

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Many lines of evidence, including epidemiologic studies from human beings and extensive animal experimental data, indicate that early life events play an essential role in influencing later behavioral and emotional responses to stressors [1-4]. Although the mechanism of how early life stress changes the behavioral and/or emotional outcome remains unclear, a recent explanation for this is that adverse early life experiences can change neural connectivity in the underlying brain network [5-9]. For example, abnormal maternal care and chronic early life stress change the number and function of neuronal synaptic activities in different brain areas, including the hypothalamus, hippocampus, septal area, etc. [10-13]. These lines of evidence suggest that maternal separation (MS) in early life may hinder the normal neural circuitry of the brain, resulting in emotional disorders such as anxiety and aggressive behavior.
The hippocampus, belonging to the limbic system, is a primitive cortical structure that plays an essential role in memory processing and spatial cognition, including how mammals learn to understand and navigate the environment. For this reason, several researchers have long focused on its role in spatial learning memory and dementia, especially concerning Alzheimer’s disease [14-18]. However, literature has also shown that the hippocampus takes part in memory and intimately in emotion to regulate a range of stress responses [19-21]. In humans, the decline in the volume of the hippocampus and its resulting dysfunction are related to psychological disorders, such as post-traumatic stress disorder, bipolar disorder, and depression [22-26]. These two distinct functional roles of the hippocampus can be explained by dividing it into two separate regions, namely, the dorsal hippocampus and the ventral hippocampus [10,27-29]. The dorsal hippocampus receives polymodal sensory inputs from the cortical areas. It is involved mainly in learning and memory, while the ventral hippocampus is more closely linked to subcortical limbic areas (hypothalamus, amygdala, etc.) and is mainly associated with the modulation of reward circuitry and emotional behavior [10,30-32].
Many lines of evidence have shown that hippocampal GABAergic interneurons are associated with mood disorders’ development [33-35]. The interneurons control the activity levels of principal neurons by gating the information flow at the synaptic cleft [10,36-38]. Although interneurons possess the largest diversity in morphology and physiological properties, Most interneurons in the central nervous system are inhibitory interneurons, which characteristically release gamma-aminobutyric acid as their neurotransmitter [39]. The hippocampus is a very susceptible structure to the effects of different stressors and is known to be involved in fear, anxiety, or aggression through direct projection reaching multiple areas of the brain, such as the hypothalamus, prefrontal cortex, basal amygdala, and nucleus accumbens [40-44]. Particularly, the hippocampus sends projections to the hypothalamic paraventricular nucleus (PVN) for involvement in the regulation of the hypothalamic-pituitary-adrenal (HPA) axis [35,45,46].
It was previously shown that early life stress, especially MS, can decrease long-term potentiation and release probability of Mossy fiber-CA3 synapses at the ventral hippocampus in neonatal mice, which may result in behavioral disorders in the adolescent period [47]. These electrophysiological and behavioral data hint at a correlation between developmental changes in the ventral hippocampus caused by early life stress and psychiatric illness. In this study, we aimed to determine whether adolescent behavioral disorders following early MS were caused by alterations in the cytoarchitecture of the ventral hippocampus.
METHODS
Animals
The adult male C57BL/6 and Balb/c mice (Samtako Inc., Osan, Korea) were used. The C57BL/6 mice were used for all experimental procedures, and the Balb/c mice were used only as an intruder in the RI test. All animals were housed in a standard cage with sterilized food and tap water available ad libitum and maintained on a 12-h:12-h light-dark cycle (light on from 7:00 AM) under constant room temperature (21°C ± 2°C). All experimental procedures were conducted in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals (NIH Publication No. 80-23, revised 1996) with the approval of the Institutional Animal Research Ethics Committee of Eulji University (approval No. EUIACUC14-13).
Maternal separation
The MS experiments were performed using the same procedures as in our previous study [47,48]. Briefly, the male and female mice were mated to produce litters of 8–12 pups. After birth, the pups were randomly divided into two groups: the MS group and the handling (HD) group. The pups in the MS group were separated from their dams for four hours (10:00–14:00) per day from the postnatal day (PND) 2–20. During the separation, the pups were placed in another isolated room with heated, clean sawdust (29°C ± 1°C) to avoid communicating with their dams through ultrasound vocalization and their dams. The pups in the HD group remained in their home cage with their dams and littermates during the four-hour separation, except briefly moving them to a clean cage and returning to their home cage. This HD procedure, never lasting for more than five minutes per litter, simulates the handling undergone by MS pups and serves as a control [49,50]. Only one proficient researcher conducted all the MS procedures to prevent potential experimental bias. Other researchers were blocked from accessing the cages.
Experimental procedures
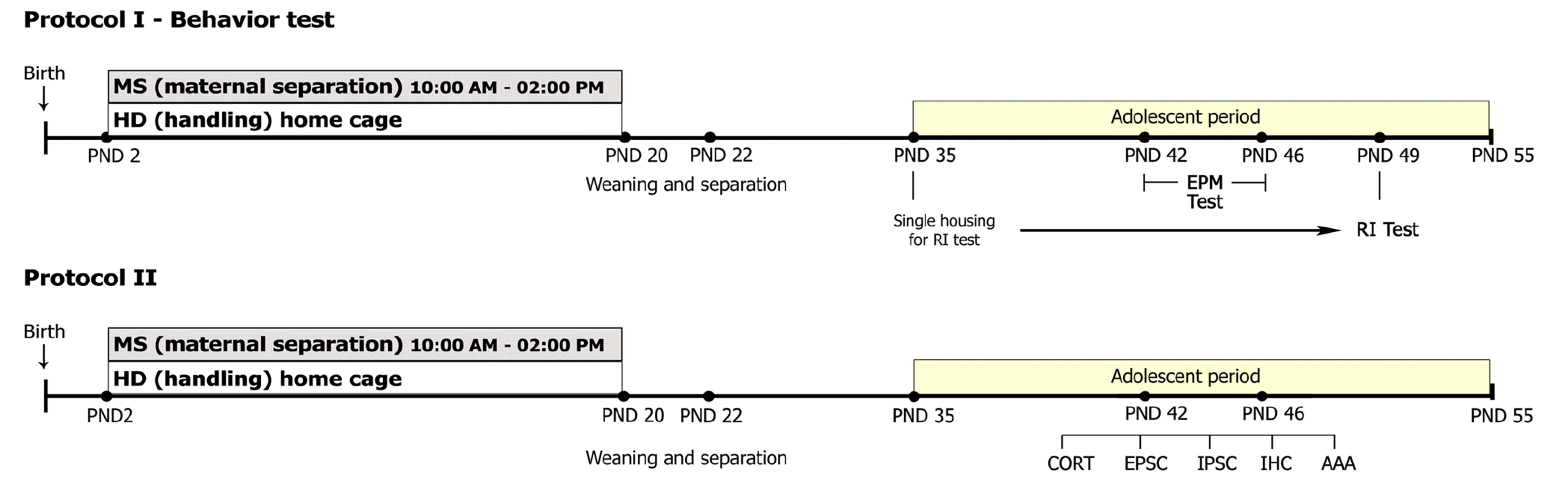
After weaning at PND 21, all pups were regrouped by gender at PND 22, and all subsequent procedures were conducted using males only. The behavioral changes of MS during the adolescence period were evaluated by accessing the elevated plus maze (EPM) test and RI test. EPM test was conducted between PND 42 and PND 46. RI test was conducted at PND 49 following single-housing of animals for two weeks from PND 35. Corticosterone (CORT) assay and immunohistochemistry (IHC) for GAD67 and parvalbumin (PV) experiments were conducted between PND 42 and 49 (Fig. 1). A total number of 68 MS and 68 HD was used in this study: EPM test (10 MS and 10 HD), RI test (10 MS and 10 HD), CORT assay (10 MS and 10 HD), IHC (13 MS and 13 HD).
Elevated plus maze test
The EPM apparatus is a plus-shaped acryl maze with two opposite open arms and two opposite closed arms, extending out from a central platform. The open arm is 65 × 5-cm, the closed arm is 65 × 5-cm, and the height is 15-cm. Animals were placed in the center platform of the maze facing one of the open arms and then allowed to explore the open or closed arms for ten minutes. The time spent in the different arms was recorded with the Ethovision Video Tracking System (Noldus Inc., Wageningen, Netherlands).
Resident-intruder test
For the RI test, from PND 35, each MS and HD mouse was single-housed in an individual home cage for 14 days. The bedding was not changed for four days (from PND 45 to PND 49) before testing. On the test day (at PND 49), the other mouse species (Balb/c) of the same age were placed into the home cage as an intruder. For ten minutes, aggressive behavior of the host mice were recorded using the Ethovision Video Tracking System. The latency, total duration, and the number of attacks on the host mouse were observed and assessed.
Blood collection and corticosterone assay
Anesthesia was induced using isoflurane (Isoflurane; Hana Pharm Co., Hwaseong, Korea) in an empty induction chamber. Following rapid decapitation, trunk blood was immediately obtained from animals, and the obtained blood (about 0.7-ml) was allowed to clot at room temperature for 30 min. After centrifugation (2,300 rpm, five min), plasma was transferred into fresh tubes and stored at –80°C until used for CORT analysis. Blood sampling occurred between 09:00 AM and 11:00 AM.
The serum levels of CORT were measured using Cayman’s Corticosterone EIA Kit according to the manufacturer’s instructions (Cayman chemical, Ann Arbor, MI, USA) as described previously by research groups [51-55]. Briefly, 100-μl standards or samples were added in duplicate to the wells of the microtiter plate. Assay buffer (200-μl) was added to the nonspecific binding (NSB) wells, and 100-μl assay buffer was added to wells to act as maximum binding wells. Then, 50-μl conjugate and 50 μl of CORT antibody (except the NSB wells) were added to each well and the plate was incubated for two hours at room temperature on a horizontal orbital microplate shaker (450–550-rpm). After the plate was washed thrice using the Washing Solution and the remaining Washing Solutions were completely removed by decantation and aspiration. Then, 200-μl p-nitro phenyl phosphate substrate was added to each well and incubated for one hour at room temperature without shaking. The optical density of the enzyme products was read at 405-nm using a plate reader (SPECTRO star Nano; BMG Labtech, Ortenberg, Germany) within 15 min after the reaction was terminated by adding 50-μl Stop Solution. The values of samples were calculated according to the standard curve (BMG Labtech).
Immunohistochemistry
Animals were anesthetized through an intraperitoneal injection mixture of ketamine (70-mg/kg) and rompun (8-mg/kg) and transcardially perfused with physiological saline, followed by 150-ml 4% paraformaldehyde in phosphate-buffered saline (PBS). Brains were eliminated immediately, post-fixed in the same fixative for two hours, and infiltrated with 30% sucrose solution overnight at 4°C until they sank. The whole brains were rapidly frozen in 2-methylbutane chilled on dry ice and mounted in Tissue-Tek OCT compound (Sakura Finetechnical Co., Tokyo, Japan). Serial horizontal sections of 40-μm-thicknesses were obtained on a Cryostat Microtome (Leica Microsystems Inc., Wetzlar, Germany). Every sixth section, at a periodicity of 240-μm, was obtained as one set, so six sets were prepared per brain. Before immunohistochemical staining, the sections of a set were scanned under a stereomicroscope (Nikon Inc., Tokyo, Japan) to select specimens, including a section passing through the ventral hippocampus between level Interaural 3.12–2.04-mm according to Paxinos and Watson [56]. A total of six specimens per brain were selected, and immunostaining was conducted.
IHC staining was executed using the free-floating method. The horizontal sections were washed for ten minutes in 0.1 M PBS, and endogenous peroxidase activity was quenched by incubating the tissue sections with 0.3% hydrogen peroxide in PBS for 30 min. After washing, the sections were incubated in PBS containing 0.1% Triton X-100 (PBST) and 10% normal horse serum for one hour to reduce nonspecific staining. The sections were then incubated with primary antibodies at 4°C overnight. The primary antibodies used in this study were mouse anti-GAD67 monoclonal antibody (1:1,000; Chemicon, Temecula, CA, USA) and mouse anti-PV monoclonal antibody (1:1,000; Sigma-Aldrich, St. Louis, MO, USA). The sections were washed thrice with PBST and incubated for two hours with biotinylated horse anti-mouse immunoglobulin G (1:200; Vector Laboratories Inc., Burlingame, CA, USA), followed by incubation for one hour with Avidin-Biotin Peroxidase Complex (Vector Laboratories Inc.). Antigens were visualized with 3, 3’-diaminobenzidine tetrahydrochloride (Sigma-Aldrich) solution containing 0.01% hydrogen peroxide. Sections were mounted on Vectabond (Vector Laboratories Inc.) coated slides, counterstained with hematoxylin, dehydrated through a graded ethanol series, cleared in xylene, and covered with coverslips using paramount (Fisher Scientific, Pittsburgh, PA, USA).
Cell count and semi-quantitative analysis
The number of GAD67- or PV-immunoreactive (-ir) neurons in the ventral hippocampus was counted using a researcher blind to the experimental conditions. Light photomicrography images were collected on an Olympus Ax 70 microscope (Olympus Inc., Tokyo, Japan) equipped with a Carl Zeiss AxioCam MRC digital camera (Carl Zeiss Inc., Jena, Germany) and AxioVision image capture software (version 4.6; Carl Zeiss Inc.). The researcher traced contours within the boundaries of each ventral hippocampal subfield (dentate gyrus [DG], CA3, and CA1) at 4× magnification. The border between each region was defined by Cappaert’s method [57]. Its coiled structure can distinguish the DG with an opened concave part directed towards the hippocampus proprius. The hippocampus proprius is subdivided into two subfields, CA1 and CA3, according to the density, size, and branching of axons and dendrites of pyramidal cells. CA3 is the region with large, less densely packed cells, and CA1 is the region with densely packed medium-sized cells. In this study, we did not include the field CA2, a narrow transitional field between CA3 and CA1.
Cells of GAD67-ir or PV-ir were then counted at 10× magnification using a meander scan profile counting technique. The meander scan profile counting method is a semi-automated scanning method that allows the user to view and count all cells within the traced contour. For a more precise comparison, we further divided each hippocampal subfield into several layers [57]. The DG was further divided into three layers: granular cell layer (gcl), hilus, and molecular cell layer (mcl). CA3 was further divided into five layers: stratum oriens (so), stratum pyramidale (sp), stratum lucidum (sl), stratum radiatum (sr), and stratum moleculare (slm). The CA1 was also further divided into four layers: so, sp, sr, and slm. To determine the density of immunoreactive cells in a given region of interest, we divided the total number of immunoreactive cells in that region by the total size of the traced contour (in mm2). Our cell counts are expressed as the average number of cells per mm2.
RESULTS
Neonatal MS induces elevated basal CORT and anxiety/aggressive-like behavior
To examine the effect of neonatal MS on behavioral characteristics in the adolescent period, two groups were assessed using the EPM test and the RI test. In the EPM test, the MS mice spent significantly less time in the open arms (HD: 65.28 ± 11.06 sec; MS: 35.29 ± 7.49 sec, p = 0.035; Fig. 2A) and spent more time in the closed arms (HD: 466.95 ± 14.02 sec; MS: 515.85 ± 15.75 sec, p = 0.030; Fig. 2B) compared to the HD group. Simultaneously, the ratio of entries into the closed arm section in the MS group was more than that of the HD mice (HD: 68.04 ± 1.73; MS: 78.24 ± 3.956, p = 0.034; Fig. 2D). These behavioral features show more anxiety-like properties in the MS mice. No significant differences were observed in the duration time in the center between the two groups (HD: 40.7 ± 3.23 sec; MS: 33.11 ± 6.13 sec, p = 0.297. data now shown). Moreover, no differences were shown in the total number of entries into all arms (HD: 27.3 ± 2.14 sec; MS: 20.77 ± 2.68 sec, p = 0.071. data now shown). These results indicate that the behavioral effects are probably unrelated to changes in activity levels.
In the RI test, the MS mice exhibited a short latency to attack intruder mice (latency: HD: 448.20 ± 96.90 sec; MS: 323.20 ± 65.62 sec, p = 0.028; Fig. 2E). Meanwhile, the MS mice exhibited significant elevated time in the total duration of attacks (HD: 8.12 ± 10.22 sec; MS: 42.23 ± 15.32 sec, p = 0.007; Fig. 2F) and increased the number of attacks (HD: 4 ± 2.84 sec; MS: 23.67 ± 5.38 sec, p = 0.001; Fig. 2G) compared with the HD mice. These results show that MS mice have more aggressive behavior than HD mice.
Based on the observed behavioral phenotypes in the MS group, basal CORT was examined to determine whether neonatal MS leads to an alternation of it. Fig. 2H showed elevated baseline CORT levels in MS mice compared to HD mice (HD: 66.14 ± 6.77-mg/ml; MS: 88.00 ± 8.89 mg/ml, p = 0.031).
Increased number of GAD67-ir neurons in the adolescent MS mice
The numerical densities of GAD67-ir neurons in vHipp were estimated, and their differences between MS and HD mice were statistically assessed (Figs. 3 and 4). The total number of GAD67-ir neurons per unit area (mm2) in DG, CA3, and CA1 (Fig. 3), subiculum, presubiculum, parasubiculum (Fig. 4) was significantly higher in MS mice than in HD mice (DG: HD: 222.92 ± 6.54; MS: 333.17 ± 10.46, p < 0.001; CA3: HD: 268.95 ± 3.24; MS: 387.62 ± 3.60, p < 0.001; CA1: HD: 189.53 ± 3.29; MS: 261.86 ± 6.61, p < 0.001; subiculum: HD: 156.39 ± 5.15; MS: 211.45 ± 6.91, p < 0.001; presubiculum: HD: 348.78 ± 9.185; MS: 589.95 ± 13.048, p < 0.001; parasubiculum: HD: 124.22 ± 6.57; MS: 184.50 ± 8.07, p < 0.001). The values for GAD67-ir neurons of each sublayer are presented in Figs. 3 and 4.
Increased number of PV-ir neurons in the adolescent MS mice
PV IHC staining of the ventral hippocampus indicated that the number of immune-reactivity cells per hippocampal area was significantly higher in the MS than the HD designed (Figs. 5 and 6). Similar results to GAD67, in the most regions the total number of PV-ir neurons in the MS group showed significantly higher than the HD group (DG: HD: 222.92 ± 1.12; MS: 333.17 ± 2.37, p < 0.001; CA3: HD: 90.48 ± 1.74; MS: 160.76 ± 3.59, p < 0.001; CA1: HD: 87.15 ± 3.09; MS: 109.91 ± 3.93, p < 0.001; presubiculum: HD: 282.43 ± 13.75; MS: 452.79 ± 18.97, p < 0.001; parasubiculum: HD: 42.45 ± 3.99; MS: 132.37 ± 13.22, p < 0.001). However, in the subiculum region (Fig. 6), there were no significant differences between the two groups (CA1: subiculum: HD: 72.27 ± 6.03; MS: 86.29 ± 6.87, p = 0.133).
DISCUSSION
In this study, the mice with neonatal MS exhibited increased anxiety-like/aggressive behavior in adolescence. This behavioral change was accompanied by an increase in the CORT level, suggesting that MS-induced anxiety and aggressive behavior may be because of changes in the HPA axis. Additionally, GAD67-positive cells increased in most hippocampus regions, such as DG, CA3, CA1, and subiculum, and parvalbumin-positive cells increased in DG and CA3.
There are five types of rodent models have been used to investigate the mechanisms linking early life stress to behavioral outcomes (mood disorders) in adulthood; 1) prenatal stress exposure, 2) acute maternal deprivation procedures, 3) chronic or periodic MS model, 4) chronic early life stress, 5) early weaning of the pups [58]. Among them, the most widely used manipulation to manufacture a rodent model of early life stress is MS. When tested in adults, rodents subjected to MS show enhanced anxiety and/or depressive behavior, deficits in learning and memory, enhanced expression of corticotropin-releasing factor, and alterations of mossy fiber density in the CA3 region of the hippocampus [59]. Because the adolescent period is a vital period for physical and mental development, the behavioral, hormonal, and neuro-architectural changes caused by MS could be facilitated and entrenched in this period. Although this is important, only a few studies have directly examined MS effect on adolescents [60]. This study aimed to examine the impact of MS on the behavioral, hormonal, and neuro-architectural changes in adolescent C57BL/6 mice.
Since rodents have innate fear of open spaces [61], the EPM is considered a gold-standard behavioral assay for anxiety-like behavior. Meanwhile, the RI test is widely used to evaluate aggressive tendencies because many species display territorial aggression against intruders in defense of mates and/or offspring [62]. In the EPM test and RI test, the MS exhibited increased anxiety and aggression in the adolescent period. These behavioral features of MS agree with other studies, showing MS-induced increased emotionality in male rodents [47]. There are a considerable amount of experimental evidence, which indicates the gender differences in the anxiety-like behavior and stress responses [63]. Usually, male rodents are more vulnerable to early life stress, specifically in anxiety-like behavior, than females. For example, Ito et al. [64] showed the sexually dimorphic patterns in stress responses by showing higher autonomic and anxiety-like behavior in male than in female rodents. They exhibited a lower frequency of entry into and shorter stay in the open arms in the EPM test and an exacerbation of autonomic responses, such as stress-induced hyperthermia and tachycardia, in the early-weaned male rats compared with the normally weaned or early-weaned female rats. For these reasons, this study was conducted using only male mice to avoid confounding issues due to sex.
Although the MS has also been proposed as a depression animal model [65-67], phenotypes other than anxiety, such as depression, did not appear in our MS study (data not shown). It is unclear why MS causes different behavior phenotypes (anxiety/aggression vs. depression) based on the researchers. This difference may be because of a different species background, the duration of separation time, feeding environment and housing conditions, and so on [68]. Therefore, efforts were made to rule out possible factors, such as temperature, ultrasound vocalization, and odors, by keeping many qualifications. We used warm bedding and an isolated room when pups were separated from their dams, and only one proficient researcher was allowed access to the area. As a result, our MS model has consistently shown stable and reliable behavioral results, so this work focuses on increasing anxiety and aggression using MS during the adolescent period. Following these behavioral results, the CORT serum level in adolescent MS mice was compared with the MD.
Excitation-inhibition imbalance (E-I imbalance) is frequently associated with a range of neurological diseases and disorders, such as epilepsy [69], schizophrenia [70], Alzheimer’s disease [71], and autism spectrum disorder [72]. The long-lasting changes in brain interneuron populations have been proposed to contribute to the pathophysiology of many important neuro-psychiatric disorders [73] since it causes E-I imbalance. Furthermore, inhibitory interneurons have been thought to affect the net output of the projection and influence local neural circuits, and contribute to precise regulation of the timing of action potentials. It has been reported that changes in the ventral hippocampal activity owing to changes in E-I balance are related to physiological or pathological anxiety and fear [74-76]. Particularly, the projection from the ventral CA1 and subiculum to the PVN has an anxiolytic effect by inhibiting the HPA axis [77,78], and it has been reported that projection to the lateral septum also decreases anxiety when activated [79]. Alternatively, the excitation of projection neurons to the lateral hypothalamus [80] or DG granule cells [81] increases anxiety, and the hippocampus plays a vital role in the overall anxiety level. The role varies based on the region and the projection pathway. Therefore, there was an attempt to investigate whether the behavioral change was caused by the alteration of the GABAergic population and whether the effect on the population differs for each hippocampus region.
Among different types of GABAergic interneurons, changes were examined in GAD67-expressing interneurons since different kinds of literature have noted GAD67 protein as the molecule most associated with different types of neuropsychiatric disorders [82]. To explore this issue, the number of GAD67 interneurons was investigated in other subfields of the ventral hippocampus of MS and HD. The IHC data showed that the populations of GAD67 interneurons in the ventral hippocampus in MS were 1.3-fold higher than in the HD (Figs. 3 and 4).
Among the diverse subtypes of GABAergic interneurons (PV, somatostatin, calbindin, and neuropeptide-Y) populating the hippocampal formation, PV-positive interneurons play a critical role in emotional states, stress response, and cognitive functions [83]. The PV interneurons are mostly fast-spiking inhibitory neurons that control the circuitry activity of pyramidal cells [10]. Many lines of evidence indicated that the number or density of PV interneurons in the hippocampus is significantly reduced in depression animal models [84,85]. This study showed that the number of PV interneurons in the ventral hippocampus, especially in the DG and CA3 regions, was significantly increased about 2.0-fold in MS compared with HD (Figs. 5 and 6). In the subiculum region, which greatly influences the HPA axis, the change in PV interneurons was insignificant and GAD67-expressing neurons increased. This suggests that the increase in CORT level and anxiety caused by MS is likely due to GABAergic neurons other than PV interneurons.
The hippocampal formation includes the hippocampal gyrus (CA1, CA3, etc.) and the DG. Different subfields of the hippocampus may perform different functions and may also be linked with various mental dysfunctions [30,86,87]. Furthermore, each subfield of the hippocampus has a distinct topographical layer organization and layer-specific connections [88]. In this study, there is an attempt to accurately estimate the number of GAD67 and PV interneurons in different subfields of the hippocampus and each layer. As a result, an increase in the GABAergic population was observed in regions known to reduce anxiety, such as the subiculum, and in regions, such as DG, which increase anxiety when excitability is increased.
Most reports on the effects of each hippocampus on anxiety have only investigated the contribution to the anxiety of a specific population through resection of that region or inhibition of a specific projection using optogenetics. In this study, overall changes in hippocampal GABAergic or PV-positive neuron populations caused by MS were observed. For more physiological conditioning events, such as MS, it is challenging to determine the extent of the effects of projection pathways or neuron subtypes on anxiety for now. The ventral subiculum sends projections to the paraventricular nucleus, a component of the HPA axis that produces corticotropin-releasing hormone, and suppresses the HPA axis activity directly [77] or indirectly [46,89]. The HPA axis is a crucial neuroendocrine stress response system linking the brain with cortisol secretion from the adrenal cortex [90,91]. If the output of the subiculum is inhibited, it can be expected that the HPA axis function will be enhanced and the anxiety and aggression levels will be altered upward [92]. Our data showed an increased number of GAD67 and PV interneurons in MS (Figs. 3–6). The disinhibition circuits to the HPA axis from the hippocampus and/or subiculum can be attributed to increased inhibition of GABAergic interneurons in MS, leading to the increased basal level of serum cortisol and anxiety- and aggressive-phenotype in adolescence. DG act as the main gateway for input in the hippocampus. Regarding anxiety and fear, DG distinguishes context and helps to selectively display fear and anxiety responses in appropriate contexts [81]. When the activity of DG is reduced, the ability to discriminate context is decreased, and fear and anxiety can be generalized [93,94], so it can be expected that an increase in anxiety response will be observed. However, this role of the DG in anxiety is less direct than the subiculum’s function of buffering anxiety responses through the HPA axis. Alternatively, when the physiological conditions of MS are applied, the reduction in output owing to the increase in the GABAergic population of the subiculum directly affects anxiety and aggression through the HPA axis, so it may overwhelm the reduction in output in other parts and have a decisive effect on the behavioral change. In future studies, it is expected that this hypothesis can be confirmed through selective inhibition experiments through techniques, such as optogenetics for the subiculum region with MS. Additionally, it is expected that the description of the GABAergic and PV populations by region and layer will be used in subsequent studies.
Compared with anxiety, little has been reported so far on the effect of the hippocampus on aggression. An association between hippocampal atrophy and increased aggressive behavior has been reported in patients with borderline personality disorder [95]; recently, it has been reported that the ventral hippocampus increases aggression via the ventromedial hypothalamus (VMH) [44]. However, our data indicate an association between an increase in inhibitory neurons and an increase in aggression in the ventral hippocampus, unlike previously reported. It is well known through several reports that the ventral hippocampus, especially the subiculum region, inhibits the activation of the PVN and HPA axes. Alternatively, the recent report that the ventral hippocampal activity increases aggression [44] suggests a role for excitatory projections from the ventral hippocampus to the VMH. This difference may be induced by the difference between the indiscriminate inhibition of the entire ventral hippocampal projection toward the VMH and physiological conditioning, such as MS. Alternatively, it can be explained that when MS is applied, as in anxiety, the inhibition of the subiculum-PVN pathway plays the most decisive role, leading to increased aggression. It is expected that this study will clarify the role of the hippocampus in physiological or pathological changes in aggression, as the effects of the output of numerous pathways starting from the hippocampus on aggression are demonstrated in subsequent studies.
As mentioned in INTRODUCTION, it was previously shown that decreased LTP at the ventral hippocampus CA3 in neonatal MS mice [47], and proposed the possibility that early life stress may influence the developmental neural network of the ventral hippocampus. This study revealed the effect of MS on neural circuits through changes in the number of GABAergic neurons in each region of the hippocampus. In the future, to investigate the functional effects of these changes, it is necessary to examine changes in excitatory and inhibitory signals and changes in synaptic plasticity through electrophysiological recordings in each hippocampal region, especially in the subiculum. Additionally, the increase in IPSC and subsequent decrease in LTP are usually accompanied by changes in the electrophysiological microenvironment in the brain region, particularly alterations in the kinetics of interneurons [96]. Therefore, there is a requirement to look at alterations in the kinetics of interneurons in the ventral hippocampus because the migration and distribution of the brain interneurons can be affected during brain development by early life stress [97].




 XML Download
XML Download