

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
Diabetic nephropathy (DN) is a serious microvascular complication that is associated with high mortality and disability [1]. There are reports linking inflammation to kidney damage in DN [2,3]. However, the mechanism through with inflammation affects kidney damage in DN is still unclear. Efforts must be taken to identify this mechanism as it may play a crucial role in developing effective treatments for DN. Pyroptosis is a form of programmed cell death—induced in response to inflammatory responses—that has been reported to be involved in the pathogenesis of DN [4,5]. Further, pyroptosis has been reported to be accompanied by the release of pro-inflammatory factors, including interleukin-1β (IL-1β) and interleukin-18 (IL-18) [6,7]. Additionally, studies have confirmed that gasdermin D (GSDMD) is the executor of pyroptosis, and that pyroptosis is initiated by the cleavage of gasdermin D by activated caspase 1 (i.e., cleaved caspase-1), which results in the formation of N- and C-terminal domains of GSDMD (GSDMD-N and GSDMD-C, respectively). GSDMD-N then translocates to the plasma membrane and induces pore formation, thereby inducing pyroptosis. Subsequently, the cell membrane ruptures and the secretion of IL-1β and IL-18 increases [8,9]. Pyroptosis also damages high glucose (HG)-treated renal tubular epithelial cells and renal glomerular endothelial cells [10,11]. However, the mechanism underlying pyroptosis in the background of DN is not entirely understood. The nuclear transcription factor-κB (NF-κB) signaling pathway plays a vital role in the regulation of inflammatory responses. Studies have found that the activation of NF-κB is involved in renal inflammation and fibrosis during the progression of DN [12,13]. Furthermore, recent studies have shown that the NLR family pyrin domain-containing 3 (NLRP3) inflammasome is directly involved in renal inflammation that leads to the progression of diabetic glomerular damage [14,15]. Studies have also shown that NF-κB and NLRP3 inflammasomes are involved in pyroptosis of cells in several diseases [16-18], including renal tubular pyroptosis in DN [10,19]. Hence, regulating the NF-κB/NLRP3 inflammasome signaling pathway could affect the pathogenesis of DN.
Discoid domain receptor 1 (DDR1), a type of receptor tyrosine kinase, is expressed extensively in various tissues, such as the brain, lungs, kidneys, and placenta [20]. DDR1 is involved in a series of physiological and pathological processes, including migration, proliferation, survival, differentiation, and extracellular matrix remodeling [21]. Recent studies have shown that DDR1 is significantly upregulated in the aortic tissues of rats with diabetes and in HG-treated human umbilical vein endothelial cells (HUVECs) [22]. Studies have also confirmed that DDR1 knockdown attenuates vascular endothelial damage [22]. In addition, the upregulation of DDR1 was observed in mice with crescentic glomerulonephritis. DDR1 knockdown inhibits the expression of inflammatory cytokines, such as macrophage chemoattractant protein-1, IL-1β, and tumor necrosis factor-α via the NF-κB pathway to alleviate kidney injury [23]. Therefore, DDR1 is considered to be associated with the inflammatory pathways. Our previous study has revealed that DDR1 is involved in apoptosis and the development of renal fibrosis during the progression of DN [24]. However, the specific effects of DDR1 on DN-associated inflammation and the NF-κB/NLRP3 pathway are still unclear, especially with respect to its effects on renal tubular epithelial cell pyroptosis. Hence, the purpose of this study is to explore the role of DDR1 in DN, both in vivo and in vitro, and to further explore the potential molecular mechanisms.
Go to : 
METHODS
Reagents
Streptozotocin (STZ) and D-glucose were purchased from Sigma-Aldrich (Taufkirchen, Germany). Hematoxylin-eosin (H&E) staining kits (Roche, Indianapolis, IN, USA), Propidium iodide (PI), HRP-labeled anti-rabbit secondary antibodies, HRP-labeled anti-mouse secondary antibodies and anti-β-actin antibodies were purchased from Beyotime Biotechnology (Shanghai, China). Enzyme linked immunosorbent assay (ELISA) kits for IL-1β and IL-18 were procured from Zhongshan Biotechnology (Beijing, China). Antibodies against GSDMD, caspase-1, and NLRP3 were purchased from Proteintech (Wuhan, China). Further, antibodies against IL-1β, IL-18, and DDR1 were purchased from Cell Signaling Technology (Danvers, MA, USA). Antibodies against NF-κB and phosphorylated NF-κB (pNF-κB) were purchased from Santa Cruz Biotechnology (Dallas, TX, USA). siDDR1, Real-time reverse transcription-quantitative polymerase chain reaction (RT-qPCR) kits, and the Lipofectamine 3000 were purchased from RiboBio (Guangzhou, China).
Establishment of a rat model of DN
The animal experimental protocol has been approved by the Experimental Animal Ethics Committee (Changsha, China), in line with internationally recognized ethical standards. Male specific pathogen-free rats of the Sprague–Dawley strain (body weight, 200 ± 20 g) were obtained from the Hunan SJA Laboratory Animal Co., Ltd (Changsha, China), and were housed room temperature (25 ± 1°C) with free access to food and water. The rats were randomly divided into two groups, i.e., control and DN. The rats in these two groups were fed for 12 weeks under the same conditions. The DN rats were administered a single intraperitoneal injection of STZ (60 mg/kg) [25]. To confirm the successful establishment of the DN model, glucose levels were measured in the blood collected from the tail vein of the DN group rats. The blood glucose was higher than 16.7 mM for 3 consecutive days, indicating the establishment of a successful DN model. The control rats were intraperitoneally injected with an equal volume of citrate buffer, pH 4.4.
H&E staining
Kidney tissues were fixed with 4% paraformaldehyde, embedded in paraffin, and sectioned (5 μm). The sections were stained with H&E, and the morphological changes in the glomeruli and tubular interstitium were observed using a fluorescence microscope (Olympus, Nanjing, China).
Measurement of related index parameters
Albumin excretion, plasma creatinine level, and plasma cystatin-C level were measured using kits (W Systems, Minneapolis, MN, USA), as per the manufacturer's instructions. The glomerular volume, fractional mesangial area, and tuft area were analyzed using Image-Pro Plus (Media 149 Cybernetics, Silver Springs, MD, USA).
Cell culture and treatment
Human renal tubular (HK-2) cells—cultured for at most 10 generations—were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). The HK-2 cells were cultured in a Dulbecco's modified eaglemedium (DMEM; Gibco Life Technologies, Grand Island, NY, USA) containing 5.5 mM glucose, supplemented with 10% fetal bovine serum (Gibco Life Technologies) and 1% penicillin/streptomycin (Beyotime Institute of Biotechnology) at 37°C in an atmosphere 5% CO2. In the experiments, the control cells were cultured in the presence of 5.5 mM D-glucose for 48 h, whereas HG cells were exposed to 33 mM D-glucose for 48 h. Subsequently, for DDR1 knockdown experiments, HK-2 cells were divided into three groups, i.e., HG (exposed to 33 mM D-glucose for 48 h), DDR1-negative control (exposed to 33 mM D-glucose for 48 h after transfection with siControl for 24 h), and DDR1 siRNA (exposed to 33 mM D-glucose for 48 h after transfection with siDDR1 for 24 h). Lipofectamine 3000 was used to transfect the pcDNA3.1-NLRP3 overexpression vector (OVE-NLRP3) and the pcDNA3.1-NC vector (OVE-NC) into DDR1 siRNA cells. The vectors were designed and synthesized by Shanghai Gene Pharmaceutical Co., Ltd (Shanghai, China).
PI staining
HK-2 cells were stained with 4',6-diamidino-2-phenylindole (DAPI) (Beyotime Biotechnology) at 25°C in the dark for 10 min, followed by PI staining (5 mg/ml) at 25°C in the dark for 10 min. Fluorescence microscopy was used to verify the high membrane permeability in HK-2 cells.
ELISA
The concentrations of IL-1β and IL-18 in the plasma of the rats and the HK-2 cells culture supernatants were determined using ELISA kits, as per the manufacturers’ instruction.
Western blotting
The rat kidney tissues and HK-2 cells were independently lysed in RIPA buffer supplemented with 1% phenylmethanesulfonyl fluoride (Beyotime Biotechnology). The proteins extracted from these samples were quantified using commercial bicinchoninic acid protein assay kits (Beyotime Biotechnology). Forty micrograms of individual samples were loaded into each lane of sodium dodecyl sulfate-polyacrylamide gels and resolved by electrophoresis. The separated proteins were transferred onto polyvinylidene fluoride (PVDF) membranes (Sigma-Aldrich). Following which, the membranes were blocked for 2 h using 5% non-fat milk at room temperature (25 ± 1°C) and incubated overnight with primary antibodies (anti-caspase-1 [1:500 dilution], anti-GSDMD [1:1,000], anti-IL-1β [1:1,000], anti-IL-18 [1:1,000], anti-DDR1 [1:1,000], anti-NF-κB [1:1,000], anti-p-NF-κB [1:500], anti-NLRP3 [1:1,000], and anti-β-actin [1:1,000]) at 4°C. Subsequently, the PVDF membranes were washed with TBS-Tween and incubated with HRP-labeled anti-rabbit (1:5,000) or anti-mouse (1:5,000) secondary antibodies for 2 h at room temperature on a shaker. Chemiluminescence was visualized using ECL kits (Beyotime Biotechnology). Protein bands on the membrane were imaged on ChemiDoc XRS system and the corresponding intensities were evaluated using ImageJ (Bio-Rad, Philadelphia, PA, USA).
RT-qPCR
Total RNA was extracted from HK-2 cells using the Trizol reagent (TaKaRa, Beijing, China), in accordance with the instructions provided by the manufacturer, and the concentration and purity of this RNA were determined using the absorbance at 260 nm. This RNA was then reverse transcribed into cDNA using 7500 Sequence Detector system (Applied Biosystems, Foster City, CA, USA), in accordance with the manufacturer’s instructions. For RT-qPCR, 2 μl of cDNA was used as template, and a SYBR Green‒based methodology was used employed. The following conditions were used for RT-qPCR: 95°C for 10 min; 40 cycles of 95°C for 15 sec, 60°C for 1 min, and 95°C for 15 sec. Quantitative analysis was performed using the 2–ΔΔCt method. The primers used for RT-qPCR were synthesized by Sangon Biotech Co., Ltd (Shanghai, China), andtheir sequences were listed in Table 1.
Table 1
Primer sequence of the gene
![]()
Statistical analysis
SPSS 20.0 (IBM Co., Armonk, NY, USA) was used to analyze the experimental data. The data were represented as mean ± standard error of the mean (SEM). The differences between two groups were analyzed using the Student's t-test, and one-way analysis of variance was used to analyze the differences among multiple groups.
Go to :
RESULTS
Pyroptosis in DN rat kidney tissues and HK-2 cells
In DN rats, we analyzed the effect of hyperglycemia on pyroptosis in the kidney tissues. Analysis of renal pathology in DN rats revealed glomerular and tubulo-interstitial alterations (Fig. 1A). Additionally, the DN rats exhibited high albumin/creatinine ratio, plasma cystatin-C levels, glomerular volume, fractional mesangial area, and tuft area compared to control rats (all p < 0.01) (Fig. 1B–F). Western blotting revealed that the expression of cleaved caspase-1, GSDMD-N, IL-1β, and IL-18 was increased. Further, ELISA revealed that the concentrations of IL-1β and IL-18 were significantly higher in the DN rats than in the control rats (all p < 0.01) (Fig. 2A–D). These findings indicated the occurrence of pyroptosis in the renal tissues of the DN rats. To further explore the role of pyroptosis in DN, we treated HK-2 cells with HG to mimic DN conditions. Consistent with the in vivo findings, pyroptosis was observed in case of HG-treated HK-2 cells, as indicated by the upregulation of cleaved caspase-1, GSDMD-N, IL-1β, and IL-18. This is important because, rupture of the cell membrane, accompanied by the release of several molecules including IL-1β and IL-18, is a typical feature associated with pyroptosis (all p < 0.01) (Fig. 2E, F). ELISA revealed that the concentrations of IL-1β and IL-18 were significantly higher in the HG group compared to those in the control group (all p < 0.01) (Fig. 2G, H). These results confirmed that HG induced the pyroptosis of HK-2 cells.
 | Fig. 1Changes in the kidney function and morphology in diabetic rats.A rat model wherein streptozotocin (STZ) was used to induce diabetic nephropathy (DN). (A) H&E staining to observe the morphological characteristics of the kidney (scale bar 50 μm, ×400; scale bar 200 μm, ×100) magnifications, (B) Albumin/creatinine ratio. (C) Plasma cystatin-C levels. (D) Glomerular volume. (E) Mesangial area. (F) Tuft area. Data are presented as mean ± SEM from 6 rats. Compared to control, **p < 0.01.
|
 | Fig. 2In vivo and in vitro expression of pyroptosis-related proteins.(A) Bands corresponding to the expression of pyroptosis-related proteins (caspase-1 p45, cleaved caspase-1, interleukin [IL]-1β, IL-18, gasdermin D [GSDMD], and GSDMD-N) in the kidneys of diabetic nephropathy (DN) rats. (B) Quantification of the bands observed in (A). (C) Concentration of IL-1β in rat plasma. (D) Concentration of IL-18 in rat plasma. (E) Bands corresponding to the expression of pyroptosis-related proteins (caspase-1 p45, cleaved caspase-1, IL-1β, IL-18, GSDMD, and GSDMD-N) in high glucose (HG)-treated HK-2 cells. (F) Quantification of the bands observed in (E). (G) Concentration of IL-1β in HK-2 cell supernatant. (H) Concentration of IL-18 in HK-2 cell supernatant. Data are presented as mean ± SEM from 6 rats and 3 replicated cell experiments. Compared to control, *p < 0.05, **p < 0.01.
|
In vivo and in vitro investigation of the role of DDR1 in pyroptosis
As DDR1—an inflammatory protein—is known to play an important role in numerous pathophysiological processes, we investigated the role of DDR1 in pyroptosis. Higher expression of DDR1 was observed in the DN rat kidney tissues and HG-treated HK-2 cells (all p < 0.01) (Fig. 3), suggesting that DDR1 may be an important regulator of kidney damage in DN.
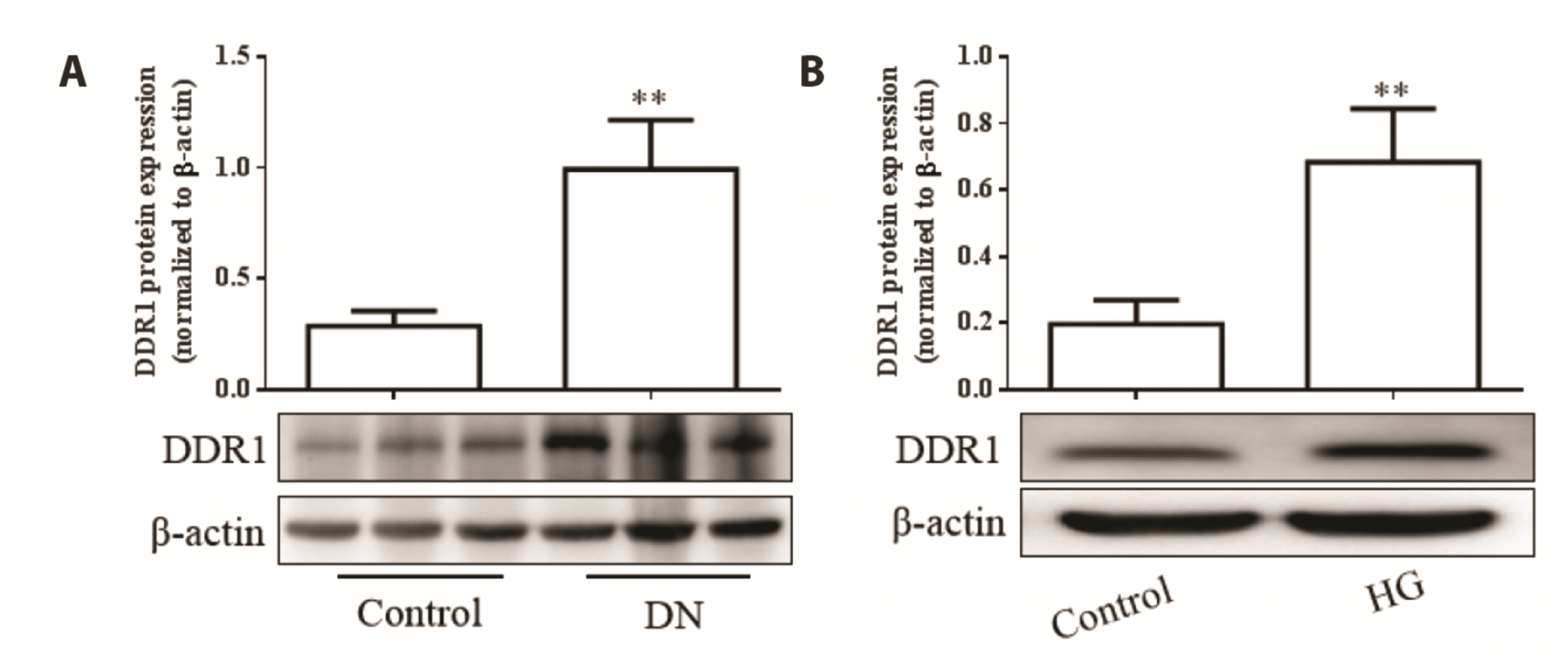 | Fig. 3In vivo and in vitro expression of discoid domain receptor 1 (DDR1).(A) DDR1 expression in the kidneys of diabetic nephropathy (DN) rats. (B) DDR1 expression in high glucose (HG)-treated HK-2 cells. Data are presented as mean protein in the kidneys of DN rats and HG-treated HK-2 cells. Data are presented as mean ± SEM from 6 rats and 3 replicated cell experiments. Compared to control, **p < 0.01.
|
Role of DDR1 in pyroptosis of HG-treated HK-2 cells
Based on the observation that the expression of DDR1 is upregulated in HG-treated HK-2 cells, we speculated that DDR1 might play a role in HG-induced pyroptosis of HK-2 cells as well. Validation experiments clearly showed that the expression of DDR1 was decreased in HG-treated HK-2 cells after transfection with siDDR1 (p < 0.01) (Fig. 4A, B). Subsequently, PI staining was performed to analyze the effect of DDR1 on membrane rupture during pyroptosis. The increase in the percentage of PI-positive cells in HG-treated cells was suppressed in the background of DDR1 knockdown (p < 0.01) (Fig. 4C, D). Furthermore, DDR1 knockdown significantly suppressed the HG-induced elevation in the levels of cleaved caspase-1, GSDMD-N, IL-1β, and IL-18 (p < 0.05 or p < 0.01) (Fig. 4E, F). The concentrations of IL-1β and IL-18 decreased after silencing DDR1 in the background of HG treatment (compared to HG group) (all p < 0.01) (Fig. 4G, H).
 | Fig. 4Effect of discoid domain receptor 1 (DDR1) knockdown on HK-2 cell pyroptosis.(A) Bands corresponding to the expression of DDR1 in the background of transfection with siDDR1. (B) Quantification of the band observed in (B). (C) Propidium iodide (PI) staining was used to analyze HK-2 cell death (scale bar: 200 μm). (D) Percentage HK-2 cell pyroptosis. (E) Bands corresponding to the expression of pyroptosis-related proteins (caspase-1 p45, cleaved caspase-1, interleukin [IL]-1β, IL-18, gasdermin D [GSDMD], and GSDMD-N). (F) Quantification of the bands observed in (E). (G) The concentration of IL-1β in HK-2 cell supernatant. (H) The concentration of IL-18 in HK-2 cell supernatant. Data are presented as mean ± SEM from 3 replicated cell experiments. Compared to high glucose (HG), *p < 0.05, **p < 0.01.
|
Role of DDR1 in NF-κB/NLRP3-mediated pyroptosis
Previous studies have shown that the NF-κB/NLRP3 pathway plays a role in HG-induced pyroptosis. Therefore, we investigated the role of DDR1 in HG-induced pyroptosis of HK-2 cells and assessed the expression of the NF-κB/NLRP3 pathway intermediaries using Western blotting. The expression of p-NF-κB and NLRP3 was upregulated in the DN rat kidney tissues (Fig. 5A, B) and HG-treated HK-2 cells (all p < 0.01) (Fig. 5C, D). Moreover, DDR1 knockdown effectively inhibited the NF-κB/NLRP3 pathway, as indicated by the downregulated expression of p-NF-κB and NLRP3 (p < 0.05 or p < 0.01) (Fig. 5C, D). Hence, all data consistently revealed that DDR1 knockdown resulted in inhibited pyroptosis, and indicated that DDR1 is possibly involved in regulating the NF-κB/NLRP3 pathway.
 | Fig. 5Discoid domain receptor 1 (DDR1) knockdown resulted in the downregulation of the nuclear transcription factor-κB (NF-κB)/NLR family pyrin domain-containing 3 (NLRP3) pathway intermediaries at the protein level.(A) Bands corresponding to the expression of phosphorylated NF-κB (p-NF-κB) and NF-κB in the kidneys of DN rats. (B) Bands corresponding to the expression of NLRP3 in the kidneys of diabetic nephropathy (DN) rats. (C) Bands corresponding to the expression of p-NF-κB and NF-κB in HK-2 cells. (D) Bands corresponding to the expression of NLRP3 in HK-2 cells. Data are presented as mean ± SEM from 6 rats and 3 replicated cell experiments. Compared to control, **p < 0.01. Compared to high glucose (HG), #p < 0.05, ##p < 0.01.
|
Role of NLRP3 in DDR1-induced pyroptosis
To confirm the role of NLRP3 in DDR1-induced pyroptosis, we overexpressed NLRP3 in HK-2 cells. The success of overexpression was verified by RT-qPCR. Our results showed that the expression of NF-κB, caspase-1, GSDMD, IL-1β, and IL-18 that had been suppressed in response to DDR1 knockdown was upregulated upon overexpressing NLRP3 (all p < 0.01) (Fig. 6). Taken together, our results show that DDR1 knockdown inhibited HG-induced pyroptosis of renal tubular epithelial cells by regulating the NF-κB/NLRP3 pathway, and that overexpression of NLRP3 partially reversed these effects of DDR1 knockdown.
 | Fig. 6NLR family pyrin domain-containing 3 (NLRP3) reversed the discoid domain receptor 1 (DDR1) knockdown‒mediated downregulation of the nuclear transcription factor-κB (NF-κB)/NLRP3 pathway intermediaries at the transcript level (assessed using RT-qPCR).Transcript-level expression of (A) NLRP3, (B) NF-κB, (C) caspase-1, (D) gasdermin D (GSDMD), (E) interleukin (IL)-1β, (F) IL-18. Data are presented as mean ± SEM from 3 replicated cell experiments. Compared to control **p < 0.01. Compared to high glucose (HG), ##p < 0.01. Compared to siDDR1, &&p < 0.01.
|
Go to :
DISCUSSION
DN is one of the main causes of chronic kidney failure. In recent years, the incidence of DN has been reported be associated with significantly increased risk of end-stage renal disease (60%), a condition whose pathogenicity is associated with multiple factors [26]. However, the mechanism underlying the pathogenesis of this condition has not been completely elucidated. Pyroptosis is a recently discovered form of cell death that is different from apoptosis and necrosis [27]. In the context of pyroptosis, the classical signaling pathway mediated by caspase-1 is the most widely studied [28]. HG has been reported to significantly upregulate the expression of caspase-1 in renal tubular epithelial cells and renal glomerular endothelial cells [10,11]. Moreover, the silencing of caspase-1 has been reported to effectively inhibit HG-induced pyroptosis, thereby attenuating DN-induced kidney damage [29]. Studies have also reported that in STZ-treated rats, the expression of pyroptosis-related proteins (cleaved caspase-1, GSDMD-N, IL-1β, and IL-18) was significantly upregulated in the kidney tissues [30,31]. Both in vivo and in vitro experiments have revealed that caspase-1-mediated pyroptosis plays an important role in the occurrence and progression of DN. In this study, we established a DN rat model by employing intraperitoneal injections of STZ. To verify the involvement of pyroptosis in DN pathogenesis, we assessed the expression of pyroptosis-related proteins and the concentrations of IL-1β and IL-18 in vivo and in vitro. Consistent with the findings of previous studies [10,30,31], our results revealed pyroptosis in DN rats and HG-treated renal tubular epithelial cells.
As the regulatory mechanism of pyroptosis in DN is not yet fully understood, we explored the role of the inflammatory protein DDR1 in pyroptosis in the context of DN. DDR1, a receptor tyrosine kinase, has emerged as a focus of interest in cardiovascular diseases [32,33]. In the context of endothelial senescence, the expression of DDR1 was upregulated—accompanied by a decrease in cell adhesion and migration—in diabetic rats and HG-treated endothelial cells [22]. Additionally, DDR1 silencing significantly improved endothelial function [22]. DDR1 has also been shown to accelerate renal fibrosis and renal cell apoptosis in DN rats [24]. These findings suggest that DDR1 is involved in diabetic vascular disease. We found that DDR1 expression was upregulated in vivo and in vitro, indicating kidney damage as an effect. Yan et al. [22] found that miR-199a-3p is downregulated in aortic tissue of diabetic rats and in HG-treated HUVECs, while overexpression of miR-199a-3p suppresses HG-induced increase in DDR1 expression. Studies have also confirmed that there is a distinct mutual regulatory relationship between insulin growth factor receptor 1 (IGF1R) and DDR1. When IGF1R is activated, it rapidly stimulates the PI3K/Akt signaling network, and triggers the inhibition of miR-199a-5p, thereby promoting DDR1 expression [34,35]. These findings suggest that the upregulated expression of DDR1 may be related to the inhibition of specific miRNAs and the activation of certain receptors and pathways in DN. DDR1, a pro-inflammatory protein, is involved in inflammatory reactions in multiple organs [36]. In mouse models of nephrotoxic serum‒induced glomerulonephritis, DDR1 is mainly expressed in the glomerulus and DDR1 downregulation attenuates glomerular inflammation [37]. In addition, DDR1 increases the infiltration and accumulation of macrophages and the production of inflammatory cytokines in the arterial intima [38,39]. Hence, we speculated that DDR1 could play a role in the pyroptosis of renal tubular epithelial cells. To evaluate the role of DDR1 in HG-induced pyroptosis, we investigated the expression of pyroptosis-related proteins and the concentrations of IL-1β and IL-18 in HG-treated HK-2 cells with DDR1 knockdown. Our results revealed that the knockdown of DDR1 reversed the HG-induced renal tubular epithelial cell pyroptosis and dysfunction, indicating that upregulation of DDR1 may be a key trigger in DN-associated renal tubular epithelial cell pyroptosis.
NF-κB has been identified as a key inflammatory pathway in the pathogenesis of DN-associated inflammation in both animal model and human clinical studies [40,41]. In DN rats, inhibition of NF-κB activation has been reported to downregulate the expression of inflammatory cytokines, thus preventing DN progression [42]. Activation of the NF-κB pathway is a prerequisite for the activation of the NLRP3 inflammasome, a phenomenon induced in response to the upregulated expression of NLRP3, member of the NOD-like receptor protein family that is responsible for forming protein complexes in the cytoplasm that detect various cell signals related to immunity and cell death, and regulating cell function [43-45]. NLRP3 offers a platform to activate caspase-1, cleaving it into enzymatically active caspase-1, thereby promoting the maturation of the IL-1β and IL-18 precursors and the secretion of active IL-1β and IL-18 proteins [46]. In this study, the expression of the NF-κB/NLRP3 pathway intermediaries was upregulated, both in vivo and in vitro. Furthermore, downregulation of DDR1 in the background of HG treatment inhibited the HG-induced activation of the NF-κB/NLRP3 pathway in HK-2 cells. Previous studies have also reported the presence of NF-κB binding sites in the NLRP3 inflammasome promoter, and that this binding upregulates the expression of the NLRP3 inflammasome [47]. Further, they have also shown that the downregulation of NF-κB results in the attenuation of HG-induced glomerular mesangial cell pyroptosis in rats by suppressing the activation of the NLRP3 inflammasome [47]. To further clarify that DDR1 is involved in the pyroptosis of renal tubular epithelial cells in diabetic rats through the NF-κB/NLRP3 pathway, we studied the effect of DDR1 knockdown on pyroptosis by overexpressing NLRP3 in this background. Our results revealed that DDR1 knockdown resulted in the inhibition of HG-induced pyroptosis of renal tubular epithelial cells by regulating the NF-κB/NLRP3 pathway, and the overexpression of NLRP3 partially reversed the effects of DDR1 knockdown. NF-κB has been reported to function upstream of NLRP3 [48]. However, the overexpression of NLRP3 can also lead to an increase in IL-1β and IL-18 levels, and several studies have reported that the IL-1β feedback loop could promote the activation of the NF-κB pathway [49,50]. Therefore, in our study, the overexpression of NLRP3 can be inferred to have induced the activation of the NF-κB pathway, despite NF-κB functioning upstream of NLRP3. Hence, the findings of this study indicate that DDR1 siRNA exerts its anti-pyroptotic effect by regulating the NF-κB/NLRP3 signaling pathway. Matsuyama et al. reported that DDR1-mediated activation of NF-κB is regulated by Sch and a novel signaling cascade involving TRAF6 and Act1, which markedly upregulates the expression and release of pro-inflammatory mediators IL-1β and IL-18, indicating that NF-κB is a novel target for DDR1 signaling [51].
In conclusion, the data from our study prove that DDR1 is upregulated in DN rats and in HG-treated HK-2 cells, and have established the effects of DDR1. Further, DDR1 knockdown inhibited the progression of DN by attenuating the HG-induced pyroptosis of HK-2 cells, which was partially mediated through the NF-κB/NLRP3 pathway. However, the mechanism through which DDR1 affects the activation of the NF-κB pathway could not be clarified in this study. This provides scope for future studies to elucidate the mechanism through which DDR1 affects the activation of the NF-κB pathway to develop new avenues for effective DN treatment.
Go to :
 XML Download
XML Download