

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Spinal cord injury (SCI) is a trauma that causes persistent neurologic deficits, including paraplegic and neuropathic pain [1,2]. Globally, there are 250,000–500,000 SCI accidents every year, with neuropathic pain occurring in 60%–69% of SCI patients [3,4]. The main causes of neuropathic pain and paraplegia are oxidative stress and inflammation [2,5]. Oxidative stress and inflammatory responses play key roles in the secondary phase of SCI [6]. The stages of SCI remodeling are divided into primary injury, secondary injury, glial scar formation and maturation, and tissue regeneration [7]. Conventional SCI therapy focused on surgery, pharmacological interventions, and rehabilitation to restore lost function and treat neuropathic pain [8].
The current treatment for SCI is centered on the secondary damage of SCI [7]. The effects of the neural stem cell are strongly beneficial in SCI repair [9]. However, there is increasing evidence of the effectiveness of stem cells, which is supported by the paracrine effects of stem cell secretome [10]. Stem cell secretomes have potential anti-neuroinflammatory, antiapoptotic, angiogenesis, and neurogenesis effects [10,11]. Pajer et al. [8], stated that intravenously administered secretome and the lesion-induced secretome modulated SCI symptoms and functional recovery through inflammatory cytokines supplemented with neurotrophic factors. However, further studies are necessary to prove the positive effects of intraspinal use. Zhou et al. [12] stated that neural stem cell secretome (NSC-S) significantly promoted the polarization of microglia from the proinflammatory M1 to the anti-inflammatory M2 phenotype, and reduced the production of proinflammatory cytokines. NSC-S promoted peroxisome proliferator-activated receptor gamma pathway activation.
Despite extensive research, recently, the pathophysiological mechanisms in the development of neuropathic pain have not been clearly revealed [13]. The role of human neural stem cell (HNSC)-secretome in neuropathic pain is unclear. The mechanism of the HNSC-secretome improvement of neuropathic pain and locomotor function in SCI remains unclear. The research question of this study was in regards to the effect and the mechanism of HNSC-secretome in improvement of neuropathic pain and locomotor fuction in SCI. Accordingly, the aim of the study is to evaluate the mechanism of HNSC-secretomes’ improvement of neuropathic pain and locomotor function in SCI by analyzing F2-Isoprostanes, tumor necrosis factor (TNF)-α, matrix metalloproteinase (MMP)-9, transforming growth factor (TGF)-β, and brain derived neurotrophic factor (BDNF).
Go to : 
MATERIALS AND METHODS
1. Ethics statement
The study’s protocol was reviewed and approved by Faculty of Dentistry, University of Jember (REC.1462/UN25.8/KEPK/DL/2021). All rats were approved by the animal health office (No.503/A.1/0005. B/35.09.325/2020).
2. Study design
This research was a true experimental study. The sample size was calculated using the Lemeshow formula (n = 5 rats), with correction factors of 20%. The rats were randomly grouped into the following three groups: the treatment group (five experimental rats had SCI with HNSC-secretome), the control group (five experimental rats had SCI without HNSC-secretome), and the normal group (five experimental rats did not have SCI and did not get HNSC-secretome). The treatment group received a 30 µL HNSC-secretome intrathecal injection at the T10 level three days after the SCI and laminectomy [8,14]. All groups were replicated five times. The authors observed the study over 28 days [15]. The independent variable of the study is HNSC-secretome, where as the dependent variables are neuropathic pain, locomotor function, TNF-α, F2-Isoprostane, MMP-9, TGF-β, and BDNF.
3. Preparation of the HNSC-secretome
The material of HNSC-secretomes was obtained by buying in installation of stem cells and tissue bank (REC.0059/KEPK/IX/2020). The neural stem cells were counted as 2 × 5 × 106, derived from Adipose-Mesenchymal Stem Cells. HNSC-secretomes are characterized by the presences of neural cells and Nestin markers.
4. Rats and SCI models
We used a reliable Rattus norvegicus SCI contusion–compression model [16]. The adult male Rattus norvegicus pure strain Sprague Dawley rats were three to four months old and weighed 300–350 g. Acclimatization was carried out for seven days by one chemist and two veterinarians. Animals were kept in separate cages, one cage containing one rat, using a plastic box, 45 × 30 × 15 cm3 with woven wire as a cover. The floor mat was covered with wood shavings and there was an underpad to absorb urine and maintain moisture. The room comfort was provided by air conditioning to maintain a room temperature of 22℃ ± 2℃ and a humidity of 50%–70%. An exhaust fan was used to remove the smell of ammonia. The environment was a quiet room with a 12-hour light and dark cycle. The light sources was a 300 lux electric lamp 1 meter from the floor. The cage was cleaned every three days with soap and running water. Feed was 30–35 g of pellets (10% of bodyweight) and 30–35 mL of mineral water (10% of bodyweight).
Contusion–compression of the spinal cord was conducted using the commercially available spinal cord impactor aneurysm Yasargil Clip, with a length of 7 mm and a load of 65 g (equivalent to 150k Dyne). The animals were anaesthetized using ketamine (75 mg/kg) and acepromazine (3 mg/kg) intraperitoneal [17]. The animals were placed on a fixation board in a prone position, and about a 3-cm portion of their back hair was shaved. The operating area was disinfected with 10% betadine and 75% alcohol. The surgical level was marked by tracing the rib at the level of the T10 spinous process using a 2 cm skin incision. A partial laminectomy was conducted the T10–T11 level to expose the level T10 spinal cord. The tip of the titanium aneurysm Yasargil Clip was placed at a distance of 1 mm from the spinal cord anteriorly and posteriorly. The spinal cord was impacted suddenly for 60 seconds by retraction of the tip using an applicator. It produced a SCI contusion-compression model with the dura appearing cloudy white and impacted flat. The operating field was cleaned using saline, and the muscle and skin were sutured together in layers.
Three days post-injury, the rat model, in both the treatment and control groups, were completely paraplegic. The treatment group was administered an intrathecal injection of 30 µL HNSC-secretome under general anesthesia, which was centered at the site of the injury and 1.5–2 mm deep from the dura to the subarachnoid space with a tilt angle of 30°–40° using a 50 µL Hamilton Syringe. The animals received physiological saline subcutaneous (s.c.), tolfenamic acid 4 mg/kg s.c. and enrofloxacin 10 mg/kg s.c. and were placed under a 5 W heating lamp. Manual bladder drainage was conducted twice daily until micturition was normal.
5. Neuropathic pain assessment
The Rat Grimace Scale (RGS) was used to observe the neuropathic pain. The RGS is an accurate, sensitive, and reliable method of identifying and measuring pain in mice [18]. The score was measured before surgery and on days 21 and 28 after surgery. It takes animal facial expressions that consists of four measures, namely orbital/eye lifting, nose/cheek protrusion, ear changes and whisker position. It is based on scoring the presence of each action unit, which consists of 0, 1, and 2. There score is “0” if there is no change (normal), “1” if the change is moderate, and “2” if the change is clear [19]. Each rat was observed for nine minutes in each testing session using a video camera. The assessment was carried out every 15 seconds, followed by a 15-second interval. Every three minutes, the score was averaged, and then the total score was averaged as the final score.
6. Locomotor assessment
The Basso, Beattie, and Bresnahan (BBB) scores were utilized to measure the locomotors recovery. To assess the locomotor expression, the BBB open-field test was performed on days 1, 3, 7, 14, 21, and 28 after injury [16]. The BBB measures the tail, body, legs, trunk stability, limb movement, and toe clearance, all of which are examined to measure the locomotor abilities [20,21]. The score shows a range of numbers between 0 and 21. A score of 0 is no movement, and a score of 21 is normal movement without a locomotor disorder. Briefly, scores 0–7 rank the early phase of recovery indicating movement of the primary 3 joints (hip, knee, and ankle), scores 8–13 describe the intermediated phases of recovery from weight bearing stance to coordinated stepping, and scores 14–21 rank the late phase with the return of toe clearance, paw position, and trunk stability [22].
7. Preparation of the spinal cord tissue for immunohistochemical assessment
The termination of the animals was carried out on day 28 through the induction of inhalation anesthetics. The sterilization of the operating area was carried out using 70% alcohol and 10% betadine, then put on sterile cloths. A transverse incision was made on the left side at the level of the ribs reaching the T8–T12, then 5 cm of spinal cord tissue was separated from the vertebral column and marked at the cranial end. The spinal cord tissue materials were put in a pot and fixed in a 10% buffer formalin.
8. Immunohistochemical assessment
The immunohistochemistry of biomarkers MMP-9, TGF-β, and BDNF, were evaluated using quantitative measurements. Five specimens of spinal cord tissue were taken from animals in each group. The average value of ten fields of view was observed, and each field of view had an area of 625 µ2 with a magnification of 400×. Macrofag cell was observed for MMP-9 and TGF-β, whereas neuron cells were observed for BDNF. Immunohistochemical operational procedures were as follows: the specimens were immersed in the xylol solution for 3–5 minutes, then in absolute ethanol for 1–3 minutes, and finally in 70% ethanol for 1–3 minutes. They were then washed 3 times with Aquabidest, and the edge of the slide was cleaned with a tissue. They were then dropped with H2O2 3%, incubated at room temperature for 10 minutes, washed 3 times in phosphate-buffered saline (PBS), and the edge of the slide was again cleaned with a tissue. They were then dropped with Trypsin 0.025%, and incubated at 37°C for 6 minutes, washed 3 times in PBS, and the edge of the slide again cleaned with a tissue. Specimens were then dropped with Ultra V Block and incubated at room temperature for 5 minutes, with the edge of the slide cleaned again (no need to wash). They were then dropped with monoclonal antibody which has been diluted (1:100) and incubated at room temperature for 25–30 minutes, washed with PBS 3 times, and the edge of the slide cleaned with a tissue. Drops of biotin, incubation at room temperature for 10 minutes, washed with PBS 3 times, and the edge of the slide cleaned with a tissue. Specimens were then dropped with horseradish peroxidase polymer, incubated at room temperature for 10 minutes, washed with PBS 3 times, and the edge of the slide cleaned with a tissue. They were then dropped with diaminobenzidine chromogen (20 μL/1 mL substrate), incubated at room temperature for 5–15 minutes in a dark room, washed Aquabidest 3 times, cleaned, painted with Meyer Hematoxylin at room temperature, then incubated for 6–15 minutes, washed in running water 3 times, and finally soaked in water for 10 minutes, drained, and mounting.
9. Enzyme-linked immunosorbent assay
The specimen for enzyme-linked immunosorbent assay (ELISA) was collected from the cardiac blood. The TNF-α analysis used the serum, and F2-Isoprostanes used plasma for analysis. The ELISA kit for the TNF-α analysis used the Sandwich-ELISA principle. The ELISA kit for the F2-Isoprostanes analysis used the Competitive-ELISA principle. TNF-α and F2-Isoprostanes were evaluated using quantitative measurements.
10. Statistical methods
The data in this research is reported as the mean ± standard deviation. SPSS software version 25 (IBM Co., Armonk, NY) was utilized for analysis. The normality was analyzed using the Shapiro–Wilk test, while the homogeneity was determined using the Levene test. A P value less than 0.05 was considered statistically significant. The differences between groups were analyzed using ANOVA followed by the Tukey HSD test. A P value of < 0.05 was considered statistically significant.
Go to :
RESULTS
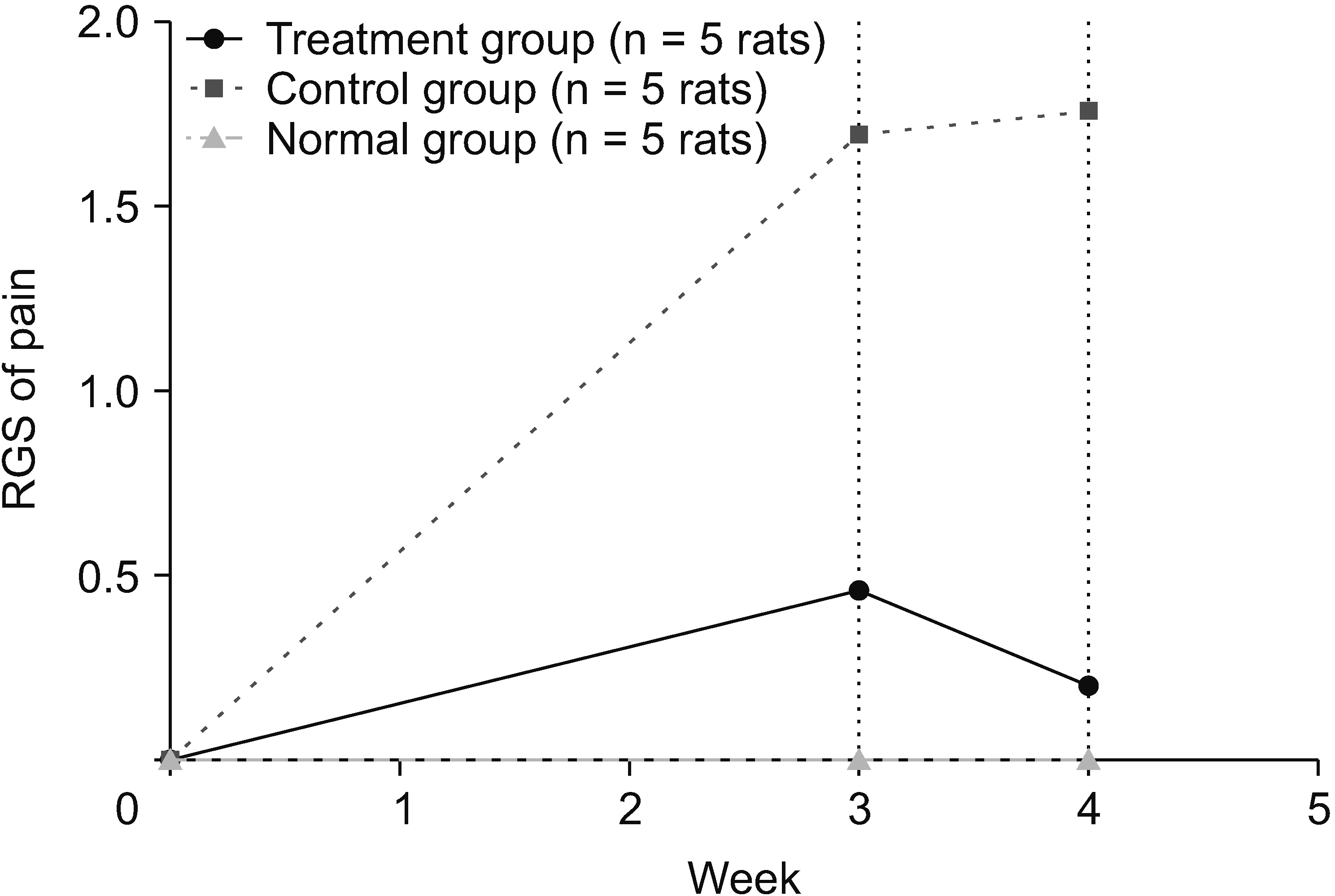
The rats were evaluated over a period of four weeks to determine the neuropathic pain reduction using the RGS. Neuropathic pain decreased on day 21 and continued to decrease until day 28. Based on the independent t-test, the results on days 21 and 28 showed that the treatment and control groups had different consecutive significances of P < 0.001 and P < 0.001, respectively. The mean differences (MD) on days 21 and 28 between the treatment and control groups were sequentially MD = 1.556 and MD = 1.238, respectively. The treatment group had a much higher effect on reducing the value of neuropathic pain in the rat SCI subacute contusion–compression model (Fig. 1).
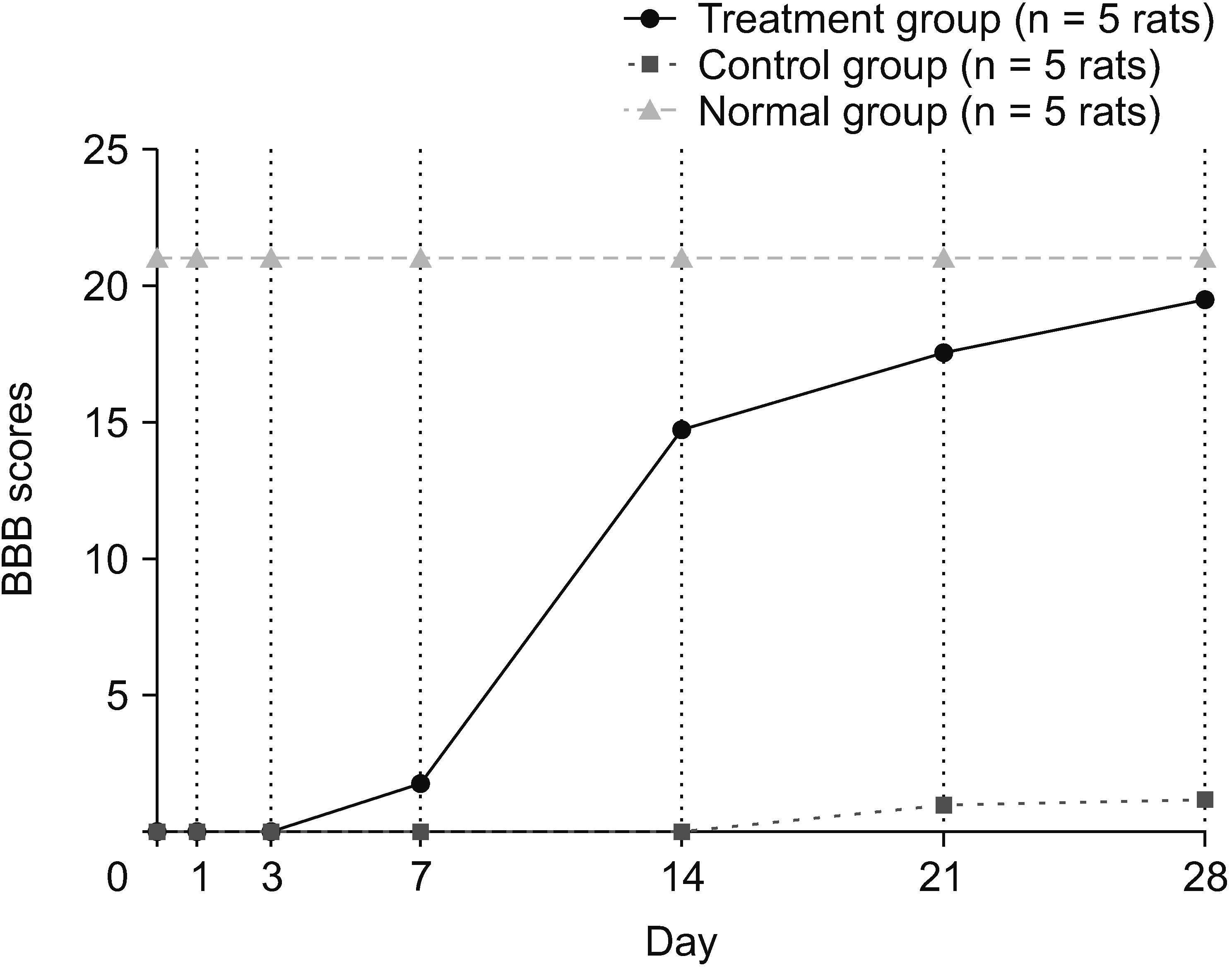
The rats were evaluated over a period of 28 days to determine motor recovery. Locomotor recovery was recorded on days 1, 3, 7, 14, 21, and 28. The mean value (x̄) BBB score of the treatment group was 19.60, whereas in the control group, it was 1.20. The MD in the BBB scores was 18.4 (BBB score 0–21). Based on the independent t-test, the control and treatment groups were different, with a significance of P < 0.001. The treatment group had a much higher effect on improving the value of the locomotor recovery in the rat SCI subacute contusion–compression model (Fig. 2).
Based on the mean value, the level of F2-Isoprostane in the treatment group was significantly different from the control group (P = 0.001). The number of cells expressing F2-Isoprostane in the treatment group (x̄ = 255.151) was smaller than control group (x̄ = 377.649) (Table 1).
Table 1
Comparison of the intergroups of biomarker F2-Isoprostanes
![]()
Based on the mean value, the level of TNF-α in the treatment group was significantly different from the control group (P = 0.032). The number of cells expressing TNF-α in the treatment group (x̄ =172.048) was smaller than in the control group (x̄ = 214.705) (Table 2).
Table 2
Comparison of the intergroups of biomarker tumor necrosis factor (TNF)-α
![]()
Based on the mean value, the number of cells expressing MMP-9 in the treatment group was significantly different from those in the control group (P = 0.038), and the number of cells expressing MMP-9 in the treatment group (x̄ = 1.200) was smaller than in the control group (x̄ = 3.140) (Table 3 and Figs. 3, 4).
 | Fig. 3A bar graph representing the comparison of cell counts between groups in matrix metalloproteinase-9 cytokine.
|
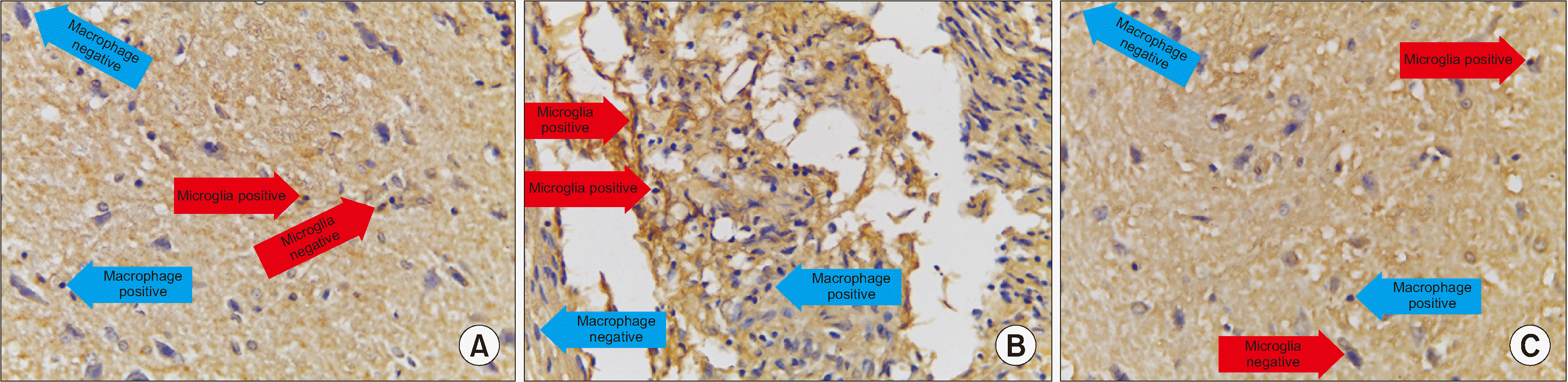 | Fig. 4We observed immunohistochemical matrix metalloproteinase (MMP)-9 average value of 10 field of views, every field of view have 625 µ2 with 400× magnification. (A) Treatment group, (B) Control group, (C) Normal group. Microglia (red arrow) are small round cells, solid nuclei and give a positive reaction with anti MMP-9 indicated by brown color. While macrophage (blue arrow) cells are large, vesicular nucleus, and sometimes elongated resembling fibroblasts (macrophages like fibroblast) and give a positive reaction with anti MMP-9 indicated by brown.
|
Table 3
Comparison of the intergroups of biomarker matrix metalloproteinase-9
![]()
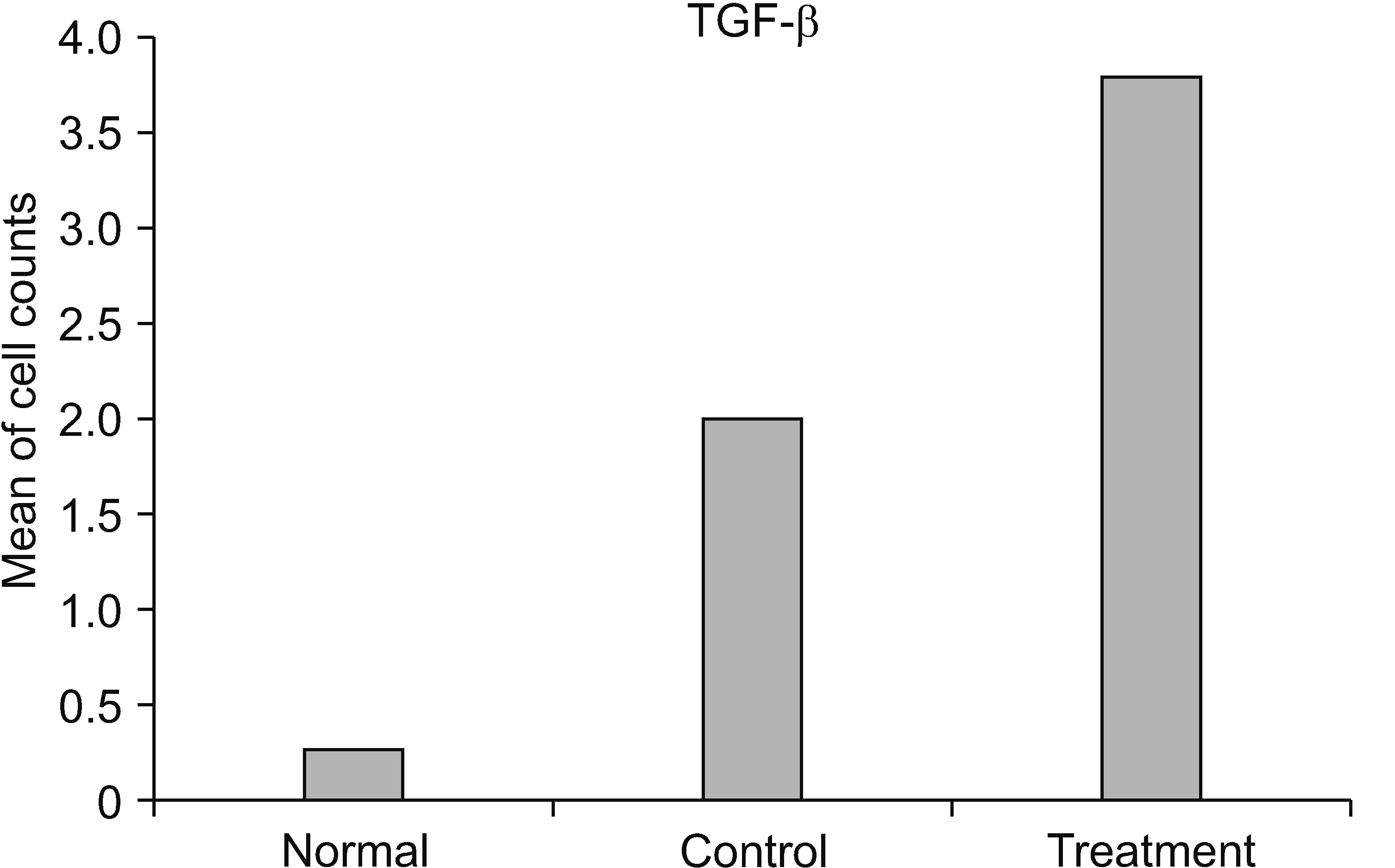
Based on the mean value, the number of cells expressing TGF-β in the treatment group was significantly different from those in the control group (P < 0.001). Moreover, the number of cells expressing TGF-β in the treatment group (x̄ = 3.800) was greater than those in the control group (x̄ = 2.000) (Table 4 and Figs. 5, 6).
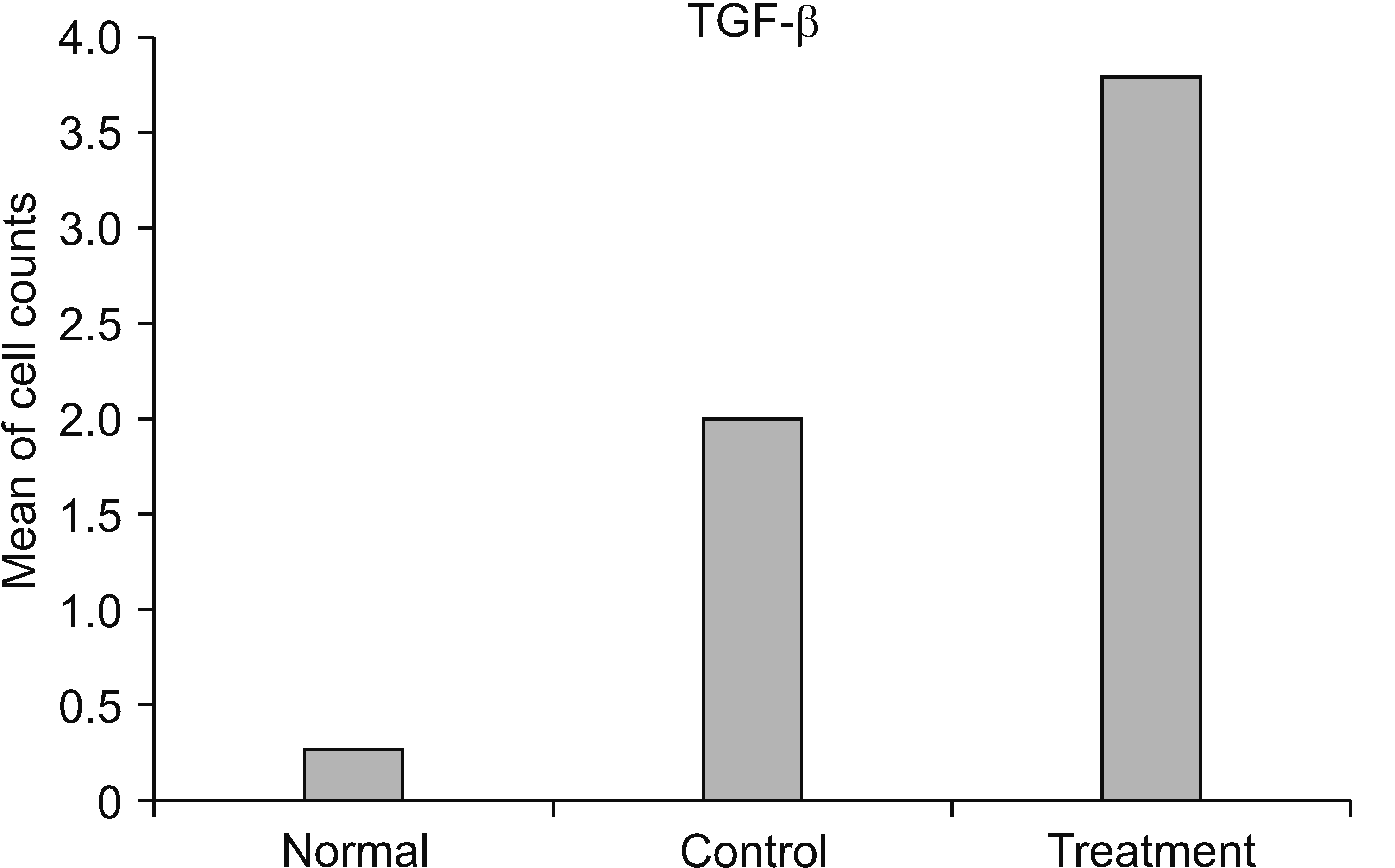 | Fig. 5A bar graph representing the comparison of cell counts between groups in transforming growth factor (TGF)-β cytokine.
|
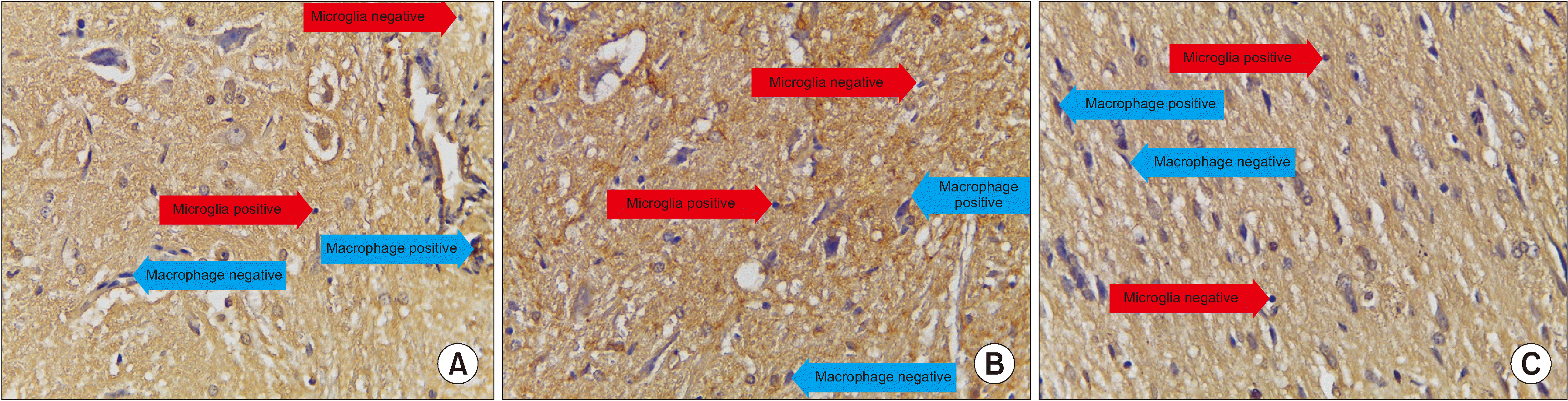 | Fig. 6We observed immunohistochemical transforming growth factor (TGF)-β average value of 10 field of views, every field of view have 625 µ2 with 400× magnification. (A) Treatment group, (B) Control group, (C) Normal group. Microglia are small round cells, solid nuclei and give a positive reaction with anti TGF-β indicated by brown color. While macrophage cells are large, vesicular nucleus, and sometimes elongated resembling fibroblasts (macrophages like fibroblast) and give a positive reaction with anti TGF-β indicated by brown.
|
Table 4
Comparison of the intergroups of biomarker transforming growth factor (TGF)-β
![]()
Based on the mean value, the number of cells expressing BDNF in the treatment group was significantly different from those in the control group (P = 0.001), and the number of cells expressing BDNF in the treatment group (x̄ = 1.980) was greater than in the control group (x̄ = 0.560) (Table 5 and Figs. 7, 8).
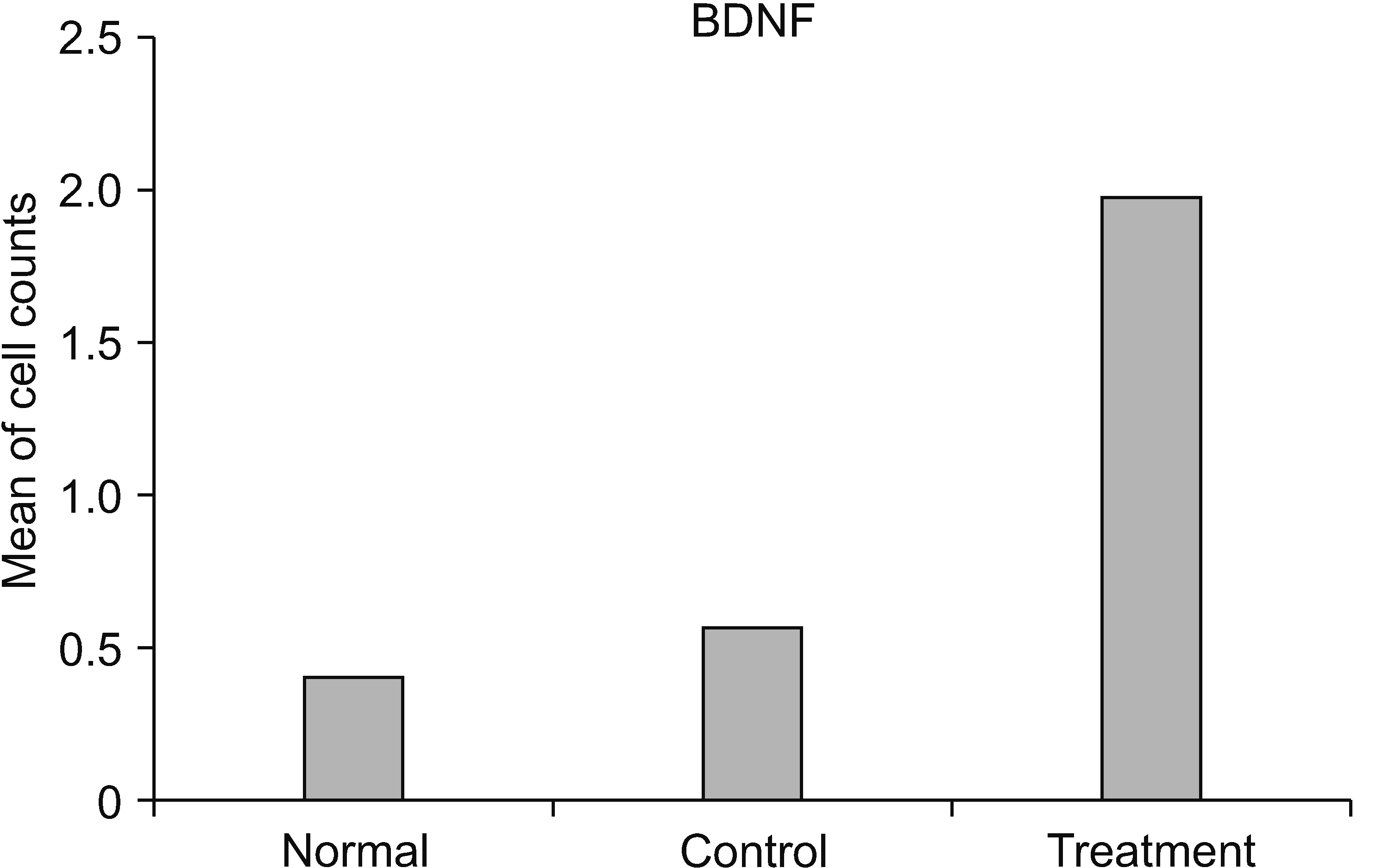 | Fig. 7A bar graph representing the comparison of cell counts between groups in brain derived neurotrophic factor (BDNF).
|
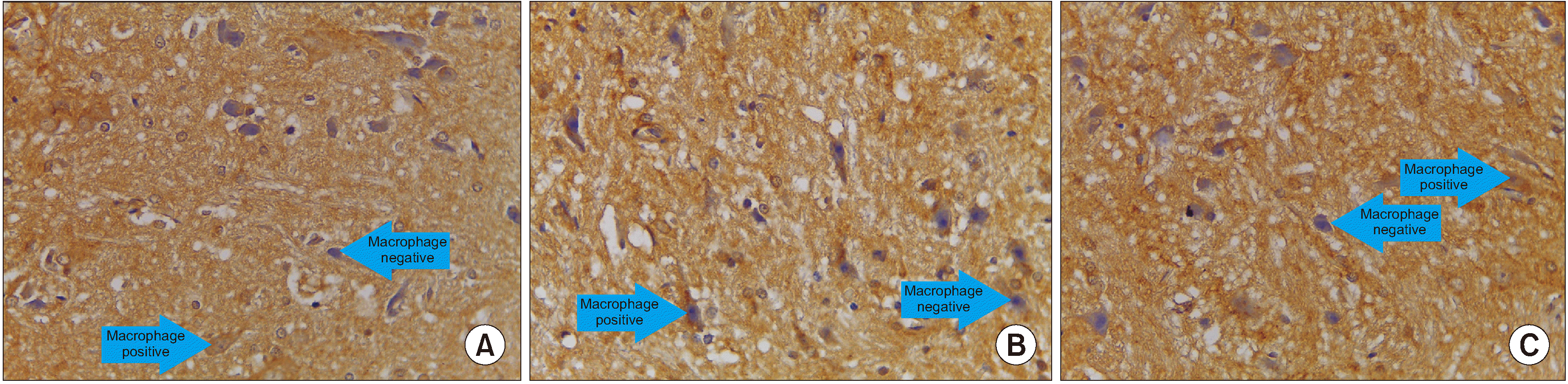 | Fig. 8We observed immunohistochemical brain derived neurotrophic factor (BDNF) average value of 10 field of views, every field of view have 625 µ2 with 400× magnification. (A) Treatment group, (B) Control group, (C) Normal group. Macrophage cells are large, vesicular nucleus, and sometimes elongated resembling fibroblasts (macrophages like fibroblast) and give a positive reaction with anti BDNF indicated by brown.
|
Table 5
Comparison of the intergroups of biomarker brain derived neurotrophic factor (BDNF)
![]()
Go to :
DISCUSSION
HNSC-secretome, which is administered in the rat SCI subacute contusion–compression model, improved neuropathic pain and locomotor function more significantly compared to the control group. Neuropathic pain was evaluated starting on days 21 and 28. This was in accordance with the research conducted by Leung et al. [23], Wu et al. [24], and Hatch et al. [25], who stated that pain before day 21 was a combination of various sources of pain, such as surgical wound pain, surgical stress and anesthesia, and post laminectomy pain. In another study found spontaneous neuropathic pain and supraspinal pain up to 5 weeks after cervical hemicontusion using the RGS score [26]. Locomotor function improvement started from day 7 and continued to day 28 post-injury. Cunningham et al. [27] stated that there are 4 studies which observed locomotor function for 4 weeks, 4 studies which observed for 6 weeks, and one study which observed for 10 weeks. To date, the time for full recovery process of locomotor function is not yet known.
Immunohistochemistry showed that HNSC-secretome treatment increased TGF-β and BDNF but decreased MMP-9. ELISA showed that HNSC-secretome treatment decreased F2-Isoprostane and TNF-α. Specifically, HNSC-secretome showed neurogenic properties that were regulated by BDNF and TGF-β upregulation as well as anti-inflammatory and antioxidative stress by decreasing F2-Isoprostanes and TNF-α, and increasing TGF-β. Moreover, HNSC-secretomes can maintain the extracellular matrix (ECM) of SCI by reducing the matrix degradation effect of MMP-9 and increasing the collagen formation effect of TGF-β. This may reduce glial scar formation and neuropathic pain.
The administration of secretomes in the brain could induce macrophages to secrete TGF-β, MMP-9, and BDNF [11]. Inflammation contributes to secondary SCI when the inflammation is slower and longer than the peripheral nerve. As a result, the healing process of SCI is becoming more complicated [28]. In this research, the role of HNSC-secretome was clear in repair of SCI, through several biomarkers and the improvement of neurological functions.
This research showed that HNSC-secretomes decrease F2-Isoprostanes. Antioxidant properties have been reported to be effective in treating neuropathic pain [29]. Jia et al. [30] stated that alleviating the oxidative stress of the secondary injury process may represent an effective strategy for therapeutic intervention in SCI. Dos Santos et al. [31] stated that NSC secretomes have an important antioxidant role in reducing F2-Isoprostane by inhibiting endoperoxidase, arachidonic acid, and reactive oxygen species (ROS). F2-Isoprostanes is increased by oxidative stress and is derived from the free radical induction of neuronal arachidonic acid in membrane phospholipids and lipoproteins [32]. ROS that are derived from microglia and leukocytes (macrophages and neutrophils) are secreted by an enzyme system (nicotinamide adenine dinucleotide phosphate oxidase, myeloperoxidases, cyclooxygenase, and xanthine oxidase) [32]. Polyunsaturated fatty acids (arachidonic acid, docosahexaenoic acid) target free radicals that produce F2-Isoprostanes, which is an expression of lipid peroxidation [33]. Additionally, they activate nuclear factor kappa-B (NFκB), which causes an inflammatory process; they release gamma-aminobutyric acid; they increase intracellular Ca2+ and Na+; they disrupt mitochondrial function, excitotoxicity and neuronal malfunction; and finally, they result in neuronal death [34].
This research showed that HNSC-secretome decreases TNF-α. TNF-α promotes the recruitment of neutrophils and macrophages to the site of the injury, cleans up debris and necrotic tissue, and secretes pro-inflammatory cytokines [35]. Additionally, neutrophils and macrophages secrete anti-inflammatory cytokines, such as interleukin (IL)-10 and TGF-β, and induce anti-inflammatory regulatory T-cells [36]. Although TNF-α decreases on the seventh day after trauma, eventually, TNF-α can stimulate synaptic plasticity and glutamatergic transmission and contribute to the survival of injured neurons [37]. TNF-α can trigger various cell signaling pathways, which protect against excitotoxicity-induced cell death and stabilize calcium homeostasis due to the promoted expression of the calcium-binding protein calbindin [37].
This research showed that HNSC-secretome decreases MMP-9. MMP-9 are endopeptidases that contribute to SCI repair by degrading ECM molecules and depleting MMP-9 [38]. MMP-9 can be promoted by ROS and pro-inflammatory mediators, such as TNF-α and IL-1β through the NFκB pathway [6]. MMP-9 is inhibited by tissue inhibitors of metalloproteinases (TIMP), whereas TIMP are inhibited by TGF-β [39]. MMP-9 is responsible for the early degradation of the basal lamina components, which leads to blood-spinal cord barrier disruption and initiates leukocyte infiltration, progressive neuroinflammation response, and demyelination [40]. This system modulates macrophage invasion and myelin destruction, which has an important role in neuropathic pain [2]. Moreover, MMP-9 contributes to neuropathic pain through glial scar formation [41].
This research showed that the HNSC-secretome increases TGF-β. TGF-β, as an anti-inflammatory cytokine, induces regulatory T-cell differentiation and promotes macrophage polarization towards the M2 phenotype [36]. It prevents M1 from synthesizing with proinflammatory TNF-α, IL-1β, IL-6, indicible nitric oxide synthase, ROS, glutamate, and proteases [42]. Moreover, TGF-β induces fibroblasts to secrete collagen, fibronectin, and proteoglycans for new ECM formation [43]. TGF-β overcomes the matrix degradation, which is caused by the effects of MMP-9 [39]. TGF-β is involved in nerve repair and regeneration, has been widely observed to combat nerve demage, and stimulate cellular survival, growth, proliferation, differentiation, and the invasion of neurons and glial cells [36].
This research showed that HNSC-secretome increases BDNF. Shahsavari et al. [44] stated that BDNF is a molecule that is involved in various neurophysiological functions such as nociception, learning, and memory. BDNF is a neurotrophin that is important in the regulation of several neurogeneses, such as collateral growth promotion, neuronal branching, dendrite formation, and synaptic plasticity [45]. Studies have demonstrated that increased BDNF in the spinal cord protects motor neurons from degeneration, increases axonal and collateral sprouting, and upregulates markers of synaptic plasticity [46]. BDNF could support cell survival and adaptive plasticity after SCI [47]. Moreover, BDNF has been shown to stimulate axonal growth, enable locomotor function in spinally transected animals, and support respiratory function after cervical trauma [48].
The result of this study demonstrated the mechanism of HNSC-secretome in improving neuropathic pain and locomotor function in SCI through antioxidant, anti-inflammatory, anti-matrix degradation, and neurotrophic activities. Although cell transplantation remains intensely studied, surprisingly little is known about the mechanisms through which transplanted cells promote repair and mediate functional improvements [7,49,50]. The limitations of this study are a small sample size and the difficulty in management of paraplegic experimental animals with impaired urinary, bowel function, which ends with death. Future research could investigate the histological features, neurologic functions, and biomarkers. Clinical relevance of this research requires animal experiments and clinical trial. Finally, HNSC-secretomes can be used clinically, mass produced, and affordable.
Go to :
 XML Download
XML Download