

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
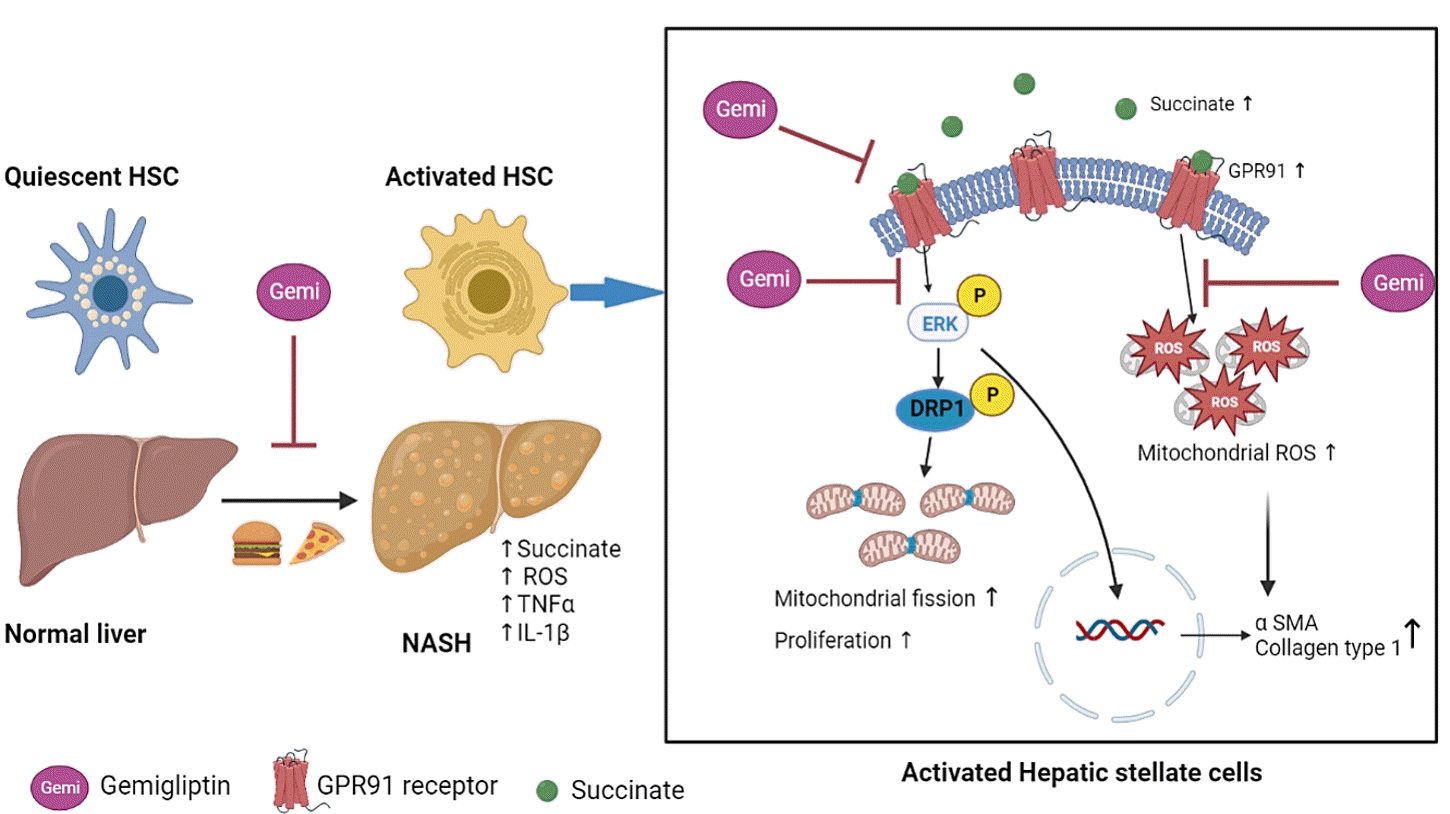
Liver fibrosis is a common feature of several chronic liver diseases, such as hepatitis B, hepatitis C, alcoholic hepatitis, autoimmune disorders, and nonalcoholic steatohepatitis (NASH), regardless of their etiology, and is characterized by excessive accumulation of extracellular matrix (ECM) [1,2]. Hepatic stellate cells (HSCs) are the principal collagen-producing cells involved in the production of ECM proteins under conditions of persistent hepatic injury [3]. In the normal liver, HSCs reside in the liver within the space of Disse in a quiescent form, storing vitamin A in lipid droplets. However, following chronic liver damage, HSCs become activated or transdifferentiate into myofibroblast-like cells with proliferative, contractile, migratory, pro-inflammatory, and fibrogenic properties, suggesting that HSC activation is the central mechanism underlying liver fibrogenesis [4,5].
Succinate, an intermediate of the tricarboxylic acid cycle, has emerged as an important signaling molecule in liver fibrosis [6]. Although succinate is normally produced in mitochondria, accumulated succinate is released into the extracellular space and acts as a paracrine and endocrine effector by binding its receptor, known as G-protein coupled receptor 91 (GPR91), in HSCs. GPR91 is exclusively expressed in HSCs, and once stimulated by succinate it activates the downstream mitogen-activated protein kinase (MEK1/2) by further promoting the phosphorylation of extracellular signal-regulated kinase 1/2 (ERK1/2) and c-Jun [7]. Subsequently, the expression of nuclear transcription factors and production of α-smooth muscle actin (α-SMA) stimulates the activation, proliferation, and migration of HSCs. In addition it inhibits apoptosis in HSCs and induces inflammation, leading to liver fibrosis and NASH [8,9].
Mitochondria are highly dynamic organelles that generate energy and preserve cellular homeostasis through their production of metabolites. Mitochondria are widespread in the cytosol and regulate various cellular physiological functions [10]. Mitochondria continuously alter their morphology to adapt to a new metabolic milieu, responding to metabolic disturbances through fusion and fission cycles [11-13]. Mitochondrial dynamics are engaged in a variety of biological activities, including redox regulation and cell proliferation [14]. Dynamin-related protein 1 (DRP1), a cytosolic GTPase, is a critical regulator of mitochondrial fission. DRP1 is recruited from the cytoplasm to the mitochondrial outer membrane upon activation, where it binds a mitochondrial membrane adaptor, mitochondrial fission factor (MFF), to cause mitochondrial fission [15]. DRP1 activation is regulated by phosphorylation. ERK2 increases the translocation of DRP1 to mitochondria and promotes mitochondrial fission by phosphorylating DRP1 at serine 616 [16].
Liver fibrosis and chronic liver diseases progress over a long period of time. Therefore, therapies for these conditions should be well tolerated over time, with good targeting to the liver and few undesirable side effects on other organs. However, no antifibrotic agent has yet been approved in clinical practice [17]. Thus, an alternative approach may be to identify an existing clinical compound that shows antifibrotic activity until new drugs become widely available [18,19]. Dipeptidyl peptidase-4 (DPP-4, also known as CD26) is a serine protease that is ubiquitously expressed in many cell types and occurs in soluble form in the circulatory system as an enzyme that breaks down incretins [20-22]. DPP-4 is also highly expressed in the liver and circulation of patients with nonalcoholic fatty liver disease and NASH. Increasing evidence suggests that DPP-4 is highly involved in the development of chronic liver diseases [23,24]. DPP-4 inhibitors (DPP-4Is), which function as glucose-lowering agents by inhibiting DPP-4, have been used for the treatment of type 2 diabetes mellitus. Previous studies have demonstrated that DPP-4Is attenuated hepatic fibrosis by suppressing activated HSCs in rats [25] and prevented NASH-associated liver fibrosis in mice, independent of their antidiabetic effects [26]. Moreover, DPP-4I has been found to attenuate brain mitochondrial dysfunction and decrease brain oxidative stress levels in high-fat diet-fed rats [27].
In this study, we investigated the correlation between succinate and mitochondrial dysfunction in HSCs and the ability of gemigliptin to attenuate succinate-induced mitochondrial dysfunction and HSC activation.
Go to : 
METHODS
Materials
The reagents and antibodies used in this study were obtained from the indicated suppliers: succinic acid, N-acetyl-L-cysteine (NAC) from Sigma (St. Louis, MO, USA); gemigliptin from LG Chem (Seoul, Korea); primary antibodies: ERK1/2, p-ERK1/2, DRP1, p-DRP1 (Serine 616), MFF, and p-MFF from Cell Signaling Technology (Richmond, CA, USA); p-DRP1 (Serine 616) from Invitrogen (Waltham, MA, USA), glyceraldehyde 3-phosphate dehydrogenase (GAPDH) from GeneTex (Irvine, CA, USA), GPR91 from Santa Cruz Biotechnology (Dallas, TX, USA), α-SMA from Abcam (Cambridge, England), and collagen 1 from Sigma-Aldrich (St. Louis, MO, USA).
Cell culture
Immortalized human HSCs (LX-2 cells) were kindly provided by professor Ja June Jang, Seoul National University. The LX-2 cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM), which included 10% fetal bovine serum and 100 U/mL penicillin/streptomycin.
Western blot analysis
Cells and liver tissues were lysed in a radioimmunoprecipitation assay buffer (Atto Corporation, Tokyo, Japan). The protein content in the total cell lysates was determined by using a BCA protein assay kit (Thermo Scientific Pierce, Rockford, IL, USA). Sodium dodecyl sulfate polyacrylamide gel electrophoresis was used to separate equal amounts of total proteins, which were then transferred to polyvinylidene difluoride membranes (Millipore, Bedford, MA, USA). After transfer, the membrane was incubated in 5% nonfat dry milk in Tris buffered saline containing 0.1% Tween-20 (TBST) for 1 hour at room temperature. After that, the membranes were washed three times with TBST and incubated with primary antibodies overnight at 4°C or for 2 hours at room temperature, according to the manufacturer’s instructions. The membranes were then incubated with appropriate horseradish peroxide-conjugated secondary antibodies at room temperature for 90 minutes. Subsequently, the blots were incubated with WestGlow PICO PLUS chemiluminescent substrate (Biomax, Seoul, Korea) and images of the blots were captured using a ChemiDoc Imaging System (BioRad Laboratories Inc., Hercules, CA, USA). Image Lab software version 6.1.0 (Bio-Rad Laboratories Inc.) was used to calculate the band intensities.
Cell proliferation assay
LX-2 cells were seeded in 24-well plates at a density of 0.5×104 cells per well and cultured at 37°C for 24 hours. The medium was replaced with fresh DMEM containing succinic acid (1,600 μmol/L) with or without gemigliptin and incubated for 24 or 48 hours. Viable cell numbers were estimated using cell counting kit-8 (CCK-8) (Dojindo Molecular Technologies Inc., Rockville, MD, USA), with 50μL of the CCK-8 reagent added to each well. The plates were incubated for 1 hour at 37°C, and the absorbance value (optical density) of each well was measured at 450nm according to the manufacturer’s instructions.
Measurement of reactive oxygen species
Intracellular reactive oxygen species (ROS) in LX-2 cells treated with succinic acid (1,600 μmol/L) with or without gemigliptin, and 5 mM NAC for 1 hour were analyzed using DCFDA/H2DCFDA (Abcam, Cambridge, UK, ab113851). After treatment for 1 hour with reagents, LX-2 cells were stained with DCFDA (20 μM) for 45 minutes at 37°C in the dark. Images were obtained using a fluorescence microscope (Olympus, Tokyo, Japan).
Mitochondrial ROS production was examined using the superoxide indicator MitoSOX Red (M36008, Invitrogen). LX-2 cells were plated and treated with succinic acid (1,600 μmol/L) with or without gemigliptin for 1 hour. Next, the cells were washed with phosphate-buffered saline and then incubated with 5 μM MitoSOX reagent and MitoTracker Green (Beyotime, Shanghai, China) for 30 minutes at 37°C in the dark. Images were obtained using a confocal scanning microscope (Olympus).
Transmission electron microscopy
LX-2 cells were plated and treated with succinic acid (1,600 μmol/L) with or without gemigliptin for 1 hour. After incubation, the cells were fixed with 2% glutaraldehyde and 2% paraformaldehyde in phosphate buffer (pH 7.4) for 1 hour at 4°C, and then post-fixed in osmium tetroxide for 40 minutes at 4°C. The cells were dehydrated in graded concentrations of ethanol solutions. The cells were treated with a graded propylene oxide series and embedded in Epon. Ultrathin sections (80 nm) were cut from each block and placed on a copper grid. Finally, the samples were stained with uranyl acetate, followed by lead citrate and then observed using a transmission electron microscope (JEOL-2100F, USA, 200 kV) at the Korea Basic Science Institute (Chuncheon, Korea).
Animal experiments
The animal study was approved by the Institutional Animal Care and Use Committee of the National Kangwon University (KW-170517-1) and was conducted in accordance with the applicable guidelines. Four-week-old male C57BL/6N mice were purchased from Doo Yeol Biotech (Seoul, Korea). The 4-weekold mice were acclimated to laboratory conditions for 1 week. All mice were housed in the Kangwon National University animal care facility at ambient temperature (22°C±1°C) with a 12/12-hour light/dark cycle and with free access to water and food. The mice were randomly divided into three groups: (1) the control group was fed a control diet (2018S, Envigo, Indianapolis, IN, USA); (2) the NASH model group was fed a high-fat, high-cholesterol (HFHC) diet; and (3) the treatment group was fed an HFHC diet mixed with 400 mg/kg/day of gemigliptin. The HFHC diet contained 38.25 g of 2018S (a fixed formula diet containing a minimum of 18% protein and 5% fat), 1.25 g of cholesterol, 60 g of cocoa butter, and 0.5 g of cholate. The study duration was 8 weeks. Mice were sacrificed at 4 weeks and the end of the study (8 weeks). Centrifugation was used to separate plasma from blood. All tissue samples were kept at –80°C.
Hematoxylin and eosin stain and Masson’s trichrome stain
Mouse liver tissues were fixed in 4% paraformaldehyde for histological analysis. The tissues were embedded in paraffin wax after being dehydrated with a graded series of ethanol solutions. Serial frontal sections were cut and stained with hematoxylin and eosin to grade NASH activity, and Masson’s trichrome to stage fibrosis. Photographs were obtained using a microscope.
Hepatic malondialdehyde determination
Liver tissue was placed in a microtube containing buffer (20 mM sodium-phosphate buffer at pH approximately 3–3.2+0.5% TritonX-100) and then homogenized in an icebox using a homogenizer. All the samples were normalized by using a bicinchoninic acid protein assay. Malondialdehyde (MDA) formation in the liver was measured using a Lipid Peroxidation (MDA) Assay kit (ab233471, Abcam, Cambridge, UK).
Quantitative real time-polymerase chain reaction
Total RNA was extracted with an AccuPrep Universal RNA Extraction Kit (Bioneer Corp., Daejeon, Korea), and then transcribed to cDNA with Maxime RT PreMix Kit (iNtRON Biotechnology Inc., Seongnam, Korea), according to the manufacturer’s instructions. Real-time polymerase chain reaction (PCR) analysis was performed using Power SYBR Green PCR Master Mix (Life Technologies Ltd., Paisley, UK) on QuantStudio 6 Flex Real-Time PCR System (Life Technologies Holdings Pte Ltd., Singapore) with the primer sequences described below. The comparative cycle threshold (ΔΔCt) method was used to evaluate the mRNA expression levels of target genes, with the cycle threshold values of target genes normalized to GAPDH as an internal standard. The primer sequences were as follows: tumor necrosis factor-α (TNFα) forward primer 5´-ATGGCCTCCCTCTCATCAGT -3´ and reverse primer 5´-TTTGGTACGACGTGGGCTAC-3´; interleukin-1β (IL-1β) forward primer 5´-GAGCACCTTCTTTTCCTTCATCTT-3´ and reverse primer 5´-TCACACACCAGCAGGTTATCATC-3´; GAPDH forward primer 5´-ACTCCACTCACGGCAAATTC-3´ and reverse primer 5´-TCTCCATGGTGGTGAAGACA-3´.
Statistical analyses
Results are presented as the mean±standard error of the mean of at least three independent replicate experiments. Differences between the treatment groups were evaluated using the Student t test with Microsoft Excel for Microsoft 365 version 2109 build 16.0.14430.20292 (Microsoft Corp., Redmond, WA, USA). Statistical significance was set at P<0.05.
Go to :
RESULTS
Gemigliptin ameliorated HSC activation
As shown in Fig. 1A, treatment of LX-2 cells with succinate for 24 hours caused a significant increase in the expression of fibrogenic proteins (α-SMA, collagen type 1) when compared to that in control cells, and co-treatment with gemigliptin effectively reduced the expression of the fibrogenic proteins in a dose-dependent manner. The proliferation of LX-2 cells at 24 and 48 hours following succinate stimulation increased in a time-dependent manner compared to the control cells (Fig. 1B). However, gemigliptin (100 μM) markedly inhibited succinate-induced HSC proliferation at 48 hours (Fig. 1B). These results suggest that gemigliptin may ameliorate succinate-induced HSC activation.
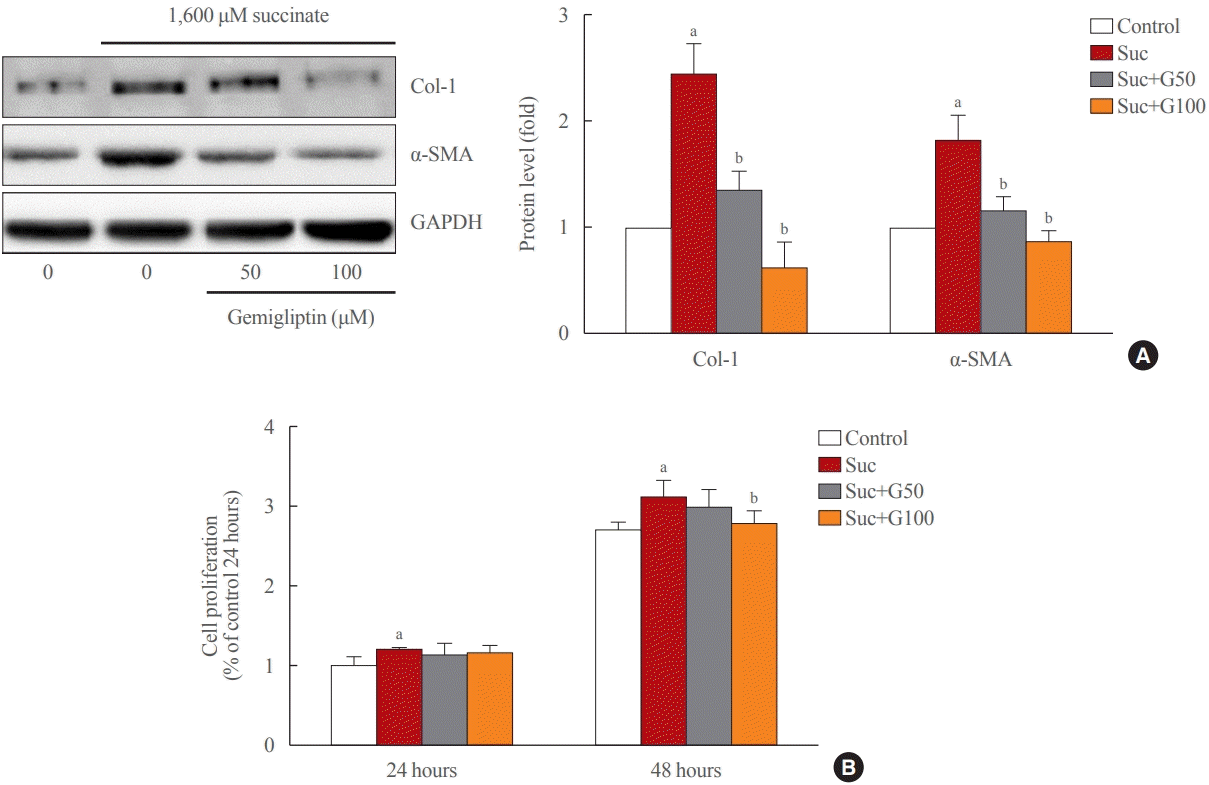 | Fig. 1.Gemigliptin ameliorated succinate-induced hepatic stellate cell activation and proliferation. (A) LX-2 cells were treated with 1,600 μM succinate and co-treated with 50 or 100 μM gemigliptin for 24 hours. Western blot analysis of α-smooth muscle actin (α-SMA) and collagen type 1 (Col-1) expression in LX-2 cells. The band intensities in Western blot images were quantified using Image Lab and plotted to the right of the representative blot images. The protein levels were normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) expression. (B) The proliferation of LX-2 cells that were treated with 1,600 μM succinate and co-treated with 50 or 100 μM gemigliptin for 24 and 48 hours. aP<0.05 vs. control; bP<0.05 vs. succinate (mean±standard error of the mean, n=3).
|
Gemigliptin inhibited the succinate-GPR91 signaling pathway in HSCs
Succinate treatment significantly increased GPR91 expression and p-ERK1/2 signaling pathway in LX-2 cells compared to that in the control (Fig. 2). Interestingly, GPR91 and p-ERK1/2 protein levels were attenuated in LX-2 cells treated with gemigliptin in the presence of succinate (Fig. 2). These data suggest that succinate leads to HSCs activation by directly activating f GPR91 and phosphorylating ERK1/2, and gemigliptin may inhibit HSC activation by inhibiting the succinate-GPR91 pathway.
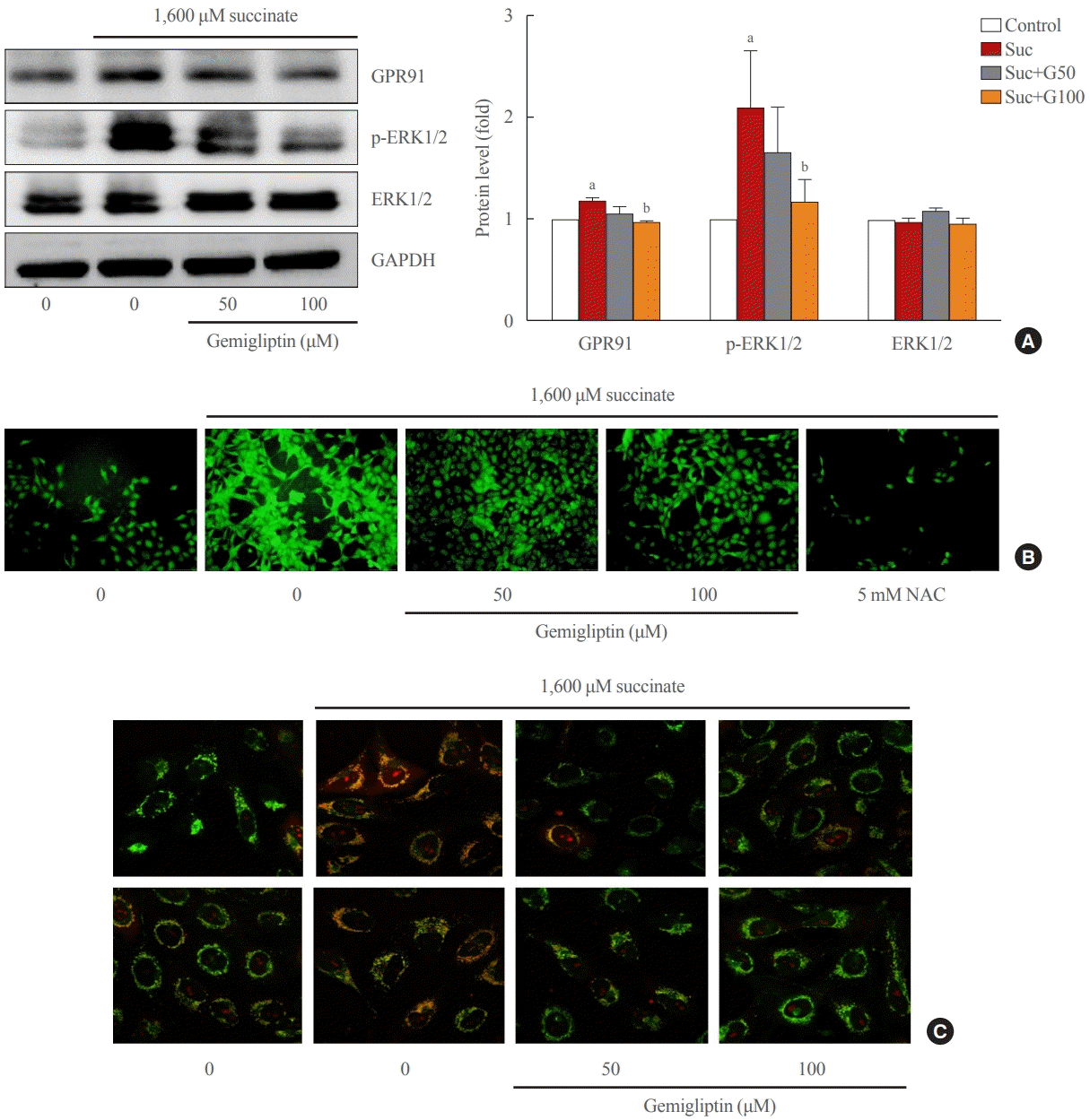 | Fig. 2.Gemigliptin inhibited the succinate-G-protein coupled receptor 91 (GPR91) signaling pathway and reactive oxygen species (ROS) production in hepatic stellate cells. LX-2 cells were treated with 1,600 μM succinate and co-treated with 50 or 100 μM gemigliptin for 24 hours. (A) Western blot analysis of GPR91, phospho-extracellular signal-regulated kinase 1/2 (p-ERK1/2), and total ERK1/2 expression in LX-2 cells. The band intensities in Western blot images were quantified using Image Lab and plotted to the right of the representative blot images. The protein levels were normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) expression. (B) Determination of cellular ROS by the dichlorofluorescin diacetate (DCFDA) assay, fluorescence microscopic images of treated cells. (C) Mitochondrial localization of ROS in LX-2 cells in various treatment groups imaged using confocal scanning microscopy (green: mitochondria; red: MitoSOX). NAC, N-acetyl-L-cysteine. aP<0.05 vs. control; bP<0.05 vs. succinate (mean±standard error of the mean, n=3).
|
Gemigliptin alleviated succinate-induced mitochondrial dysfunction
In a stressful environment, the HSCs were activated; they not only produced ECM proteins, but also generated ROS [28]. Fig. 2B indicated that succinate markedly increased intracellular ROS, and gemigliptin protected HSCs by reducing intracellular ROS in LX-2 cells. Furthermore, we observed that succinate also induced mitochondrial ROS generation, which was attenuated by treatment with gemigliptin for 1 hour (Fig. 2C).
In normal HSCs, the mitochondria presented long tubular formations, whereas succinate stimulation caused mitochondrial fragmentation, a morphological change referred to as mitochondrial fission (Fig. 3A). Gemigliptin (100 μM) alleviated succinate-induced mitochondrial fission, demonstrating its protective effect on mitochondrial integrity (Fig. 3A). Succinate increased DRP1 phosphorylation (Ser616), a marker of mitochondrial fission, which was reversed by treatment of LX-2 cells with gemigliptin for 1 hour (Fig. 3B). Furthermore, succinate treatment increased the phosphorylation of MFF, which was decreased by gemigliptin treatment (Fig. 3B). These results demonstrate the protective effect of gemigliptin on mitochondrial integrity and ROS production during HSC activation.
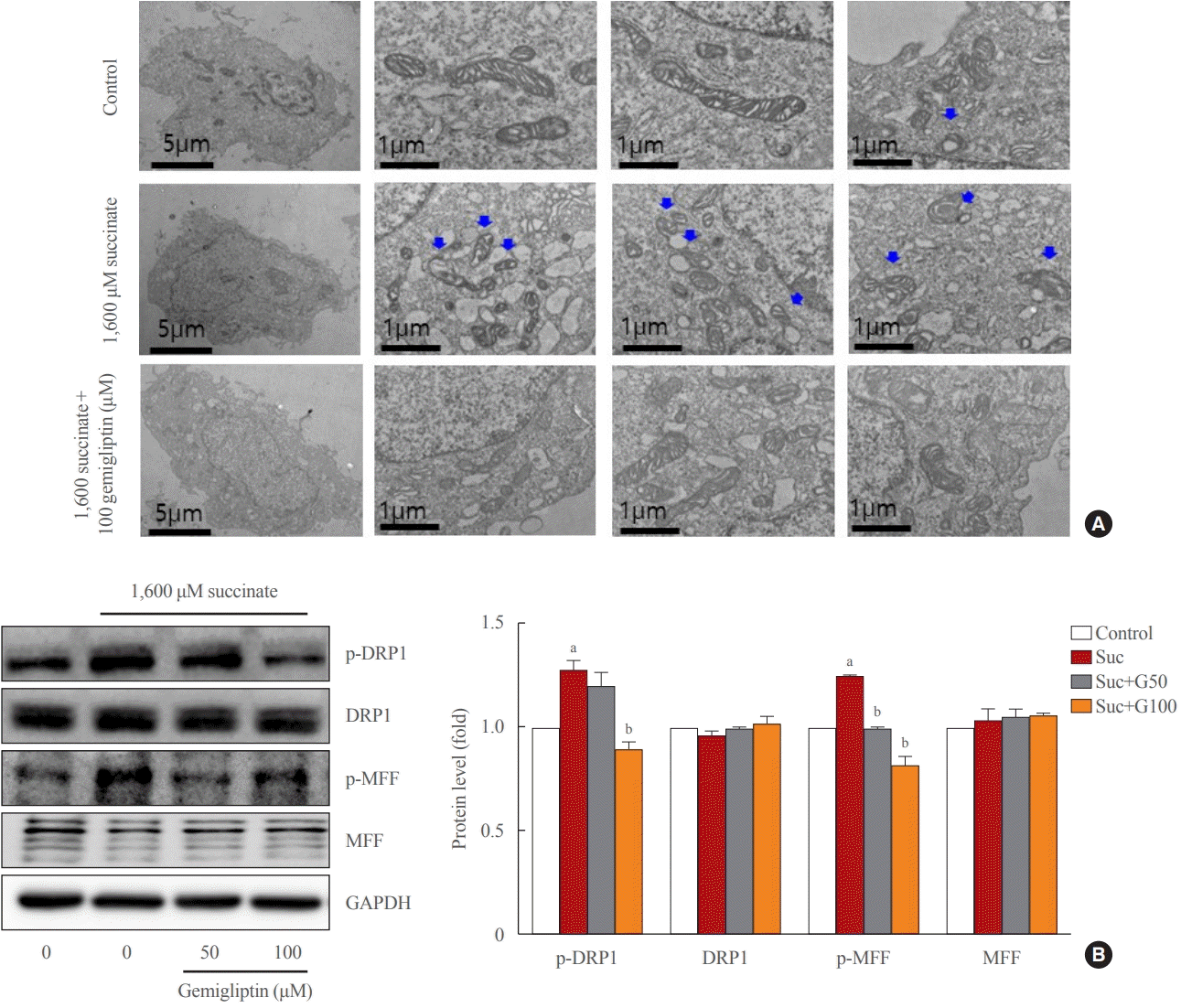 | Fig. 3.Gemigliptin alleviated succinate-induced mitochondrial fragmentation. LX-2 cells were treated with 1,600 μM succinate and cotreated with 50 or 100 μM gemigliptin for 1 hour. (A) Mitochondrial fission was detected using transmission electron microscopy. The first left column shows a single LX-2 cell in the control and treatment groups (scale bar, 2 μm). The two panels in the right column are high-magnification images (scale bar, 1 μm). Blue arrows indicate mitochondria. (B) Western blot analysis of phospho-dynamin-related protein 1 (p-DRP1), phospho-mitochondrial fission factor (p-MFF), total DRP1, and total MFF expression in LX-2 cells. The band intensities in Western blot images were quantified using Image Lab and plotted to the right of the representative blot images. The protein levels were normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH) expression. aP<0.05 vs. control; bP<0.05 vs. succinate (mean±standard error of the mean, n=3).
|
Gemigliptin attenuated liver inflammation and fibrosis
At 4 weeks of the HFHC diet, significantly increased steatosis was observed in the HFHC diet group, and gemigliptin co-treatment attenuated the steatosis. After 8 weeks of the HFHC diet, there was prominent fibrosis, and the gemigliptin co-treatment group showed less fibrosis than the HFHC diet group (Fig. 4A, B). Western blot results of the 8-week treated mice showed that the HFHC diet enhanced the hepatic expression of collagen type 1 and α-SMA and gemigliptin treatment suppressed α-SMA and collagen type 1 protein levels (Fig. 4C). In addition, the effect of gemigliptin on HFHC-induced hepatic inflammation, as measured by mRNA expression levels of TNFα and IL-1β, was attenuated after 8 weeks (Fig. 4D).
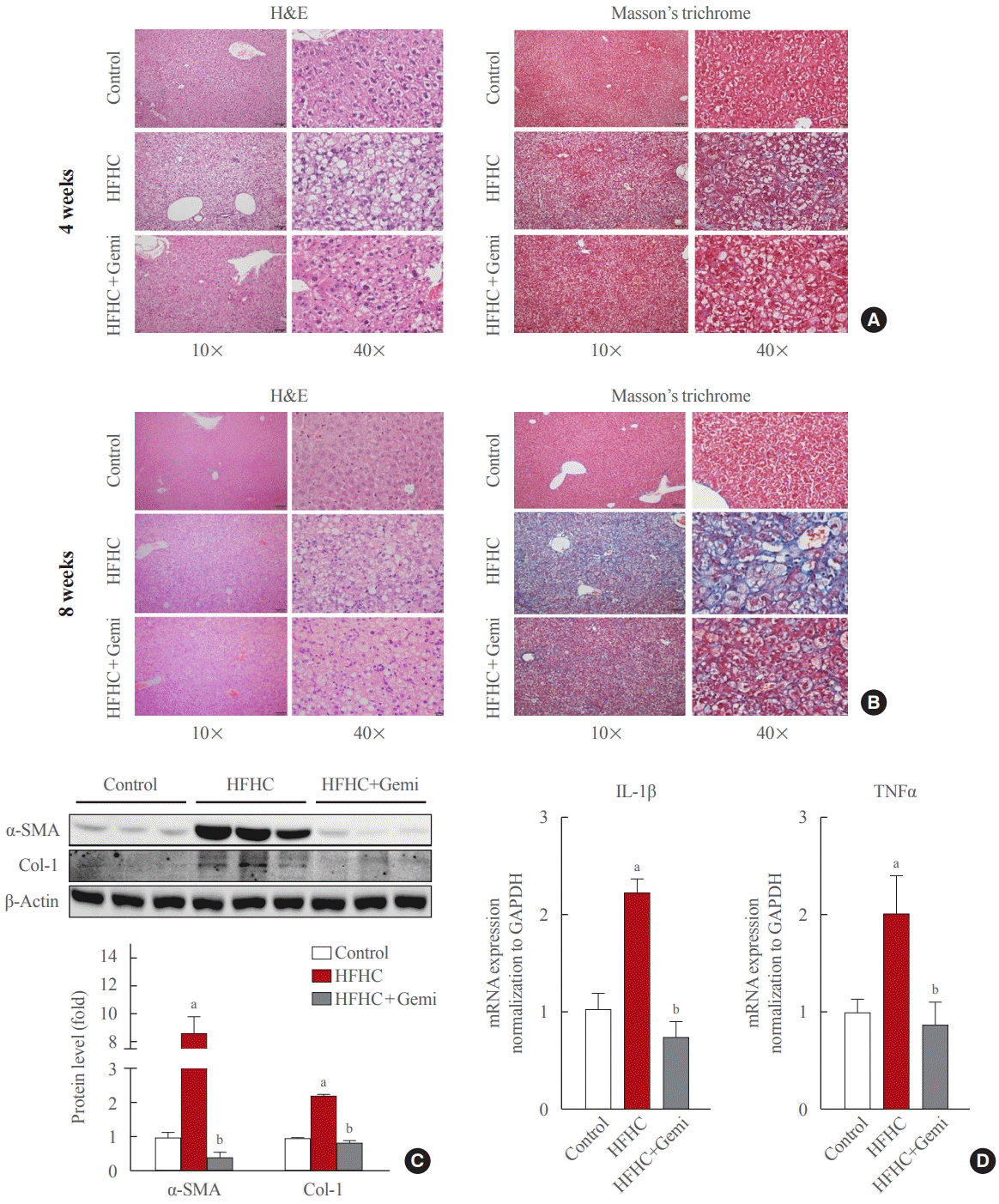 | Fig. 4.Gemigliptin attenuated liver fibrosis and inflammation in a high-fat, high-cholesterol diet mouse model. (A) H&E (left panels) and Masson’s trichrome staining (right panels) were performed to evaluate steatosis and liver fibrosis, at 4 weeks of feeding, respectively. The magnification of the image is shown. (B) H&E (left panels) and Masson’s trichrome staining (right panels) were performed to evaluate steatosis and liver fibrosis after 8 weeks of feeding, respectively. The magnification of the image is shown. (C) Western blot analysis of α-smooth muscle actin (α-SMA), and collagen type 1 (Col-1) expression in the liver from control, high-fat, high-cholesterol (HFHC) dietfed, and HFHC diet+gemigliptin-treated mice. Representative blots are shown above of the plots of the corresponding band intensities. (D) Interleukin-1β (IL-1β) and tumor necrosis factor-α (TNFα) mRNA expression levels in liver tissue were measured in the liver from control, HFHC diet-fed, and HFHC diet+gemigliptin-treated mice. aP<0.05 vs. control group; bP<0.05 vs. HFHC diet group (mean±standard error of the mean; control group [n=5], HFHC group [n=11], HFHC+gemigliptin group [n=11]).
|
Gemigliptin protected the liver from oxidative stress and mitochondrial fission
Succinate concentrations in the blood are clearly elevated in inflammatory-related health conditions, including obesity and type 2 diabetes [29-32]. Therefore, we tested the level of succinate in the blood of 8-week treated mice, and our data showed that gemigliptin administration also markedly reduced succinate levels in the serum of the HFHC diet group, which showed significantly higher succinate levels than the control diet group (Fig. 5A). Next, we measured the formation of MDA, a marker of oxidative stress, in liver tissue of 8-week treated mice. The MDA level was higher in the HFHC group than in the control group, and gemigliptin alleviated MDA levels in the co-treatment group (Fig. 5B).
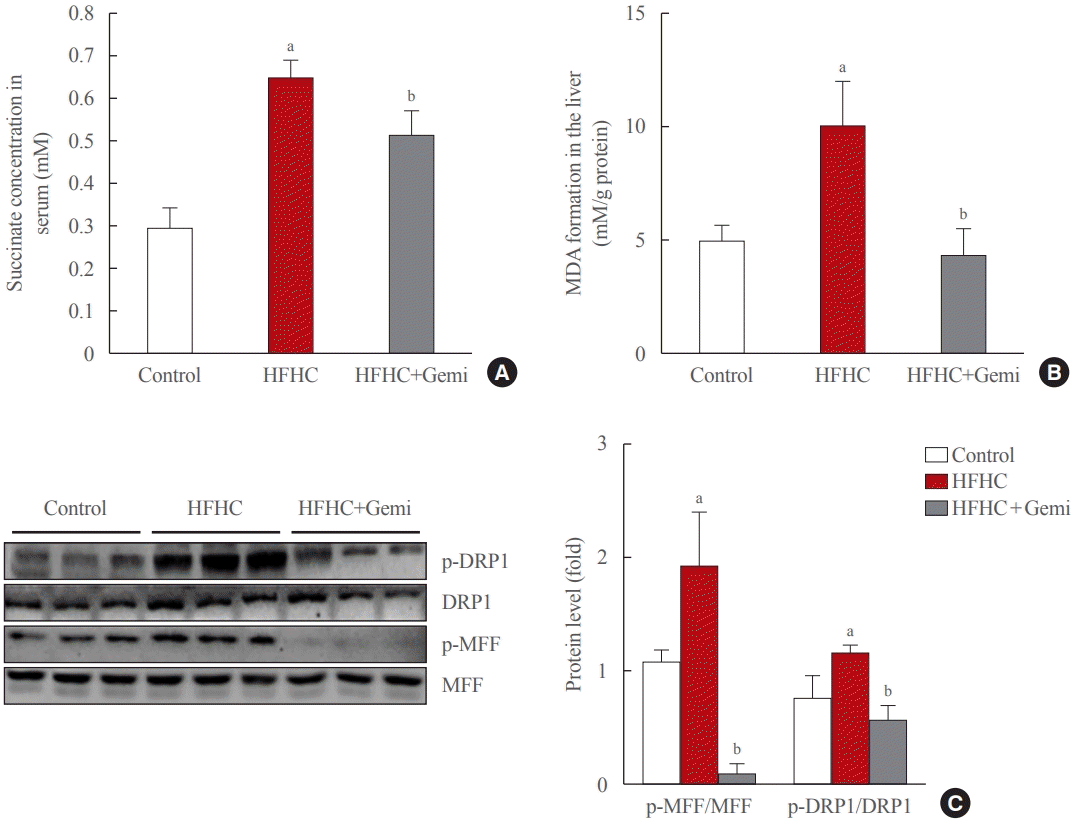 | Fig. 5.Gemigliptin protected the liver from oxidative stress and mitochondrial fission. (A) Succinate concentration in mouse serum samples collected from the indicated groups. (B) malondialdehyde (MDA) formation was measured in the liver from control, high-fat high-cholesterol (HFHC) diet-fed, and HFHC diet+gemigliptin-treated mice. (C) Western blot analysis of phospho-dynamin-related protein 1 (p- DRP1), phospho-mitochondrial fission factor (p-MFF), total DRP1, and total MFF expression in the liver from control, HFHC diet-fed, and HFHC diet+gemigliptin-treated mice. Representative blots are shown to the left of the plots of the corresponding band intensities. aP<0.05 vs. control group; bP<0.05 vs. HFHC diet group (mean±standard error of the mean; control group [n=5], HFHC group [n=11], HFHC+ gemigliptin group [n=11]).
|
Western blot results showed that the HFHC diet enhanced the hepatic expression of mitochondrial fission markers (p-DRP1 and p-MFF) and gemigliptin decreased p-DRP1 and p-MFF proteins in the liver after 8 weeks of feeding (Fig. 5C). Thus, our results suggest that the protective effects of gemigliptin on HFHC diet-induced mouse liver fibrosis may involve altered mitochondrial dysfunction, such as mitochondrial fission.
Go to :
DISCUSSION
In the present study, we showed that succinate levels were higher in the serum of HFHC-induced mice and that succinate acted as a signaling molecule to mediate mitochondrial dysfunction and HSC activation via the GPR91 signaling pathway, thus highlighting the pathological impact of succinate in the development and progression of liver fibrosis. Gemigliptin protected the morphological and functional integrity of mitochondria from the succinate insult, thereby attenuating HSC activation in vitro and reducing liver fibrosis in an in vivo HFHC-induced mouse model.
Although succinate is a citric acid cycle intermediate that is generally synthesized in the mitochondria, it can be released into the extracellular space due to metabolic disturbances and serves as a signaling molecule that regulates cellular responses, including hypoxia, inflammation, oxidative stress, and tumorigenesis [33,34]. High levels of succinate have been detected in the plasma, urine, and cerebral white matter of patients with metabolic diseases [35,36]. In line with these reports, we found elevated succinate levels in the serum of a murine NASH model, suggesting the possibility that released succinate may influence cellular responses in an endocrine manner. Gemigliptin decreased the levels of succinate in circulation in mice treated with both HFHC and gemigliptin. These results suggest that gemigliptin may improve mitochondrial dysfunction by reducing serum succinate levels.
Mitochondria are important organelles for biosynthetic processes, regulation of stress responses, and cell energy metabolism [37]. Mitochondria act as both a source and a target of intracellular ROS and are involved in the progression of oxidative stress-mediated hepatic fibrosis [38]. In hepatic fibrosis, the activation of HSCs is an important and highly dynamic process involving the synthesis of large amounts of ECM. In vivo, we found that the increase in fibrogenesis markers and inflammatory cytokines may have been correlated with the increase in phosphorylation of DRP1 and MFF, and intracellular ROS in the NASH model. In addition, gemigliptin attenuated liver fibrosis in the treatment group, accompanied by the suppression of mitochondrial fission markers. The succinate insult resulted in increased phosphorylation of DRP-1 (Ser616) and MFF in LX-2 cells, and reduced mitochondrial integrity by inducing mitochondrial fragmentation in HSCs at an early stage. We observed that succinate-induced mitochondrial fission and increased mitochondrial ROS production in LX-2 cells. Interestingly, gemigliptin attenuated mitochondrial fission by inactivating DRP1 and MFF phosphorylation. This effect may contribute to the suppression of mitochondrial ROS production in the HSCs.
Mitochondrial fission is a highly regulated process that alters the metabolism, proliferation, and apoptosis of cells. MFF has been proposed to act as a DRP1 adaptor to mediate mid-zone fission for mitochondrial biogenesis during cell proliferation [39]. In cardiomyocytes, succinate-mediated GPR91 activation increases ERK1/2 phosphorylation and promotes ERK1/2 translocation to the mitochondria, where it phosphorylates MFF to facilitate DRP1 localization to the mitochondria, thereby contributing to mitochondrial fission [40]. In the liver, HSCs exhibit increased GPR91 expression following succinate stimulation, leading to their activation [8]. GPR91 receptor activation elicits the phosphorylation of downstream signaling molecules, such as ERK, which enhances the expression of nuclear transcription factors and production of α-SMA, thereby stimulating the activation and proliferation of HSCs [8,9]. Taken together, our results show that succinate-induced mitochondrial fission at early time points in HSCs may be associated with the upregulation of GPR91 and increased phosphorylation of the ERK1/2 downstream pathway, which ultimately enhances the proliferation of HSCs. Furthermore, the beneficial effect of gemigliptin on mitochondrial dysfunction also protects against HSC activation by downregulating the succinate-GPR91 signaling pathway.
Overall, the present study shows that succinate acts as a signaling molecule that mediates mitochondrial dysfunction in parallel with its activation in HSCs. Gemigliptin preserves the morphological integrity of mitochondria and attenuates mitochondrial ROS production by inhibiting DRP1 translocation and MFF phosphorylation. Our work not only provides new insights into the protective effects of gemigliptin against liver fibrosis, but also suggests that suppression of the succinate-GPR91 pathway and modulation of DRP1 phosphorylation represent potential therapeutic strategies for the treatment of liver fibrosis.
Go to :
 XML Download
XML Download