people in the bottom 30% of MMI, an unpaired t-test was performed for continuous variables showing normal distribution. Otherwise, Wilcoxon’s rank-sum test was performed. Difference in proportion between people in the top 30% and people in the bottom 30% of MMI was tested using χ2 test for categorical variables. If the assumption of χ2 test did not meet, Fisher’s exact test was performed. PLINK and R software 4.1.0 (R Core Team, R Foundation for Statistical Computing, Vienna, Austria, 2020) were used to conduct statistical analysis (version 1.9). The significance level was set at P < 0.05. The PLINK software was used to filter out SNPs that were irrelevant. Genotyping call rate (0.05) was used as the SNP filtering parameter to exclude missing genotypes of SNPs. Significant SNPs between people in the top 30% and people in the bottom 30% of MMI are presented by a Manhattaplot using qqman package version 0.1.8 of R software.
RESULTS
General characteristics for demographic study
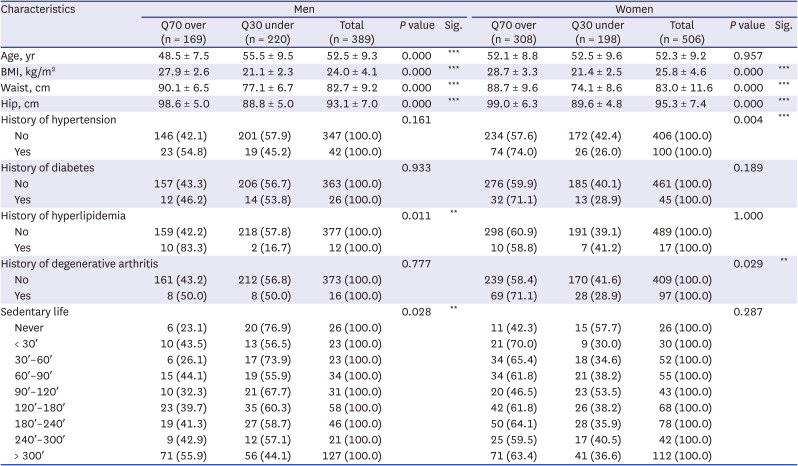
When subjects were divided into the top 30% and bottom 30% of MMI, top 30% people had 169 men and 308 women and bottom 30% people had 220 men and 198 women. In men, statistically significant differences in age, BMI, waist, hip, history of hyperlipidemia, and sedentary life status were observed between top 30% and bottom 30% of MMI groups. Particularly, age, BMI, waist and hip showed significant differences between the two groups with P value less than 0.005. In women, age, BMI, waist, hip, and hypertension history also showed significant differences between top 30% and bottom 30% of MMI groups (Table 1). Results for all demographics are presented in Supplementary Table 1.
Table 1
Demographic study containing significant results
GWAS study
GWAS results are presented with a Manhattan plot (Fig. 1). There were 13 significant SNPs in the male group and 14 significant SNPs in the female group. QQ plots also showed GWAS results against expected association results between people in the top 30% and people in the bottom 30% of MMI (Fig. 2). All results of significant SNPs containing related genes are shown in Table 2. All results of SNPs of the present study are represented in Supplementary Table 2. Genes associated with variants in the male group identified using the dbSNP were LRP1B containing rs11679458 and RGS6 containing rs11848300. Pi4K2A was a gene associated with variants in the female group. It also contained rs1189312 as a variant. Published studies about identified genes are presented in Table 3.
Fig. 1
Manhattan plot of each group classified by gender. (A) Manhattan plot in the men group, (B) Manhattan plot in the women group.
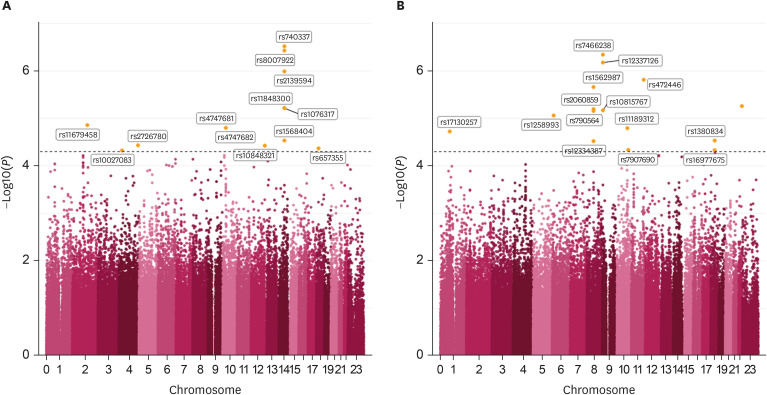
Fig. 2
Q-Q plot of each group classified by gender. (A) Q-Q plot in the men group. (B) Q-Q plot in the women group.
Q-Q = Quantile-Quantile.
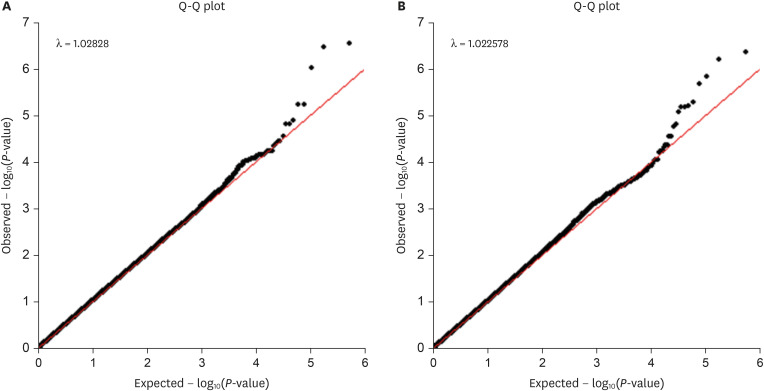
Table 2
SNPs results of GWAS data containing related genes
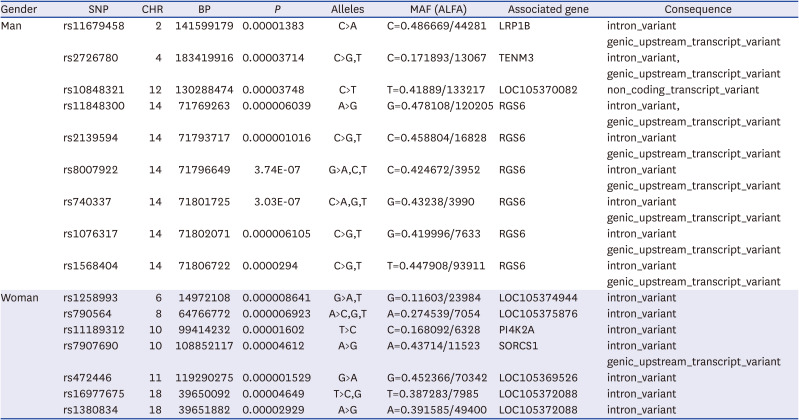
Table 3
Summary of muscle atrophy related genes

| Gender | Associated SNP/gene | Associated studies | Summary of study |
|---|---|---|---|
| Men | rs11679458/LRP1B | Tanaga et al.34 | The proliferation-dependent expression of LRP1B may influence SMC migratory activity via modifying PDGF and uPA signals. |
| rs11848300/RGS6 | Ahlers-Dannen et al.35 | RGS6 expression is observed in muscular and connective tissues around epithelial cells | |
| Women | rs11189312/PI4K2A | Simons et al.36 | Pi4k2 knockout mice with no detectable kinase activity have no evident phenotype when they are young. Later on, they developed tremors, spastic gait, muscle weakness, and feeding issues, which exacerbated as they grew older. |
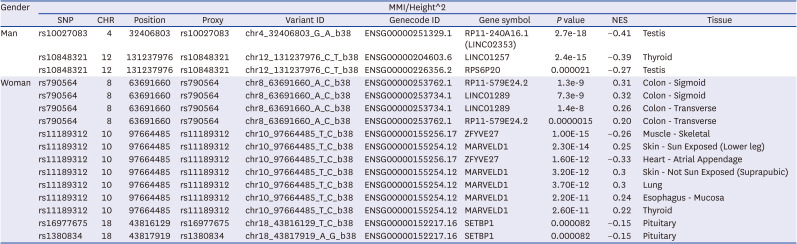
We also conducted eQTL analysis to assess whether candidate variants were related to specific gene expression in other various tissues. As a result, rs10027083 and rs10848321 in men and rs790564, rs11189312, rs16977675, and rs1380834 in women were eQTL in each gene. Among them, only rs11189312 in women was eQTL of ZFYVE27 in skeletal muscles. It showed very significant results. Therefore, other variants associated with ZFYVE27 and skeletal muscle related disease were investigated using dbSNP. Finally, a total of three polymorphisms (rs10882883, rs17108378, and rs35077384) were identified, including one disease (spastic paraplegia) related polymorphism. We also conducted LD matrix statistics to determine whether rs11189312 was related to those variants (Fig. 3). The LD heatmap plot was prepared using reference population of East Asian. The result showed that rs11189312 had little relevance to other variants. Rs11189312 had R squared values of 0.001, 0.018, and 0 with D prime values of 0.247, 0.231, and 0.037 against rs17108378, rs10882993, and rs35077384, respectively. All significant results of eQTL analysis in GTEx containing related genes are shown in Table 4. All eQTL analysis results are presented in Supplementary Table 3.
Fig. 3
Interactive heatmap matrix of pairwise linkage disequilibrium statistics in rs11189312 and ZFYVE27 associated SNPs. (A) All pairwise R square statistics. (B) All pairwise D prime statistics. (C) Interactive heat map of associated SNPs.
SNP = single nucleotide polymorphism.
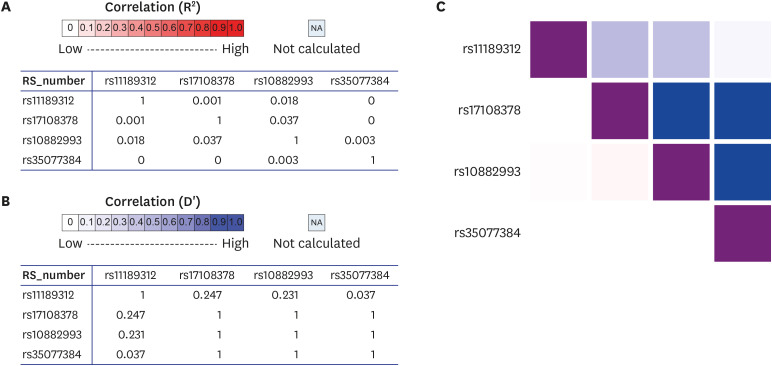
Table 4
eQTL analysis in GTEx containing related genes
DISCUSSION
In the present study, demographic analysis was conducted based on MMI adjusted by square of height. Age, BMI, waist and hip showed significant results in men. These are obesity-related factors associated with muscle loss.28 In particular, significance was found in people who had a history of asthma disease. As people age, their respiratory muscle mass decreases, their respiratory muscle strength weakens, and their respiratory function deteriorates.29 Several studies have consistently revealed the association between respiratory diseases (such as asthma) and sarcopenia or muscle loss.2930 In addition, the MMI adjustment value tended to be low in people who had a sedentary life. In women, there were significant differences in BMI, waist and hip. However, there was no significant difference in age. In particular, there was a significant difference in people with a history of degenerative arthritis. Muscle wasting as a natural part of aging has recently been proven in individuals with OA.3132 It has been suggested that muscle wasting has a direct impact on joint stability and that loss of mobility can lead to articular cartilage degeneration.33
GWAS was performed to determine the relationship between muscular study and each SNP associated genes. In men, rs11679458 was an intron variant of LRP1B. It has been suggested that proliferation-dependent expression of LRP1B may influence SMC migratory activity by modifying platelet-derived growth factor (PDGF) and urokinase-type plasminogen activator (uPA) signals.34 Rs11848300 is an introgenic variant of RGS6 because its expression is observed in muscular tissues around epithelial cells.35 In women, rs11189312 was an intronic variant of PI4K2A. One study has shown that Pi4k2 is associated with aging-related muscle weakness.36 Pi4k2 knockout mice without detectable kinase activity had no evident phenotype when they were young. However, when they grew older, they developed tremors, spastic gait, muscle weakness, and feeding issues that exacerbated as time went by.36
Sarcopenia is a well-known aging-related disease. Recent studies have suggested that neuromuscular junction (NMJ) degeneration progresses with the onset of sarcopenia during aging.3738 NMJ is a complicated synapse that connects muscle fibers to motor neurons. Its damage and morphological change gradually occur during aging, causing muscle paralysis or weakness.39 Spastin is an ATPase-containing domain that can interact with microtubules.40 According to one study, knockdown of spastin localized at the NMJ of Drosophila results in synaptic undergrowth.41 In vitro studies have shown that spastin can regulate stabilization of microtubules to maintain activity.4243 In addition, previous research studies have shown that mutation of spastin could result in hereditary spastic paraplegia, a neurological disorder with progressive spasticity and weakness of leg muscle.444546 Therefore, patients with genetic factors that can induce mutation in spastin could be congenitally vulnerable to muscle loss and growth.
SNP rs11189312 has not been reported in association studies. However, our study revealed that rs11189312 could affect ZFYVE27 gene. ZFYVE27, also known as SPG33, belongs to the FYVE-finger family of proteins responsible for regulating endocytic membrane trafficking.474849 According to one study, ZFYVE27 can interact with endogenous spastin whose mutation is the most common cause for hereditary spastic paraplegia.47 In our study, eQTL analyses using GTEx showed rs11189312 in skeletal muscle for ZFYVE27 with a P value of 1.00E-15. As rs10882993, rs17108378, and rs35077384 were reported SNPs of ZFYVE27 in dbSNP database, we conducted LD matrix statistics to identify whether rs11189312 was associated with those reported SNPs. Our results only showed a slight correlation. Therefore, rs11189312 could be a novel variant affecting ZFYVE27 expressed mainly in skeletal muscles. This indicates that rs11189312 could lead to muscle disability or damage by affecting ZFYVE27 function. It might also be directly associated with sarcopenia because people in the bottom 30% of MMI showed more significant results than those in the top 30% of MMI in the present study.
In the present study, people in the bottom 30% of MMI group were vulnerable to muscle loss. Thus, nutrition and exercise should be paid attention to. Since GWAS analysis focuses on identification of SNPs as biomarkers for specific diseases, early screening of high-risk groups for sarcopenia and clinical treatment will be required.50 In addition, results of present study including SNPs and genes can be used in the development of new drugs. Therefore, genetic results from a GWAS study could lead to clinical significance through research and development of biomarkers.
In conclusion, the present nation-wide study in Korea had the largest size reported for identifying sarcopenia associated factors. Demographic study and GWAS were performed by dividing participants into a group with a high muscle mass and a group with a low muscle mass based on MMI adjusted by square of height. Particularly, rs11189312 might be associated with sarcopenia. It was a novel discovery in our study. Based on our variant results, further study is needed to determine the association between sarcopenia and ZFYVE27 which might play an important role in spastin.
The present study has some limitations. First, there was no restriction on compositions of participants due to cohort research. Second, sarcopenia screening was not conducted using tests such as handgrip test. Third, a further study of newly identified variants is needed for evaluating their genetic influence on the onset of sarcopenia.
 XML Download
XML Download