

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Apart from well-documented calcium channels, increasing evidence demonstrated that store-operated calcium entry (SOCE) may also have a critical implication for maintaining synaptic functions in psychiatric disorders [1]. The endoplasmic reticulum (ER) acts as intracellular calcium storage. The ER membrane-expressed Ca2+ sensor stromal-interacting molecule-1 (STIM1) maintains calcium homeostasis and innate inflammatory response through coupling with cell membrane transient receptor potential channels (TRPCs) or Orai1 to mediate calcium refilling in physiological conditions [2]. Cytosolic calcium, as the second messenger to regulate various functions, shows increased concentrations in the cytoplasm following lipopolysaccharide treatment in microglia [3]. Intracellular calcium activates calcineurin and immune transcription factor nuclear factor of activated T cells in acute brain insults, SOCE deficits might trigger Ca2+ overload and synaptic remolding in dopaminergic neurons in relation to ER stress [4]; it also acts as the dominant factor in the pathogenesis of Alzheimer’s disease (AD) [5]. Therefore, the alteration of SOCE functions is a cause or a critical target for mental disorders.
SOCE has a crucial role in modulating neuronal plasticity such as stroke, hypoxic/ischemic damage, and AD [6,7]. SOCE inhibitor CM-EX-137 completely abolishes inflammation-induced nitric oxide release, reduces calcium accumulation, and improves neurological deficits in a rodent model of experimental brain trauma [8]. SKF96365 is a prominent candidate in neuropsychiatric diseases; it significantly enhances N-methyl-D-aspartic acid receptor (NMDA)-mediated calcium influx and can be a novel therapeutic and diagnostic drug in calcium-dependent neuronal process [9]. Recent studies have identified that SKF96365 reverses leptin-induced carotid sinus nerve hypoxia and contributes to neural glucose metabolism via mediating SOCE activity [10]. SKF96365 was able to suppress high glucose-induced neuronal apoptosis in primary hippocampal neurons and alleviated isoflurane-induced neurotoxicity in high glucose treated SH-SY5Y cells [11]. By contrast, the functional role of SOCE in mediating neuropathic pain is largely unknown.
Using siRNA to specifically knockdown STIM1 and Orai1 can completely rescue the expressions levels of tumor necrosis factor-α and interleukin-6, which are identified as the dominant factors at the early stage of neuropathic pain [12]. Activation of SOCE in dorsal root ganglion (DRG) primary neurons remarkably enhances membrane depolarization and neuronal intrinsic excitability, which are characterized by pain pathology and pain messages transduction [13]. In addition, activation of SOCE may significantly promote extracellular calcium influx and strengthen synaptic transmission and neuronal resting membrane potential, it has demonstrated that knockdown of STIM1, or Orai1 exhibit a distinct connection of neuronal activity in formalin-induced spontaneous nocifensive behaviors [14]. A previous study demonstrated that SOCE mediated calcium entry in DRG is associated with inflammatory neuropathic and diabetic pain [13,15]. Therefore, SOCE is involved in inflammatory and neuronal activity at synaptic levels, which is the typical characterization in neuropathic pain.
Thus, abnormal function of SOCE may be a cause or leading factor for pain development. In our study, intrathecal SKF96365 produced remarkable analgesic effects in neuropathic rats. As a result of its complicated cell types in the spinal cord, immunofluorescence staining indicated that SOCE was distinctly expressed in neurons. We also found that the underlying plasticity mechanism of SKF96365 in neuropathic pain was mediated through inhibition of spinal glutamatergic transmission and mitigation of ER stress. Consequently, our studies investigated that SOCE antagonist, SKF96365, could potently suppress the activation of spinal SOCE, relieve ER stress, and suppress hypersensitivity after nerve ligation-induced neuropathic pain in rats.
METHODS
Chemicals and reagents
Picrotoxin (PTX) and SKF96365 were purchased from Sigma-Aldrich (St. Louis, MO, USA). Tetrodotoxin (TTX) and strychnine were obtained from Aladdin (Shanghai, China) and Toronto Research Chemicals (North York, ON, Canada), respectively. Anti-STIM1 antibody was from Novus Biologicals (St. Charles, MO, USA).
Animals
Wistar rats were obtained from SLAC Lab (Shanghai, China). All experiments followed the animal care guidelines of the National Institutes of Health and were approved by the Animal Care and Welfare Committee of Shanghai Jiao Tong University. They were raised in a controlled humidity environment and controlled temperature (23°C ± 1°C) on a 12 h light/dark cycle with lights on at 7:00 AM. Food and water were provided ad libitum.
Intrathecal catheterization and injection in rats
After being accustomed to the circumstances for 7 days, Wistar rats were anesthetized by intraperitoneal injection of pentobarbital sodium (50 mg/kg). A catheter with 0.28 mm inner diameter and 0.61 mm outer diameter (PE-10; Clay Adams, Parsippany, NJ, USA) was inserted into the lumbar spine of the rats and fastened by two joints near the ilium and neck in case of displacement. After 10 μl of 4% lidocaine in artificial cerebrospinal fluid (ACSF: 125 mM NaCl, 1 mM MgCl2, 3 mM KCl, 26 mM NaHCO3, 2 mM CaCl2, 1.25 mM NaH2PO4, and 10 mM D-glucose, pH 7.3) was administered and flushed with 15 μl of ACSF, typical bilateral paralysis was observed in well-operated rats.
Spinal nerve ligation (SNL) and behavioral testing
According to the well-documented literature [16], SNL model were generated by ligations of left spinal nerves L5 and L6 using a man-made glass rode and tightly ligated with 6–0 silk sutures in turns. The wound after those procedures was sutured before using broad antibiotics to prevent any infections. Before the behavioral tests, the rats were allowed to recover without locomotion deficits, and they eventually developed hypersensitivity with paw withdrawal thresholds < 8 g. The sham rats were operated identically without any ligations of L5 and L6.
For mechanical threshold testing, the rats were well-handled by investigators and randomly divided into groups who were blind to the investigators. The rats were gently transferred into the plastic box and acclimated for 30 min before the tests. Drugs were intrathecally injected in 10 μl of ACSF, followed by 15 μl ACSF. The mechanical threshold, which was stimulated by three times of upward force at 1 min intervals, could induce withdrawal, lifting, and licking in rats. The mechanical threshold was measured before, and 30 min, 1 h, 2 h, and 4 h after injections.
Spinal slice preparation
Under inhaled isoflurane anaesthesia, L3-L5 of the spinal cord was gently separated and quickly immersed into ice-cold high sucrose ACSF (234 sucrose, 3.6 mM KCl, 25 mM NaHCO3, 1.2 mM NaH2PO4, 12 mM glucose, 2.5 mM CaCl2, and 1.2 mM MgCl2). Using a vibratome (Leica VT-1200S; Leica Microsystems, Wetzlar, Germany), lumber enlargements were sliced at a speed of 0.03 mm/s and 400-μm width against agarose. The slices of the spinal cord were transferred into 32°C ACSF for 30 min and cooled to room temperature for 30 min before recording.
Whole-cell patch-clamp recordings
Neurons in spinal lamina II were attached with a pipette from a horizontal puller (P-97, Sutter Instruments) based on previous studies [17]. Before the recording, the pipette filled with K-based interval solution (135 mM K-gluconate, 5 mM EGTA, 5 mM HEPES, 2 mM MgCl2, 5 mM Mg-ATP, and 0.5 mM CaCl2, pH7.3) was applied to record the mEPSCs, and bath application of 0.5 μM TTX, 100 μM PTX, and 1 μM strychnine was used to inhibit inhibitory neuronal transmission and sodium channel activity. mIPSCs were recorded under high-Cl- interval solution (140 mM CsCl, 1.1 mM EGTA, 10 mM HEPES, 2 mM MgCl2, 3 mM Mg-ATP and 0.3 mM Tris–guanosine triphosphate, pH7.4 adjusted with CsOH) and clamped at −70 mV after bath application of TTX (0.5 μM), CNQX (20 μM) and D-AP5 (50 μM). Neurons were clamped at –70 mV for 5 min for analysis of the parameter of access resistance ≤ 30 MΩ. Data were filtered at 2 kHz and digitized at 5 kHz using pClamp10 software and then acquired in an Axonpatch 200 B amplifier (Axon Instruments, Foster City, CA, USA).
Immunofluorescence staining
Rats were perfused with 30 ml of ice-cold 0.9% NaCl solution followed by 50 ml of 4% paraformaldehyde (w/v) in pH 7.3 phosphate buffer saline (PBS) after anesthetizing via intraperitoneal injection of pentobarbital sodium (50 mg/kg). The lumber enlargements were postfixed for 2 h at 4°C in 4 % paraformaldehyde and dehydrated in 30% sucrose twice. Spinal slices were cut into 30 μm slices in optimal cutting temperature compound embedding compound and immersed in PBS. Spinal slices blocked with 10% bovine serum albumin (v/v) and 0.3% Triton X-100 (v/v) for 1 h at room temperature (23°C ± 1°C) were further incubated with primary antibodies including goat anti-iba-1 antibody (1:300; Abcam, Cambridge, MA, USA), chicken anti-GFAP antibody (1:300; Millipore, Billerica, MA, USA), mouse anti-NeuN antibody (1:300; Millipore), and rabbit anti-STIM1 (1:300; Novus) for 24 h at 40 rpm.
RNA isolation and quantitative reverse transcription PCR (qRT-PCR)
Enlarged L3-L5 of the ipsilateral spine from the sham group and SNL group were gently separated into cold ACSF. TRIzol was used to extract total RNA, and the ratio of 260/280 > 1.8 was selected for further experiments. About 1 μg of mRNA was used for cDNA synthesis using a first-strand cDNA synthesis kit (Thermal, K1622). The primers used were as follows:
CHOP Forward: TCCCCAGGAAACGAAGAGGAAGAAT;
CHOP Reverse: CCCTGGCTCCTCTGTCAGCCA;
HSP70 Forward: TGTCTTCTCAGCATCAAGCAAGG;
HSP70 Reverse: CCAACACTTCCTGGACAGGCTT;
GAPDH Forward: CATCACTGCCACCCAGAAGACTG;
GAPDH Reverse: ATGCCAGTGAGCTTCCCGTTCAG.
Real-time quantitative PCR experiments were performed using Takara qPCR Mix. The relative expression of CHOP and HSP70 was calculated using the 2-∆∆CT method after normalization to GAPDH.
Data evaluation and statistics
The percentage of the maximal possible effect (% MPE) was calculated by using the following formula: (post-drug threshold in ipsilateral hindlimb – baseline threshold in ipsilateral hindlimb) / (baseline threshold in sham hindlimb – baseline threshold in ipsilateral hindlimb) × 100. The % MPE values near to 100 were indicated as mechanical thresholds (i.e., near sham group thresholds), while values closed to 0 indicate nerve injury-induced mechanical allodynia. For analysis of the dose-response curves, the parameters, i.e., the minimum effect, maximum effect (Emax), half-effective dose (EC50), and Hill coefficient (n), were statistical calculated by fitting nonlinear least squares curves to the relation Y = a + bx, where x = [C]n / (EC50n + [C]n) using GraphPad Prism (Version 7.0; GraphPad Software Inc., San Diego, CA, USA). The values of EC50 and b (Emax) were statistical calculated by yielding a minimum residual sum of squares of deviations from the theoretical curve.
Data were performed for Gaussian distribution assessment by Shapiro–Wilk normality test after transformation. Data are shown as the means ± standard error of the mean (SEM). The variance was analyzed using unpaired, two-tailed Student’s t-test, and one-way or repeated measures two-way ANOVA by GraphPad Prism followed by Sidak’s post-tests. Statistical significance was set as p < 0.05 in our study. Data were presented using GraphPad Prism 7.0 (GraphPad Software Inc.) and CorelDraw 2019.
RESULTS
Intrathecal SKF96365 exerted mechanical antiallodynia in neuropathic pain via inhibition of SOCE
To investigate the analgesic effects of the SOCE antagonist SKF96365, we tested behavioral thresholds using non-invasive mechanical tests in both sham and neuropathic rats induced by ligation of spinal L5 and L6. Four groups of neuropathic rats were intrathecally injected with SKF96365 (saline: ACSF 10 μl, SKF96365: 10, 30, or 100 μg). The operated hindpaw withdrawal thresholds to von Frey monofilament stimulus were measured before and at 1, 2, and 4 h after injections. In sham rats, intrathecal injection of 3, 10, 30, and 100 μg of SKF96365 and ACSF did not change contralateral hindpaws during the 4 h observations. By contrast, intrathecal delivery of SKF96365 prolonged ipsilateral hindpaws in a dose- and time-dependent manner (Fig. 1A, mechanical allodynia F(9,160) = 16.22, p < 0.0001; two-way repeated-measures ANOVA followed by Sidak’s post-tests). Based on previous study [18], the projected Emax and ED50 values for mechanical anti-allodynia inhibition were 82.5% MPE and 18 μg, respectively (Fig. 1B).
STIM1 is widely expressed in excitatory cells of the spinal cord
SOCE mainly encompassed STIM1, which could bind with cell membrane Orai1 and TRPCs. Using immunofluorescence staining in the spinal cord, STIM1 was treated as a specific biomarker of SOCE. DAPI was applied to mark each cell. To further characterize the neuronal cells that expressed STIM1, we carried out immunofluorescence staining to locate the co-expression of STIM1 with differential cell markers of the spinal cord including NeuN, GFAP, and Iba1. We found that STIM1 co-labeled with NeuN but not with astrocytes or microglia (Fig. 2). Our data demonstrated that STIM1 was specifically distributed in neurons without expressing on astrocyte or microglia.
Inhibition of STIM1 by SKF96365 ameliorates nerve injury-induced glutamatergic transmission in substantia gelatinosa
Recent studies demonstrated that imbalance in spinal substantia gelatinosa can synchronously lead to spinal neuronal transmitter release, pathological neurotransmitter release, and pain sensation arousal. Herein, we assessed the electrophysiological mechanism of SKF96365 at neuronal transmission levels. Both frequency and amplitude of mEPSCs were increased in the spinal cord in neuropathic rats. We observed that bath application of SKF96365 (3, 10, and 30 μM) could significantly suppress the increased frequency and amplitude of mEPSCs in a dose-dependent manner. Meanwhile, 30 μM SKF96365 did not show any inhibition on neurons from sham rats (Fig. 3A–C Frequency, F(5,64) = 16.56, p < 0.0001; Amplitude, F(5,64) = 15.37, p < 0.0001, one‐way ANOVA followed by Sidak's post‐tests).
Inhibitory neurotransmission is another cause of pain pathology that results in signaling transmission of magnified pain. The frequency of mIPSCs was significantly suppressed in neuropathic rats compared with that in sham rats. Bath application of 30 μM SKF96365 did not show any inhibition on the frequency and amplitude of mIPSCs in both neuropathic and sham rats (Fig. 3D–F Frequency, F(3,42) = 5.621, p = 0.0025; Amplitude, F(3,42) =0.05198, p = 0.9841, one‐way ANOVA followed by Sidak’s post‐tests). These data indicated that SKF96365 restored pathological neurotransmission in the spinal cord via alleviating glutamatergic transmission in neuropathic pain.
SKF96365 ameliorates nerve injury-induced neuronal ER stress in neuropathic rats
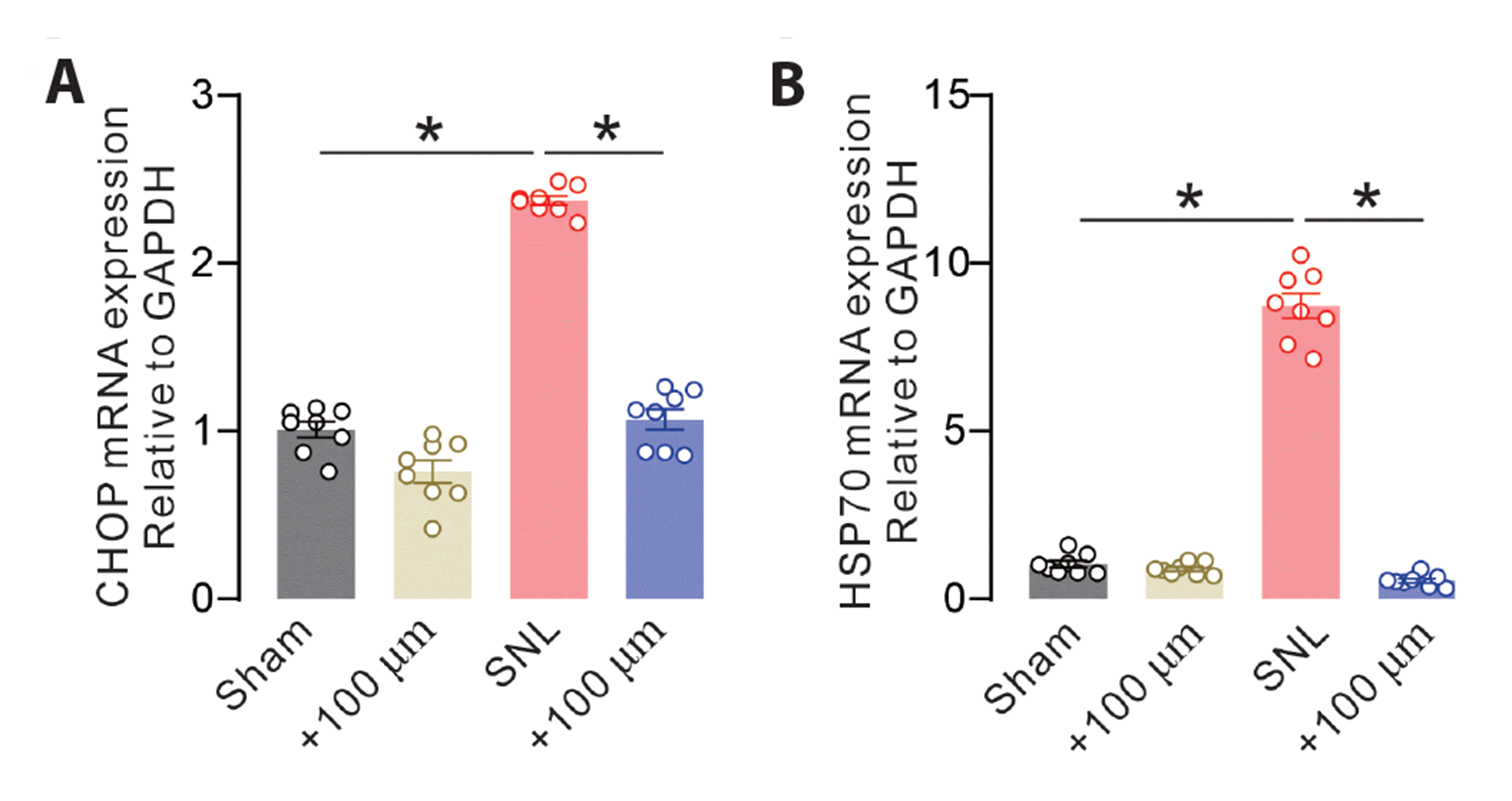
Given STIM1 is specifically expressed in the neuronal ER membrane, its maladaptive expression may drive pathogenesis in pain [19,20]. A previous study demonstrated that inhibition of spinal ER stress may have remarkable potential for pain relief [21]. Thus, we further explored the neuronal mechanism of SKF96365. Neuropathic rats exhibited strong CHOP and HSP70 expression, and SKF96365 significantly suppressed abnormal mRNA expression of CHOP (Fig. 4A, F(3,28) = 187.5, p < 0.0001) and HSP70 (Fig. 4B, F(3,28) = 404.5, p < 0.0001) in the spinal dorsal horn. These data indicated that SKF96365 ameliorated nerve injury-induced neuronal ER stress via inhibition of STIM1 functions in neuropathic rats.
DISCUSSION
In a population-based case–control study, patients with pain sensations have been reported to have a greater prevalence of mental disorders and somatization than that found in the general population [22]. The clinical epidemiology investigation demonstrates that pain patients are more likely developed to anxiety, depression, social deficits, and insomnia [23]. Therefore, pain, characterized by abnormal neuronal circuits in the spinal cord and brain, is a complicated mental disorder, and eventually leads to the high morbidity of diseases [24]. Emerging evidence demonstrated that imbalance of spinally glutamatergic and GABAergic transmission contributes to brain synaptic transmission and behavioral changes in neuropathic pain, accompanied with increased activity of hyperactivation including voltage-dependent calcium channel family, transient receptor potential vanilloid channels, and calcium-activated potassium channels [25-27]. Synaptic calcium channels can initiate and drive neuronal depolarization; these alterations can further trigger calcium influx into neurons and neuronal transduction, especially at the early stage of pain [28]. In addition, inflammatory mediators or pro-inflammatory factors can remarkably sensitize those calcium channels and contribute to maladaptive neuronal plasticity in pain pathology [29].
A previous study demonstrated that therapeutical manipulation of spinal neuronal transmission including inhibition of glutamatergic transmission, is a predominant factor for pain management [30]. Calcium depletion in ER might drive the formation of ER tethering protein and UPR-independent PERK dimerization, which are commonly associated with human platelet aggregation and AD [31-33]. In accordance with pain pathogenesis, increased calcium in the presynaptic zone mediated by a variety of calcium channels have linked to mediate inflammation, neuronal plasticity, and pain states, which might present a promising strategy in pain management [34,35]. Endogenous SOCE is the dominant calcium channel by mediating calcium influx upon calcium deletion as well as neuronal excitability and neuronal plasticity. In addition to being involved in glial inflammation, SOCE may be implicated in neuronal excitability, which is accompanied with numerous protein expressions and behavioral abnormalities. Synaptotagmin-7 (Syt-7) was the typical calcium sensor in mediating calcium homeostasis in mental psychiatry. It was a both leaky and asynchronous calcium sensor, involved in neuronal membrane trafficking in processes of neurodegeneration [36]. SNARE-complex and the Syt isoform 7 results in a relatively high energy barrier and the rapid release of neurotransmitter vesicles evoked by various inputs [37]. In addition, Syt isoform 7 deficits can trigger multiple forms of glutamate release and NMDAR-mediated bipolar-like behavioral in rodent [38]. Therefore, calcium homeostasis is crucial for maintaining neuronal plasticity and behavioral phenotypes.
Current opinion from electrophysiological data indicated that restoration of glutamatergic transmission in the spinal cord, a key region that mediates pain signal transmit, can remarkably alleviate hypersensitivity following nerve injury. Exposure to SKF96365, serving as a pharmacological agent, produced anti-allodynic effects in neuropathic rats and exerts long-lasting alterations in synaptic transmission. Recent studies illustrated that glutamatergic transmission contributes to allodynia, reconstruction of inhibitory transmission, and SOCE expression. Inhibition of spinal glutamatergic transmission in rats resulted in remarkable hyperpolarization and downregulation of glutamate release along with analgesic effects in neuropathic pain following SKF96365 treatment. Neurons in substantia gelatinosa received glutamatergic transmission from DRG and inhibitory transmission inputs from periaqueductal gray, rostral ventromedial medulla, and dorsolateral funiculus, which were the major components in descending inhibitory system [31]. Unlike synaptic transmission in brain, both glutamatergic and inhibitory synaptic transmission are separated in anatomic structure. In addition, previous study have thoroughly demonstrated that inhibition of glutamatergic transmission was sufficient to abolish hypersensitivity in neuropathic pain [39].
Previous study demonstrated that inflammatory mediators may chronically potentiate intracellular calcium level and microglia-mediated inflammatory-like response in psychiatric disease [40]. The neurosteroid pregnenolone sulfate exerted a broad impact on the excitability of medullary dorsal horn neurons and alleviated nociceptive behaviors by decreased calcium concentration within glycinergic nerve terminals in neuropathic model [41]. Therefore, many study inferred that manipulation of calcium concentration might produce analgesic effects. SOCE is the major channel in modulating intracellular calcium refilling. YM-58483, a potent SOC channel inhibitor, dramatically attenuated inflammatory mediators-induced overactivation in SNI and complete Freund adjuvant-induced hypersensitivity [42]. TRPC6 subunits mediated capacity was remarkably up-regulated in spinal cord in morphine analgesic tolerance model, its inhibitor could be successfully utilized for the prevention of chronic pain [43]. Therefore, manipulation of calcium channel capacity in presynaptic may produce analgesic effects in pain sensations.
Electrophysiological data found that inhibitory neurotransmission in spinal lamina II neurons is significantly diminished, and deficits in inhibitory transmission can lead to hypersensitivity, exerting long-lasting message transformation to the central neural system. Interestingly, targeting glutamatergic events might partially alleviate neuronal imbalance and prolonged paw withdrawal latency to control levels in the ipsilateral paw [44,45]. Therefore, our study uncovered that inhibition of glutamatergic transmission is sufficient for pain management in neuropathic pain, consistent with the findings of a previous study [46].
In this study, we explored the pathological and physiological roles of SOCE in pain states application of SKF96365. It produced distinct effects on thapsigargin(tg)-induced calcium refilling. In particular, SOCE contributed to spinal-mediated calcium homeostasis in neurons and sensory process [47]. Inflammatory mediators including ATP and bradykinin in pain underlined a strong and long-lasting intracellular calcium influx, indicating a significant and robust nocifensive behavior in rats associated with the upregulation of SOCE [48,49]. Meanwhile, many studies have demonstrated that naïve SOCE can potentiate spinal-mediated excitatory action and maintain signal transmission in primary neurons [14]. Previous studies demonstrated a strong positive correlation between inhibition of SOCE and nocifensive behavior [48]. Using the inhibitor of SOCE, we identified that the inhibition of SOCE is involved in the pathological calcium changes, a process that mediates the alterations in glutamate levels. Therefore, we unveiled that novel SOCE may stimulate neuronal activity in maintaining normal sensory activity.
SOCE in astrocytes or microglia also exhibited neuroprotection by decreasing Aβ42 accumulation and clearance, and the imbalance of SOCE may lead to glial inflammation and the deposition of amyloid-beta (Aβ) [50]. Apart from the modulation of neuronal functions, astrocytic SOCE also necessary for glycogen metabolism and synaptic formations [13,51]. Related studies suggested that calcium channel, including Orai1 and STIM1, underline a crucial entry of calcium influx in astrocytes [52]. Long-term exposure to 2-[2-[4-(4-nitrobenzyloxy)phenyl]isothiourea caused remarkable calcium alterations in both neurons and astrocytes, as well as activation of SOCE [53]. Furthermore, application of pharmacological inhibitors of 1-(5-chloronaphthalene-1-sulphonyl)-1H-hexahydro-1,4-diazepine (ML-9) completely abolished neuronal calcium aggregations, and reduced synaptic exocytosis between connected neurons in primary cultured cells [54]. STIM1 is a key component in maintaining calcium refilling, double immunofluorescence revealed that STIM1 was totally colocalized with neuronal marker in the spinal cord. In addition, bath application of SKF96365 produced direct effects on the frequency and amplitude of mEPSCs. However, STIM1 is widely expressed in various brain regions, including the hippocampus, cortex, thalamus, and amygdala in the cognitive processes, as well as pain pathology characterized by an increased calcium level in presynaptic calcium [55,56]. Our results illustrated the possibility of the neuronal calcium channel contributing to neuronal transmitter release after nerve injury.
ER stress has been identified as the dominant characterization, which is associated with pathological SOCE related to pain [57]. ER stress inhibitor significantly reduces pain hypersensitivity behaviors depending on Prostaglandin E2 expression [20]. SOCE is crucial for ER response towards stress, and ER stress is remarkably altered in type 2 diabetes mellitus by modulating basal ER Ca2+ levels [58]. Nerve injury induced ER stress may be implicated in the impairments of neuropathic pain [59]. The actions of long-term exposure to ER stress caused significant neuronal death and synaptic plasticity deficits [60]. As a dominant characterization following ER Ca2+ alterations, maintaining ER stress provided a critical method for neuronal plasticity. Taken together, we uncovered that SKF96365 completely alleviated ER stress, which might exhibit anti-hypersensitivity.
Our results uncovered that SKF96365 produced long-lasting anti-hypersensitivity in neuropathic rats, and alleviated spinal glutamatergic neurotransmission through the inhibition of neuronal SOCE. Fig. 5 schematical illustrates the possible mechanism of SKF96365 on arrested calcium influx from the extracellular environment, restored neurotransmission, and behavioral hypersensitivity in neuropathic pain. These data support the concept that manipulation of SOCE might be an effective target in the strategy of pain management.



 XML Download
XML Download