

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
The prevalence of gestational diabetes mellitus (GDM) is increasing worldwide, and this may be related to older maternal age at pregnancy and obesity.12 GDM can cause various perinatal and fetal complications, such as preeclampsia, macrosomia, and fetal distress, and affects the short- or long-term risk of type 2 diabetes and cardiovascular disease of the mother even after delivery.2345 In normal pregnancy conditions, insulin resistance increases according to fetal growth, and the beta cells of the pancreas promote cell proliferation and secrete more insulin in response to the insulin requirement. GDM is characterized by the inability of pancreatic beta cells to respond adequately to increased insulin resistance, resulting in hyperglycemia of various degrees.6 While treatments with lifestyle and pharmacological interventions have demonstrated short-term benefits, the long-term impact of an intervention, genetic and environmental mechanisms of GDM, and its complications remain unclear.7
Pregnancy is a process in which the mother suppresses various rejection reactions to maintain the semi-allogenic fetus in an immune-tolerant state, and the placenta is an essential organ for maintaining pregnancy.8 Since localized mechanisms may contribute to fetal evasion of maternal immune attack, the induction of maternal immunosuppression against maternal alloreactive reaction is an important process for maintaining maternal-fetal tolerance.9 Evidence that disrupted homeostasis of proinflammatory and anti-inflammatory T cells is associated with preeclampsia or recurrent miscarriage,10111213 supports the impact of innate immunity in pregnancy. Several alterations in the immune system have also been observed in patients with GDM. These include increased serum levels of pro-inflammatory cytokines, which can mediate monocyte and macrophage inflammation in the adipose tissue,14 and the patients with GDM presented reduced levels of regulatory T cells (Tregs) and elevated levels of serum interleukin 6 (IL-6) and tumor necrosis factor alpha.15 Increased total number of lymphocytes was observed in peripheral blood mononuclear cells (PBMCs) of patients with GDM,16 and a decrease in immune-escape molecule, cytotoxic T cell antigen-4 (CTLA-4), and an increase in the activation CD4 markers and CD8 were observed.17 A recent study using single-cell RNA sequencing (scRNA-seq) confirmed the infiltration of immune cells in the GDM placenta and cell-to-cell interaction.18 However, they could not validate the precise characteristics of T cell subsets due to the lack of immune cell sorting processes.
Accumulating evidence suggested the critical role of immune checkpoint to sustain immunological hemostasis under pregnancy status.19 The inhibitory signals provided by ligation of (programmed cell death-1 (PD-1) with its ligands programmed death-ligand 1 (PD-L1) and PD-L2 lead to maintenance of the necessary homeostasis between T-cell activation.20 Aberrant PD-1 signaling was involved in pregnancy associated complications, such as preeclampsia or miscarriage.21 Moreover, low PD-1 expression in patients with T1DM associated with increase T-cell proliferation and activation, leading to destruction of β cells.22 However, the role of immunoescape signaling in GDM are not fully understood.
This study aimed to identify the changes and contribution of the placenta in the GDM based on RNA-seq. We found changes in various immune-related signaling pathways in GDM placentas, and investigated the characteristics of T cell subsets and immune checkpoint molecules by flow cytometry (fluorescence-activated cell sorting [FACS]) analysis in the placenta of patients with GDM compared to controls.
METHODS
Human specimens
Twenty-seven patients with GDM and 27 normal pregnant women were recruited from Chungnam National University Hospital. All patients with GDM were diagnosed according to the American Diabetes Association guidelines.23 Additionally, we used the following enrolment criteria: singleton, age < 40, and not complicated by other diseases. Pregnant women with body mass index (BMI > 30 kg/m2) or those undergoing insulin treatment were excluded. There was no participant with gestational hypertension, preeclampsia, proteinuria, or history of polycystic ovary syndrome. Nine cases (six GDM and three controls) were used for RNA-seq, and other cases were used for validation experiments for FACS analysis of placental mononuclear cells (MNCs), immunohistochemical analysis, real-time polymerase chain reaction (PCR), and western blot analysis. Placenta tissue was collected from the subjects during cesarean delivery. Each of the four placenta quadrants was sampled approximately 1.5 cm away from the umbilical cord attachment from the fetal side of the placenta. Fetal membranes and visible large vessels were removed, and phosphate-buffered saline (PBS) was used to wash placenta samples before separating into maternal- and fetal-side samples.
RNA extraction for sequencing and library preparation
First, the maternal placentas were collected en bloc as 1 × 1 cm pieces for transcriptomic analysis. Then, the tissue samples were homogenized using a mortar and pestle, and total RNA was extracted using the easy-spin Total RNA Extraction Kit (iNtRON, Seongnam, Korea) following the manufacturer’s protocol. All experiments were then conducted under clean conditions, and the equipment was pre-autoclaved. The quality of the extracted RNA was evaluated using an Agilent 2100 Bioanalyzer RNA Nano Chip (Agilent Technologies Inc., Santa Clara, CA, USA). The extracted RNA was used to construct RNA libraries using the TruSeq Stranded mRNA Sample Prep Kit v2 (Illumina, San Diego, CA, USA), according to the manufacturer’s protocol. The quality was analyzed using an Agilent 2100 Bioanalyzer and an Agilent DNA 1000 kit. Finally, the samples were sequenced using an Illumina HiSeq2500, which yielded an average of 38 million paired-end 100 nucleotide reads.
Transcriptomics analysis
The reads were aligned to the UCSC Homo sapiens reference genome (GRCh37/hg19) using TopHat2 v2.1.5. The default TopHat parameter options were used. To analyze the differentially expressed gene (DEG) profiles between the compared groups (Control vs. GDM), the Tuxedo protocol was used.24 The aligned reads were processed using Cufflinks v2.2.1, which is based on the fragments per kilobase of exon model per million reads mapped (FPKM), and unbiased, normalized RNA-seq fragment counts were used to analyze the relative transcript levels. Gene transfer format (GTF) files were generated to quantitatively compare the transcript levels in each sample to those in a reference GTF file. Next, we used Cuffdiff to calculate the differences in the FPKMs between each group. Heat maps were generated using PermutMatrix Version 1.9.3 (LIRMM, Montpellier, France).
GEO dataset analysis
The GSE154377 dataset was analyzed using GEO RNA-seq Experiments Interactive Navigator (GREIN, http://www.ilincs.org/apps/grein/). The counts table was normalized to the gene level and downloaded to draw Supplementary Fig. 1. Differential expression analysis was performed using GREIN to compare gene expressions of GDM vs. control.
Isolation of placental MNCs
First, the placentas were collected at the time of delivery and immediately washed with isotonic sodium chloride solution. The placental villi were scraped from the chorionic membrane using a scalpel. Single-cell suspensions were obtained by homogenizing placental samples in Dulbecco’s phosphate-buffered saline (DPBS) (Welgene, Daegu, Korea), and digested with 5 mL of RPMI digestion solution in a gentleMACS dissociator (Miltenyi Biotec, Bergisch Gladbach, Germany) according to the manufacturer’s protocol, and then filtered using a 70 μm nylon mesh (BD Falcon, Frankin Lakes, NJ, USA). Placental MNCs were isolated by centrifugation on a Ficoll-Paque density gradient (GE Healthcare Life Science, Buckinghamshire, UK) at room temperature. After centrifugation, the MNC layer was collected and washed with DPBS (Welgene). The isolated MNCs were resuspended in DPBS (Welgene) containing 0.5% BSA and 0.05% sodium azide. Trypan blue dye exclusion testing was used to determine the number of viable cells in the suspension, which was used for flow cytometry analysis.
FACS analysis of placental MNCs
As described previously,25 placental MNCs were incubated with fluorochrome-conjugated monoclonal antibodies for 40 minutes at 4°C. MNCs were pre-incubated with an anti-human BD Fc blocker (BD Pharmingen, San Diego, CA, USA), followed by staining with the live/dead marker anti-FVD-APC-Cy7 (eBioscience, San Diego, CA, USA). The antibodies used in this study were anti-CD3-PerCP-Cy5.5, anti-CD3-PE-Cy7, anti-CD4-AF700, anti-CD8-PE, anti-CD8-APC, anti-CD28-APC, anti-CD45RA-FITC, anti-CD45RO-PE-Cy7, anti-CD57-FITC, and fixable viability dye-APC-Cy7 (all supplied by eBioscience). MNCs were stimulated with phorbol-myristate acetate/ionomycin/brefeldin A/monensin for 5 hours. Cells were fixed and permeabilized using a Fixation/Permeabilization Buffer kit (eBioscience). The permeabilized cells were washed and resuspended in 1% formaldehyde and further stained for intracellular cytokines with anti-interferon gamma (IFN-γ)-PE-Cy7 and anti-IL-17A-APC. Multicolor flow cytometry was performed using a BD LSRFortessa flow cytometer (BD Biosciences, San Jose, CA, USA), and data were analyzed using FlowJo software (FlowJo, LLC, Ashland, OR, USA).
Immunohistochemical staining analysis
PD-1 and PD-L1 immunohistochemical staining was performed on the placental tissues of 68 patients, including control and patients with GDM. Whole tissue sections were cut from representative placental paraffin blocks using a microtome and mounted onto coated slides, which were then transferred to Ventana (Tucson, AZ, USA) and Dako (Glostrup, Denmark) automated immunostainers. Staining was performed according to the manufacturer’s protocol using anti-PD-L1 (Ready-to-Use, clone SP263; Ventana) and anti-PD-1 (NAT105, 1:100; Cell Marque, Rocklin, CA, USA) antibodies. Tonsil tissues were used as the controls. The immunostaining pattern of PD-L1 in the placenta was diffusely expressed on the outer surface of syncytiotrophoblasts of the chorionic villi. The cytotrophoblastic cells and villous stroma were negative, and the intermediate trophoblastic cells were very weak. The pattern of PD-L1 expression was membranous, without cytoplasmic or nuclear expression. PD-L1 was expressed diffusely on the syncytiotrophoblasts of the placenta, and only the intensity of the immunostaining was scored and categorized as weak, moderate, or marked. PD-1 immunohistochemistry (IHC) in the same placentas was evaluated and expressed on immune cells. PD-1 positive immune cells were scattered at the implantation sites of the placenta. The number of positive cells was counted per 50 high-power fields. PD-1 scores were categorized as “Negative (positive cells/50 HPF < 1), Weak (positive cells low (1 ≤ positive cells/50 HPF < 5) and Moderate (5 ≤ positive cells/50 HPF).”
Real time PCR
Real time PCR was performed as described previously.26
The primer sequences were: PDCD1-F: 5′-CGTGGCCTATCCACTCCTCA-3′, PDCD1-R: 5′-ATCCCTTGTCCCAGCCACTC-3′; PD-L1-F: 5′-AAATGGAACCTGGCGAAAGC-3′, PD-L1-R: 5′-GATGAGCCCCTCAGGCATTT-3′; PD-L2-F: 5′-GGACCCACAGAATTCCTTCA-3′, PD-L2-R: 5′-GGCAACCCATCTGTTTCTGT-3′; CTLA-4-F: 5′-CAGGGAAGTTTTGTGGAGGA-3′, CTLA-4-R: 5′-CCAAAGCACATGTCAACACC-3′; IFN-γ-F: 5′-AGCTCTGCATCGTTTTGGGT-3′, IFN-γ-R: 5′-CGCTACATCTGAATGACCTGC-3′.
Western blot analysis
Cells were lysed in buffer containing 150 mM NaCl, 1.0% Nonidet-P 40, 0.5% sodium deoxycholate, 0.1% sodium dodecyl sulfate, 50 mM Tris, pH 8.0, and a protease inhibitor cocktail (Roche Applied Science, Vienna, Austria). Electrophoresis was performed as described previously.27 The following primary antibodies were used for western blot analysis: anti-phospho-STAT1 (S727) (1:1,000; Cell Signaling Technology, Danvers, MA, USA), anti-total STAT1 (1:1,000; Cell Signaling Technology), anti-phospho-STAT3 (S727) (1:1,000; Cell Signaling Technology), anti-phospho-STAT3 (T705) (1:1,000; Cell Signaling Technology), anti-total STAT3 (1:1,000; Cell Signaling Technology), anti-phospho-PI3K-p85 (1:1,000; Cell Signaling Technology), anti-PI3K-p85 (1:1,000; Cell Signaling Technology), and anti-β-actin (1:1,000; Cell Signaling Technology). Following incubation with the corresponding horseradish peroxidase-conjugated secondary antibodies (1:1,000; Santa Cruz Biotechnology, Dallas, TX, USA), immunoreactive bands were visualized using enhanced chemiluminescence detection. software (Bio-Rad, Hercules, CA, USA).
Statistical analysis
The results were expressed as mean ± standard deviation (SD). Chi-square and paired t-tests were used to compare clinicopathological data. P value (< 0.05) were used to select genes differentially expressed between the two groups. All in vitro experiments were repeated three times, and statistical significance was analyzed using the two-tailed Student’s t-test or one-way analysis of variance followed by Tukey’s post hoc test. Data are expressed as mean ± SD, and a value of P < 0.05 was deemed statistically significant (P < 0.05). SPSS statistical software for Windows, version 20 (SPSS, Chicago, IL, USA) was used for all statistical analyses.
RESULTS
The placenta of GDM group at the terminal pregnancy exhibit various inflammatory signaling using transcriptomics analysis
Transcriptomics analysis were performed in 6 GDM patients and 3 control participants. The clinical information of the participants enrolled in the study is described in Fig. 1A. Compared to the control group, we identified 758 significantly differentially expressed genes (DEGs) (P value < 0.05) between GDM placentas and controls (Supplementary Table 1). Volcano plot analysis revealed the significant DEGs with fold change ≥ 1.5 and P value < 0.05, such as SCN3A, ACTC1, ASBS2, APOB, and CXCL9 (Fig. 1B). The heatmap analysis revealed the distinct gene expression patterns between two groups (Fig. 1C). Gene ontology analysis based on DEGs revealed the upregulation of diverse mitochondria related pathways, including oxidative phosphorylation, ATP synthesis, and oxygen transport (Fig. 1D). Interestingly, the placenta of GDM exhibited down regulation of various immune related signaling pathways, including regulation of lymphocyte activation, immune effector process, T cell differentiation, and immune response signaling pathway. Collectively, these data suggest that altered T cell immunity related with GDM in pregnant women.
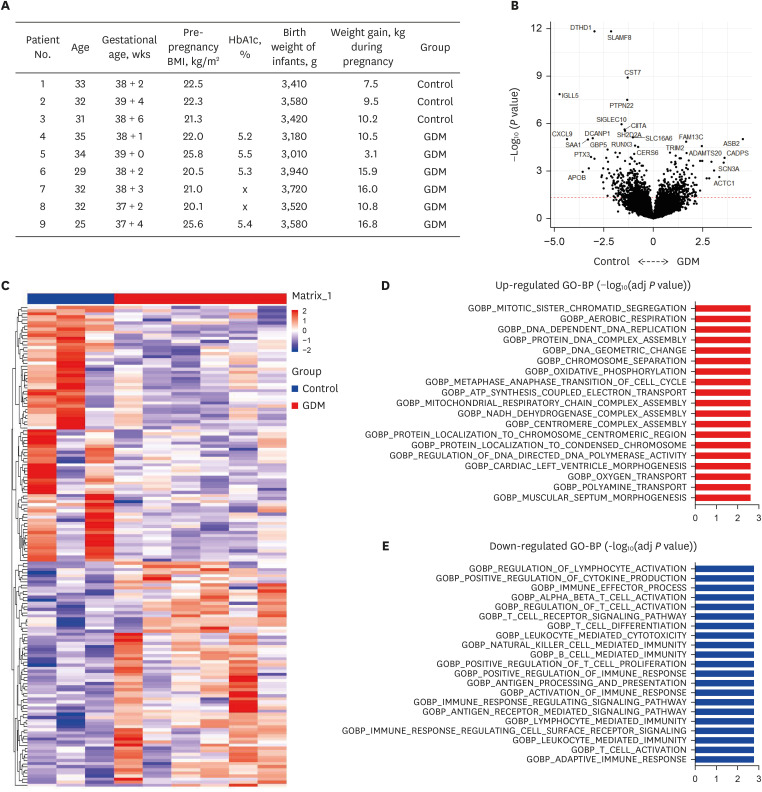
Fig. 1
Clinical information and differentially expressed canonical pathways in comparison between GDM patients and controls. (A) Clinical information including age, BMI, glycosylated hemoglobin (HbA1c), BMI, birth weight of infants, and weight gain. (B) DEGs in comparison between GDM patients and controls by volcano plot analysis. (C) Heatmap analysis of DEGs. (D) Upregulated differentially expressed canonical pathways in comparison between GDM patients and controls. (E) Downregulated differentially expressed canonical pathways in comparison between GDM patients and controls.
GDM = gestational diabetes mellitus, BMI = body mass index, HbA1c = hemoglobin A1c, DEG = differentially expressed gene.
![]()
Population of memory CD4+ T cells and proinflammatory cytokine-producing CD4+ T cells increased in placenta of GDM compared with control
To identify the immunophenotype of T cells in the placenta of women with GDM, we first investigated the frequency and activation status of T cells from the pregnant women’s placenta with or without GDM. Six patients with GDM and eight controls were included in the FACS analysis. Various clinical parameters of participants are described in Table 1. The GDM group demonstrated higher pre-pregnancy BMI and BMI at delivery, compared to controls; however, there was no significant difference between two groups. Four patients among the GDM group (66.7%) revealed the post-partum complications, whereas, no such complication were encountered in the control group (Table 1). Of the four patients who experienced complications, one patient experienced both maternal and fetal complications, including preterm labor and premature fetus; two patients experienced maternal complications; and one patient experienced a fetal complication of congenital genitourinary tract anomaly (Table 1).
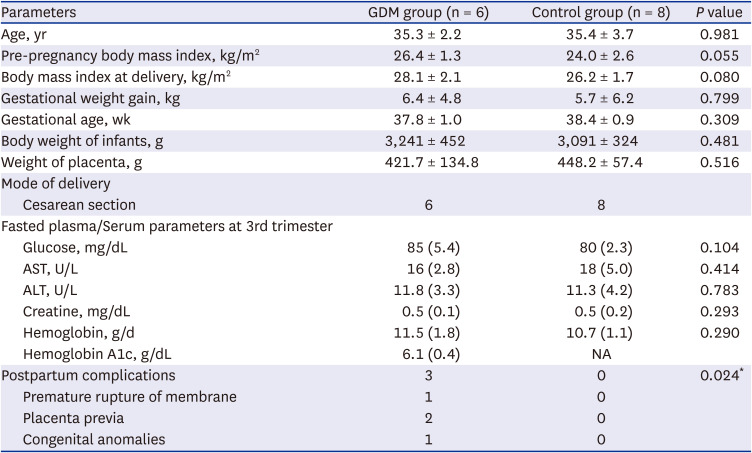
Table 1
Various clinical parameters of total patients in fluorescence-activated cell sorting analysis (N = 14)
Data are given as mean ± standard deviation and values in brackets are percentages (%). The χ2 test and Independents-Samples t-test were used to evaluate the significance between two groups.
GDM = gestational diabetes mellitus, AST = aspartate aminotransferase, ALT = alanine aminotransferase, NA = not available.
*P < 0.05 compared with the corresponding controls.
![]()
Placental MNCs from normal controls and patients with GDM were first gated single cells and lymphocytes, and the lymphocyte population was then further analyzed for uptake of a fixable viability dye. To investigate the phenotypic characterization of CD4+ T cells, we evaluated the frequency of CD45RA+CD45RO− and CD45RA−CD45RO+ among CD4+ T cells in the placental MNCs from normal controls and patients with GDM. The naïve CD4+ T cell population, CD45RA+CD45RO−, tended to decrease, along with an increased population of memory CD4+ T cells and CD45RA-CD45RO+ CD4+ T cells in patients with GDM (Fig. 2A). The population of IFN-γ+CD4+ T cells was also significantly higher in patients with GDM (Fig. 2B). We also analyzed the population of Th17 T cells using IL-17A+CD4+ T cells between the GDM and control groups. There was a significant increase in the population of IL-17A+CD4+ T cells in the placental MNCs of patients with GDM (Fig. 2C). These data suggest that patients with GDM have a larger population of memory CD4+ T cells and proinflammatory cytokine-producing CD4+ T cells in the maternal placenta.
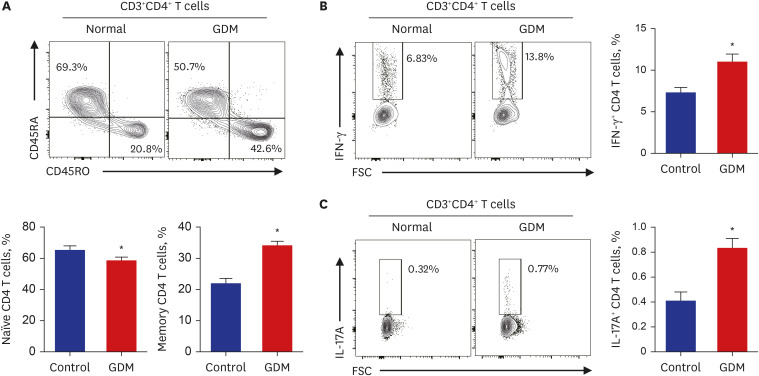
Fig. 2
Immunophenotype of CD4+ T cells of the maternal placenta in pregnant women with and without GDM. (A) Representative flow cytometry plots are presented for CD45RA and CD45RO expression by CD4+ T cells in patients with GDM (n = 6) and normal controls (n = 8). Statistical analysis of the population of CD45RA+CD45RO− (naïve) or CD45RA−CD45RO+ (memory) T cells in CD4+ T cells in the two groups. (B, C) The frequency of IFN-γ- and IL-17A-secreting cells in the population of CD4+ T cells was compared between the two groups. Data are expressed as mean ± standard error of the mean. Flow cytometry plots are representative of all independent experiments.
GDM = gestational diabetes mellitus, IFN = interferon, IL = interleukin, FSC = forward scatter.
*P < 0.05 compared with the corresponding controls.
![]()
The population of memory CD8+ T cells and proinflammatory cytokine-producing CD8+ T cells increased in the placenta of patients with GDM than the control group
Next, we analyzed the phenotypic characteristics of CD8+ T cells. The frequency of CD45RA+CD45RO− and CD45RA−CD45RO+ among CD8+ T cells and the IFN-γ+ expression in CD8+ T cells in the placental MNCs from normal controls and patients with GDM were investigated. Comparison of the phenotype CD4+ T cells in the placenta between GDM and controls showed that the naïve CD8 T cell population, CD45RA+CD45RO−, tended to be decreased, along with the increased population of memory CD8+ T and CD45RA−CD45RO+ CD8+ T cells in patients with GDM (Fig. 3A). The expression of IFN-γ in CD8+ T cells was significantly higher in pregnant women with GDM than in controls. These findings indicate a larger quantity of memory CD8+ T cells and proinflammatory cytokine-producing CD8+ T cells in the placenta (Fig. 3B).
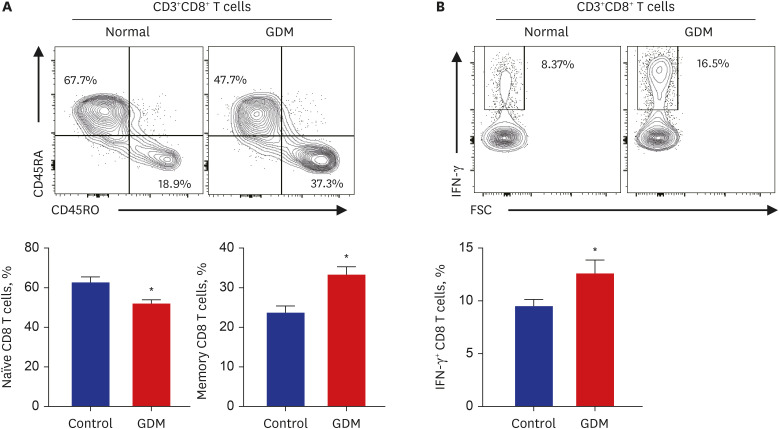
Fig. 3
Immunophenotype of CD8+ T cells of the maternal placenta in pregnant women with and without GDM. (A) Representative flow cytometry plots are presented for CD45RA and CD45RO expression by CD8+ T cells in patients with GDM (n = 6) and normal controls (n = 8). Statistical analysis of the population of CD45RA+CD45RO− (naïve) or CD45RA−CD45RO+ (memory) T cells in CD8+ T cells in the two groups. (B) The frequency of IFN-γ-secreting cells in the population of CD8+ T cells was compared between the two groups. Data are expressed as mean ± standard error of the mean. Flow cytometry plots are representative of all independent experiments.
GDM = gestational diabetes mellitus, IFN = interferon, IL = interleukin, FSC = forward scatter.
*P < 0.05 compared with the corresponding controls.
![]()
Senescent CD8+ T cells are more numerous and CD279 (PD-1) dominantly expressed in placenta of GDM
A previous study reported that senescent T cells have the ability to secrete abundant proinflammatory cytokines in diverse metabolic diseases.28 Moreover, our transcriptomic analysis revealed upregulation of T cell exhaustion signaling in GDM. For these reasons, we investigated the functional phenotypic changes in senescent CD4+ and CD8+ T cells from the placenta of patients with GDM and controls. Among CD4+ T cells, the CD28−CD57+ senescent population tended to be larger in patients with GDM, but this difference was not statistically significant (Fig. 4A). The population of CD28−CD57+CD8+ T cells and senescent CD8+ T cells were significantly increased in the placenta of the GDM group (Fig. 4B). We also analyzed the population of CD279 (PD-1)-positive senescent T cells, since PD-1 is precise molecules related immune-escape signaling. Although the CD279+CD28-CD57+ population tended to be larger in the GDM group without statistical significance (Fig. 4C), CD279 (PD-1) was predominantly expressed in the placenta of patients with GDM in senescent CD8+ T cells (Fig. 4D), indicating that there are more immunosenescent CD8+ T cells, which predominantly express CD279 (PD-1) in the GDM placenta group than in the control group. To validate these results in RNAseq, we analyzed the expression of PD-1, PD-L1, and PD-L2 in patients with GDM and controls; however, there was no expression value for PD-1 and no significant difference between PD-L1 and PD-L2 (Supplementary Fig. 1A). Notably, in GSE154377, a study focusing on the role of the placenta in GDM, PD-L1 expression decreased significantly in the first trimester of pregnancy and tended to increase in the third trimester (Supplementary Fig. 1B). These results suggest that immune-escape signaling has different effects on the placenta in early and late pregnancy and also provides a potential explanation for the increase in PD-1 in senescent T cells in this study.
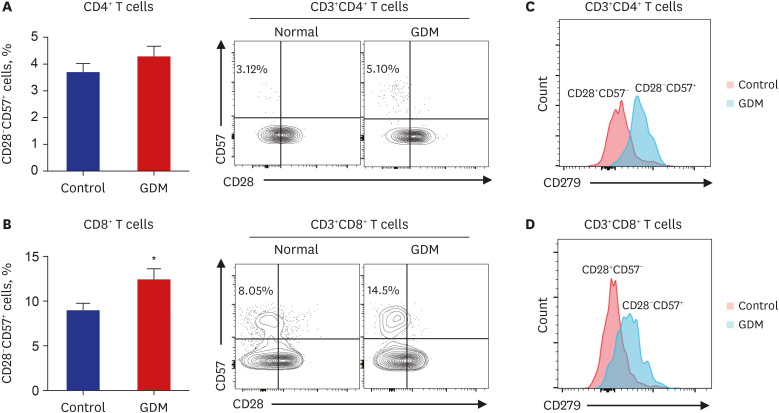
Fig. 4
Population size of CD28−CD57+ senescent T cells from normoglycemic subjects and patients with GDM. (A, B) Representative flow cytometry plots are presented for CD57 and CD28 expression by CD4+ or CD8+ T cells in patients with GDM (n = 6) and normal controls (n = 8). Statistical analysis of the population of CD28− and CD57+ T cells in CD4+ and CD8+ T cells in the two groups. (C, D) CD279 (PD-1) expression of placental CD4+ and CD8+ T cell subsets from patients with GDM and normal controls. Data are expressed as mean ± standard error of the mean. Flow cytometry plots are representative of all independent experiments.
GDM = gestational diabetes mellitus, PD-1 = programmed cell death-1.
*P < 0.05 compared with the corresponding controls.
![]()
PD-L1 immunoreactivity was more intense and mRNA expression of immune-check point molecules were increased in the placenta of patients with GDM
Accumulating evidence suggests the significance of immunological tolerance in the placenta in GDM pathophysiology29; however, the precise role of PD-1/PD-L1 signaling has not been clearly demonstrated. Since we first identified the upregulated PD-1 expression in senescent T cells in the maternal placenta of patients with GDM, we validated the expression of PD-1/PD-L1 in maternal placentas using immunohistochemical analysis. A total of 61 participants were enrolled in the IHC analysis (Table 2). The GDM group demonstrated significantly more increased gestational weight gain and decreased gestational age, compared to the control group (Table 2). As results of the FACS analysis, postpartum complications were significantly increased in patients with GDM, compared to the control group. PD-L1 immunoreactivity was significantly detected in the syncytiotrophoblasts (Fig. 5A). PD-L1 immunoreactivity was significantly more intense in the syncytiotrophoblasts of chorionic villi in the placenta of patients with GDM (Fig. 5A, Supplementary Fig. 2A). PD-1 expression was detected in some decidua lymphocytes (Fig. 5B, Supplementary Fig. 2B). We defined the absence of PD-1 positive cells as negative, the presence of 1 to 4 cells as weak, and the presence of 5 or more cells as moderate in the 50 high-power field. PD-1 expression showed a tendency to increase in GDM group compared to in the control group (Fig. 5B). These findings indicate that the higher intensity of PD-L1 expression is consistent with increased PD-1+ senescent CD8+ T cells in the maternal placenta of patients with GDM. Moreover, the mRNA expression of PD-1, PD-L1, and CTLA-4 was significantly increased in placentas of patients with GDM (Fig. 5C). Since we hypothesized that the PD-1/PD-L1 signaling in the placenta is dysregulated in GDM based on our transcriptomics and FACS data, we investigated STAT signaling, a known regulator of PD-L1 expression in other tissues. There were more patients with increased phosphorylation of STAT1, phosphorylation of STAT3, and phosphorylation of PI3K in placentas from patients with GDM, even though the expression patterns of PD-1-PD-L1-STAT-PI3K were not consistent and there were differences between individuals (Fig. 5D, Supplementary Fig. 2C). These data suggest that immuno-escape signaling is dysregulated in the placenta of patients with GDM, although the interrelationship and mechanism could not be elucidated.
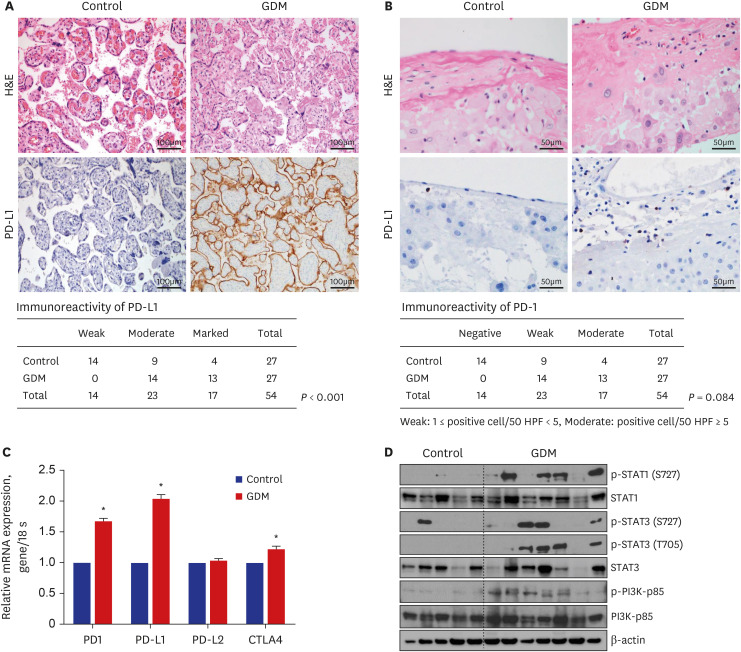
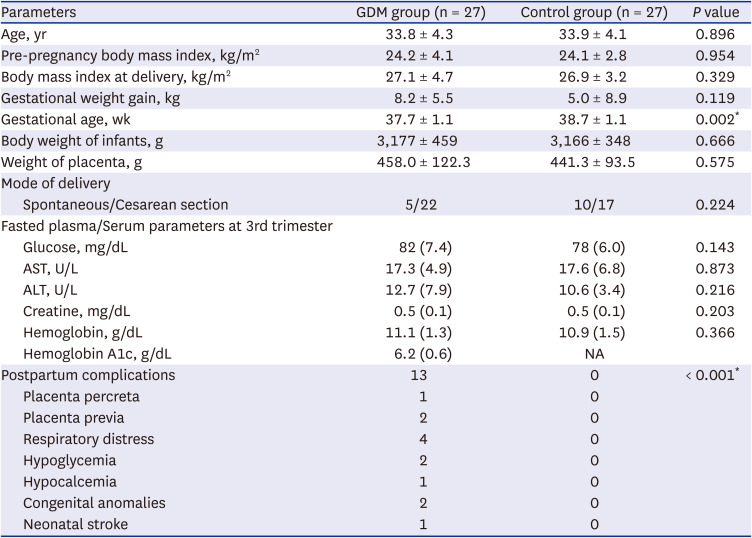
Fig. 5
Dysregulated immune-escape signaling in placenta from GDM participants. (A) Representative images of H&E statin and PD-L1 expression between control and GDM. PD-L1 were scored as weak, moderate, and marked. Scattered PD-L1 positive lymphocytes in implantation site of decidua. (B) Representative images of H&E statin and PD-1 expression between control and GDM. PD-1 were scored as negative, weak, and moderate. The number of PD-1 positive lymphocytes were counted per HPF. (C) mRNA expressions of genes related with PD-1, PD-L1, PD-L2, and CTLA-4 between two groups (Control vs. GDM). (D) Representative images of Western blot analysis about p-STAT1, STAT1, p-STAT3, STAT3, p-PI3K, PI3K. Comparison of densitometry of blot images between two groups (Control vs. GDM).
H&E = hematoxylin and eosin, PD-L1 = programmed death-ligand 1, GDM = gestational diabetes mellitus, PD-1 = programmed cell death-1, HPF = high-power field, CTLA-4 = cytotoxic T cell antigen-4.
*P < 0.05 compared with the corresponding controls.
![]()
Table 2
Various clinical parameters of total patients in immunohistochemistry analysis (N = 54)
Data are given as mean ± standard deviation and values in brackets are percentages (%). The χ2 test and Independents-Samples t-test were used to evaluate the significance between two groups.
GDM = gestational diabetes mellitus, AST = aspartate aminotransferase, ALT = alanine aminotransferase, NA = not available.
*P < 0.05 compared with the corresponding controls.
![]()
DISCUSSION
In order to identify the changes and contribution of the placenta in GDM, we evaluated T-cell subsets and immune checkpoints in the placenta of patients with GDM through RNA-seq and FACS. We demonstrated changes in various immune-related signaling pathways and altered pro-inflammatory T cells, senescent T cells, and dysregulated immune-escape signaling in the placentas of patients with GDM. The placenta expresses diverse receptors and secretes a wide variety of cytokines, chemokines, and adipokines, and plays an important role in maintaining pregnancy and the interaction of maternal and fetal units. Thus, studying the role and changes in the placenta is needed to identify the mechanism of GDM. A previous study based on microarray data in the placenta identified an increase in apoptosis-related genes and inflammatory signaling in GDM.30 In addition, single-cell transcriptomics analysis in the GDM placenta revealed the presence of diverse immune cell infiltration and altered NK cell status18; however, the role of T cell homeostasis in the placenta remains unclear. In our data, various immune related signaling, particularly T-cell immunity related pathways, such as regulation of lymphocyte activation, T cell receptor signaling, T cell differentiation, and positive regulation of immune response, were downregulated in placenta of GDM.
Whereas previous studies using the GDM placenta predicted signaling by measuring the genetic expression or concentration of cytokines representing the T cell subset, we sorted the placenta into the T cell subset using FACS analysis to demonstrate the T-cell immunity in placenta of GDM patients. From our results based on T cell subset, pro-inflammatory cytokines, especially IFN-γ and Th17, were increased in CD4+ T cells of the GDM placenta. While most of the studies about the inflammatory cytokine in GDM were observational studies related to increases in serum or amniotic fluid of inflammatory cytokines,231 our results suggested the possibility that the placenta is a source of inflammatory cytokines and the impact of inflammatory T cell subset associated with GDM. Moreover, the increase in memory CD4+, memory CD8+ T cells, pro-inflammatory T cells, IFN-γ+CD4+ T cells, INF gamma+CD8+ T cells, and IL-17A+CD4+ T cells observed in our study was consistent with a previous study that focused on the increase in the immune profile of Th2 and Th17 cells. Previous study revealed an increase in the pro-inflammatory T cell subset, the ratio of Th17/Treg, and association with postprandial glucose levels,29 supporting the link between dysregulation of adaptive immunity and hyperglycemia.
CD28-T cells are known to have shortened telomerase, reduced diversity, and cytotoxic capacity of T cell receptors, and CD57+ T cells are known to be senescent T cells with poor proliferation response to antigen-specific stimulation.3233 Increasing evidence suggests the failure of T cells is mainly due to the dysfunction of T cells and inducing T cell senescence is a key strategy in maintain immunoescape signaling.31 Senescent T cells, with defects in proliferation and effector functions, are known to be accumulated in aging, chronic viral infections, and autoimmune disorders where antigen stimulation persists.34 Previously, we identified the increased senescent T cells in type 2 diabetes patients and suggested the role of senescent T cells in metabolic dysregulation status.25 In the present study, we firstly identified an increase in the number of senescent T cells, CD28-CD57+CD4+ T cells, and CD28−CD57+CD8+ T cells using FACS in GDM placenta, the consistent findings with the RNA-seq. Collectively, our results suggested the contributions of T cell senescence in placenta related with GDM, although functional studies are needed to determine whether this is a cause or consequence of hyperglycemia or insulin resistance.
Maternal-fetal immunotolerance represents a real immunological challenge for the immune system of the mother. It is well known that immune checkpoints play a role in maintaining pregnancy and complications.35 The expression of PD-1, PD-L1, PD-L2, and CTLA-4 at the feto-maternal interface is detected in the decidua and placenta, and miscarriage occurs when monoclonal blocking antibodies of PD-L1 or PD-L2 are administered.36 Disrupted homeostasis PD-1/PD-L1 signaling has also been observed in pregnancy-related perinatal complications, implantation failure, recurrent spontaneous abortions, and preeclampsia.3738 Diverse reported evidence, consistent increased serum soluble PD-L1 along with increasing gestational age,39 PD-L1 expression in syncytotrophoblasts, extravillous cytotrophoblasts,40 and the exosome of placenta,41 supporting the role of PD-L1 in the maintenance of pregnancy. The importance of PD-1+ in Tregs was observed, such as a study showing a significant decrease in CD279+(PD-1+) CD4+CD25+ in PBMCs and decidua compared to controls in recurrent miscarriage mothers.42 A PBMC study reported that PD-1 expression on T cell subsets increased during normal pregnancy, but this increase was not observed at the time of diagnosis as GDM, and PD-1 expression was restored to normal levels with good glucose control.43 In another study, the assessment of immune checkpoint expression in the peripheral blood T subpopulation showed lower frequencies of CD3+CD4+CTLA-4+ and higher frequency of CD4+PD-1+ T cells in patients with GDM compared to healthy pregnant women, indicating dysregulated maternal-fetal tolerance associated with GDM.17 To investigate the role of T-cell immunity in the placenta, we investigated the characteristics of T-cell subsets, exhausted T cells and PD-1 expression in GDM placentas using FACS. We also examined PD-1/PD-L1 expression and related signaling through IHC and western blot analyses. Notably, based on the GSE results, this pattern was different from that of early pregnancy. Therefore, our results suggest that the increase in the expression of the immune checkpoint molecule, PD-1 or PD-L1, at the end of pregnancy may be a compensatory reaction to various inflammations rather than a proper immunosuppressive role in the maternal-placental interaction.
In GDM, the placenta serves as a target organ for increased insulin-like growth factor 1 (IGF-1) and hyperinsulinemia, and it is known to transport various amino acids and glucose to the fetus.44 Additionally, upregulation of IGF-1 signaling and diverse growth factors are associated with the development of macrosomia in patients with GDM.45 GDM placenta has been reported to have more increased glycogen deposits, increased volume including villous edema, increased angiogenesis, and deposition of lipid droplets compared to normal individuals.46 The best-known mechanism of these changes is persistent hyperglycemia and upregulation of insulin signaling caused by hyperinsulinemia due to insulin resistance.47 We observed increased STAT1, STAT3, and PI3K signaling, known as ones of insulin- or T cell fate-related signaling, as in the previous literature on GDM placenta by western blot analysis. Additionally, we also observed the upregulation of cell ATP synthesis or oxidative phosphorylation in GDM compared to in controls, although the further study is necessary to identify the precise mechanism.
This study has several limitations. First, the mechanisms or interactions could not be elucidated by transcriptomic analysis and expression measurement using whole tissue because the placenta is composed of diverse and heterogeneous cells. Second, the sample size was small, and validation of protein expression and related signaling was not enough. It remains unclear whether the change in the placenta may be caused by GDM or contribute to GDM or complications. Further molecular mechanistic studies should be performed for a more comprehensive understanding of exhausted T cells and PD-1/PD-L1 signaling in GDM. However, this is the first study that evaluated senescent T cells and immune-escape signaling in the placenta of patients with GDM using FACS analysis.
In conclusion, this study demonstrated that altered pro-inflammatory T cells, senescent T cells, and immune checkpoint molecules are present in GDM placentas. Our data suggest that dysregulation of immune-escape signaling and senescent T cells in the maternal placenta could be related to the pathophysiology of GDM.
 XML Download
XML Download