

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Strain typing can be used to evaluate the relatedness of clinical isolates. Pulsed-field gel electrophoresis (PFGE), multi-locus sequence typing (MLST), and whole-genome sequencing (WGS) are well-established typing methods in microbiology laboratories [1, 2], but have important limitations. PFGE is generally labor-intensive, time-consuming, and lacks reproducibility. Although MLST has excellent reproducibility, it is expensive and limited to applicable strains [1]. WGS is also limited for routine use because of its high cost and requirement of skilled researchers [2]. Matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF MS) is a routine method for species identification; however, a dendrogram based on main spectrum projection has also been suggested as a possible strain-typing method [3-5]. Recently, IR Biotyper (Bruker Daltonics GmbH, Bremen, Germany) was introduced to distinguish strains by quantifying the absorption of infrared light by lipids, nucleic acids, proteins, carbohydrates, and lipopolysaccharides from bacterial cell walls [6]. IR Biotyper uses protein fingerprints to supplement MALDI-TOF MS [7].
A simple and easy detection method is needed in the assessment of an outbreak in a healthcare facility or in investigating the source of antibiotic resistance genes. MALDI-TOF MS or IR Biotyper are good candidates for routine use in microbiology laboratories. To recommend a routine method for evaluating the clonality of strains, we compared the discriminatory power of IR Biotyper or MALDI-TOF MS with that of WGS as the reference method. To our knowledge, there are limited studies assessing strain typing methods using the etiologic agents of healthcare-associated infections. This is the first study to evaluate the discriminatory ability of IR Biotyper using community-origin strains.
MATERIALS AND METHODS
Bacterial isolates
Eighty-three extended-spectrum β-lactamase–producing Escherichia coli (ESBL-EC) isolates were included in this study. They were isolated from poultry (N=37), the poultry farm environment (N=6), slaughtered chicken meat (N=17), a slaughterhouse worker (N=1), the slaughterhouse environment (N=7), chicken meat on sale (N=14), and the retail shop environment (N=1) from January to August 2019 during a project conducted in collaboration with the Korean Centers for Disease Control and Prevention [8]. Informed consent was obtained from the slaughterhouse worker for sample collection and analysis. This study was exempted from approval review according to the Institutional Review Board of the National Health Insurance Service Ilsan Hospital, Goyang, Korea (NHIMC 2022-04-020).
WGS
DNA from freshly subcultured isolates was extracted using GenElute Bacterial Genomic DNA Kit (Sigma-Aldrich, St. Louis, MO, USA), and 8 μg of input genomic DNA was used for WGS. The entire genomes of the ESBL-EC isolates were sequenced using a NextSeq 550 instrument (Illumina, San Diego, CA, USA). Sequences were assembled using Spades (version 3.11.1) [9] and annotated using Prokka (version 1.13.7) [10]. Core-genome MLST (cgMLST) was performed using the online tool of the Center for Genomic Epidemiology [11]. A dendrogram using cgMLST was obtained from WGS data with a single linkage, and the Euclidean similarity index was calculated in PAST (version 4.03) [12].
MALDI-TOF MS
Isolates freshly cultured at 37°C for 18–24 hours in MacConkey agar were used after ethanol-formic acid extraction. MALDI-TOF MS covered the region of 2,165–18,869 m/z. FlexControl 3.3 software was used for analysis [13]. Spectra were obtained using the Biotyper Main Spectrum (MSP) identification standard method (2,000–20,000 Da, linear positive method) and edited using Flex Analysis 3.4 software (Bruker Daltonics). The mass spectra were edited using MALDI Biotyper 2.0 software (Bruker Daltonics), including MATLAB 7.1. Principal component analysis (PCA) and hierarchical cluster analysis were performed, and a PCA dendrogram was created. The dendrogram reflects the inter-relational proximity of organisms at any distance. The distance measure was calculated based on the average Euclidean distance and linkage algorithm. Only strains with a distance of more than 2.0 in the dendrogram were included for further analysis.
IR Biotyper
Isolates were cultured on Muller–Hinton medium (Thermo Fisher Scientific, Hampshire, UK) at 37°C for 18 hours because the IR Biotyper method requires that the samples are grown under identical conditions as much as possible. The culture isolates were tested three times. Colonies were selected and 1 μL of the colony was mixed well in a suspension vial containing 70% ethanol (50 μL), followed by the addition of 50 μL distilled water and mixing for 10 minutes. Three spots (15 μL each) were loaded onto a microtiter plate. The completely dried plate was mounted on the IR Biotyper system and inspected. A dendrogram was created with the raw data to cluster the separation spectrum. The cut-off value (COV) on the dendrogram was automatically calculated using OPUS 7.5 software [14]. The distance matrix based on the average Euclidean distance and linkage algorithm reflects the separation distances of all strains.
Statistical analysis
Simpson’s diversity index (SDI) and the adjusted Rand index (aRI) were used for comparison of the analytical performance of MALDI-TOF, IR Biotyper, and WGS. These indices were calculated using the online Comparing Partitions tool (www.comparingpartitions.info) [15]. The SDI represents the probability of two randomly extracted strains from a population belonging to two types [16]; the values ranges from 0 (no diversity) to 1 (infinite diversity). The SDI was calculated separately for the results obtained from MALDI-TOF, IR Biotyper, and WGS.
The Rand index (RI) quantitatively evaluates the overall consistency between two divisions [17]. There is a high possibility that good values will emerge even when random clustering is performed. To solve this problem, the aRI was developed to subtract the expected value of the RI from the original value and readjust the expected value and variance. In the case of random clustering, the aRI has a high probability of zero. Concordance and aRI of MALDI-TOF or IR Biotyper were calculated and compared with those of WGS.
RESULTS
Dendrogram
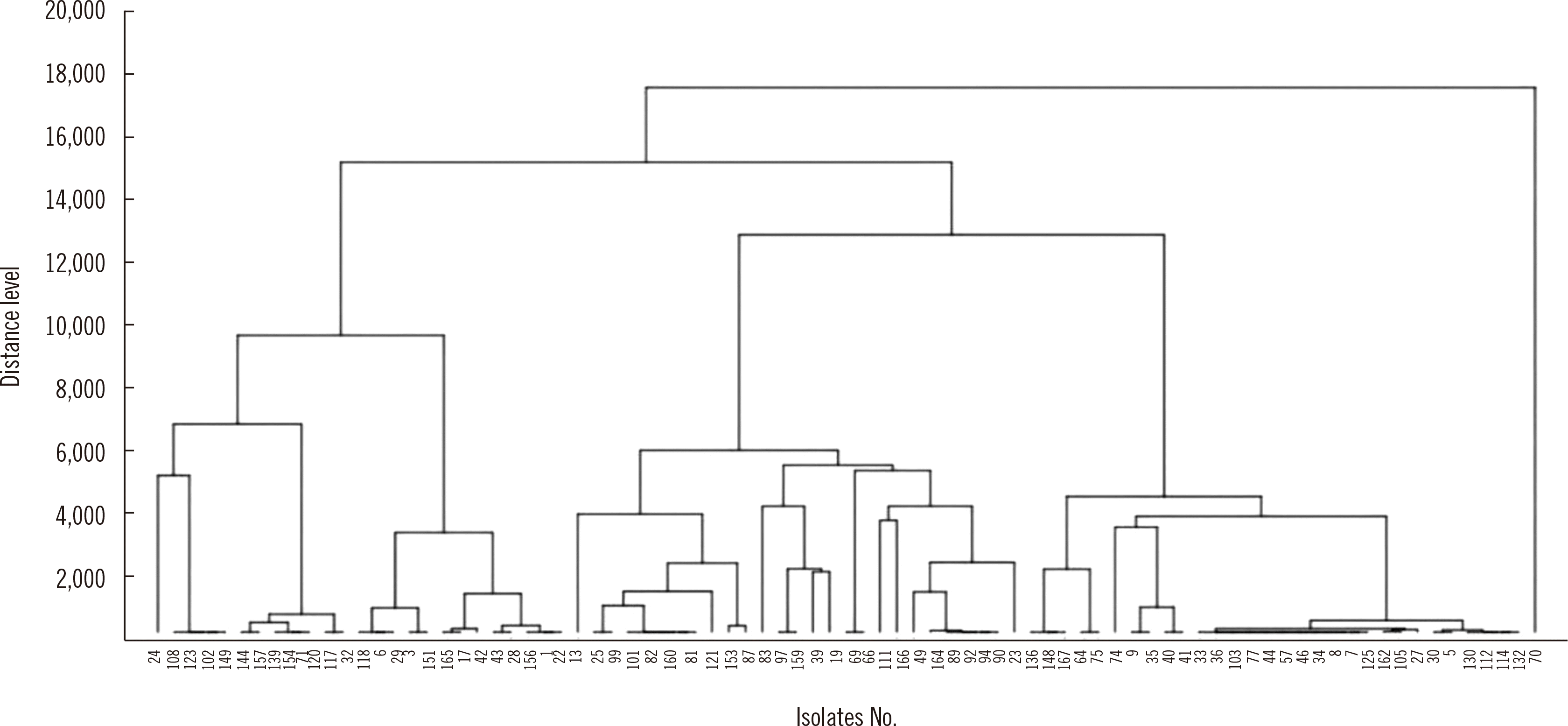
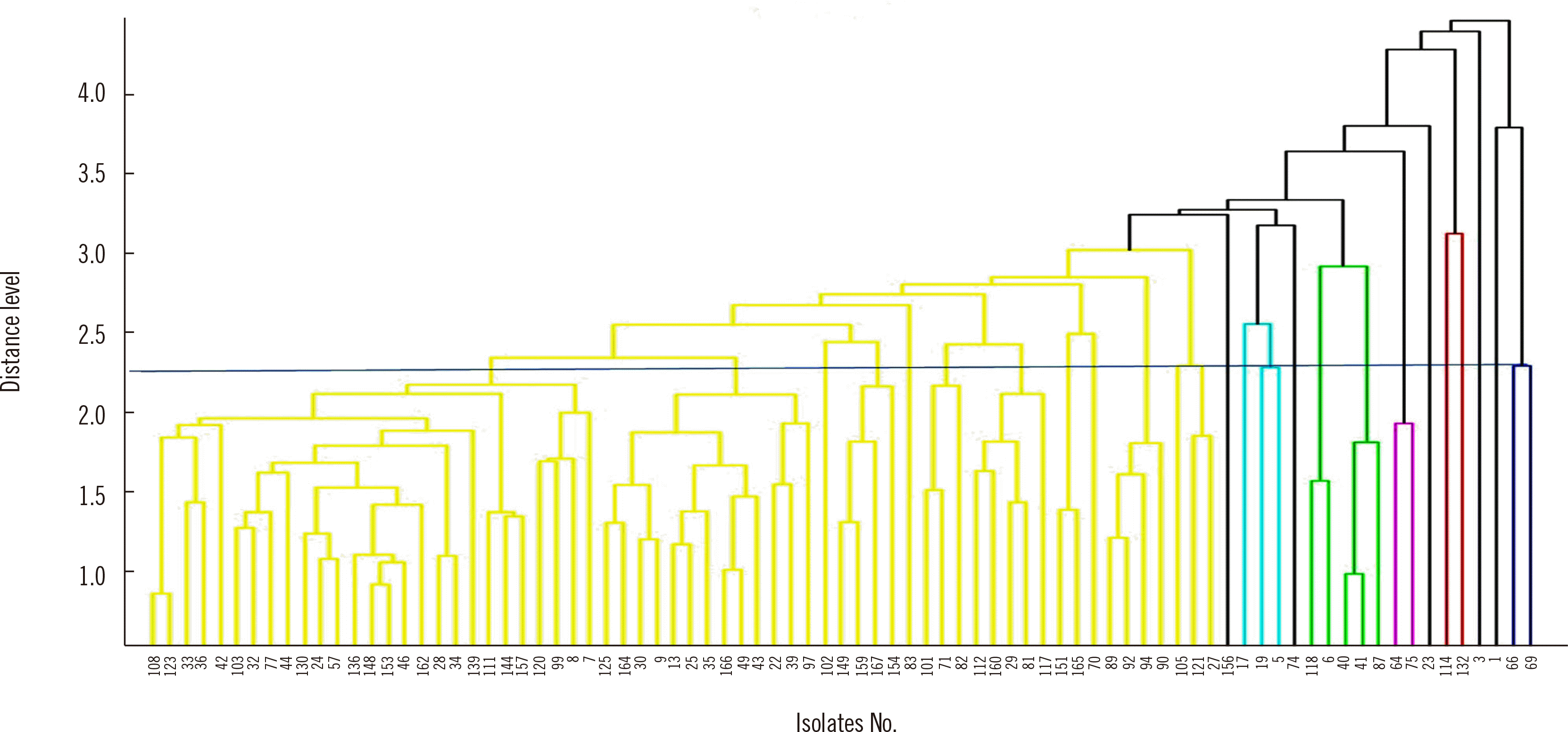
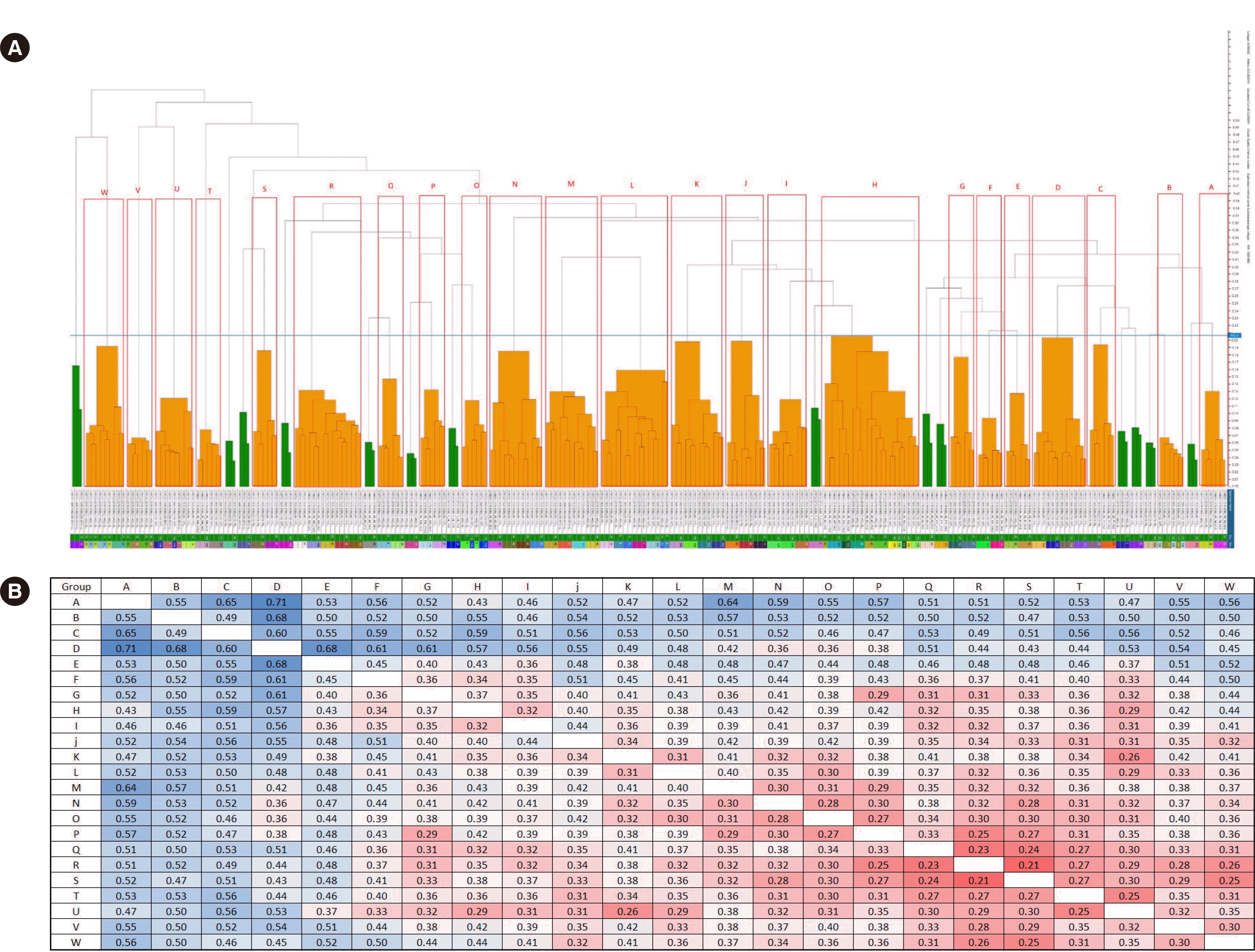
cgMLST divided the 83 ESBL-EC isolates into three groups in the WGS dendrogram (Fig. 1). The number of clusters was 28 using MALDI-TOF MS with a distance in the range of 2.0–2.5 (Fig. 2). These 83 strains were classified into 23 groups with IR Biotyper (Fig. 3A). The average distance value of the integrated 23 groups showed a COV of 0.206; groups S and R were the most similar (COV=0.21), whereas groups A and D were the least similar as independent groups (COV=0.71) (Fig. 3B).
Diversity analysis
The SDI (95% confidence interval) was 0.969 (0.952–0.985) for WGS, 0.865 (0.807–0.924) for MALDI-TOF MS, and 0.974 (0.965–0.983) for IR Biotyper (Table 1). Compared with WGS as a reference, IR Biotyper showed compatible diversity, whereas MALDI-TOF MS did not.
Comparison with WGS
The concordance was 66.3%, and the aRI was 0.173 when MALDI-TOF MS was compared with WGS. The concordance was 84.3%, and the aRI was 0.538 when IR Biotyper was compared with WGS (Table 1).
DISCUSSION
MALDI-TOF MS is a quick, simple, and inexpensive species identification method for diagnosing bacterial infections. However, the performance of MALDI-TOF MS as a first-line epidemiological tool is hindered owing to its limited robustness and exportability [3]. Polymorphism discrimination using MALDI-TOF MS is poor compared with that obtained using amplified fragment length polymorphism for strain typing of ESBL-EC [4]. Although MALDI-TOF MS could be used to detect sequence types of 131 E. coli with a peak at 9,713 m/z, which is useful for sequence type (ST) screening [5], it showed insufficient discriminatory power to determine the clonality of multidrug-resistant Acinetobacter baumannii isolated from patients admitted to intensive care units [18].
IR Biotyper was developed as an easy and fast method, including an enhanced Fourier-transform infrared (FTIR) spectrometer. FT-IR spectroscopy, first introduced in 1950, measures the absorption of energy corresponding to vibration and rotation in the binding of molecules when infrared rays are irradiated onto a sample [19]. Structural information can be obtained based on the presence or absence of a specific covalent bond because only the infrared frequency matches the intrinsic vibration frequency of a specific covalent bond of a molecule [19]. A specific infrared spectrum of all chemical components can be generated to identify microorganisms at the subspecies level, and the results are expressed in a PCA plot, dendrogram, or three-dimensional scattering view [19]. IR Biotyper can be used in an easy and cost-effective manner to differentiate microorganisms at the strain level, commonly referred to as strain typing. Unlike previous FTIR techniques, IR Biotyper is an integrated system that provides a turn‐key solution for microorganism typing [7]. IR Biotyper could accurately cluster Klebsiella pneumoniae strains, and typing results were almost completely concordant with those from PFGE and WGS [7]. Considering its advantages such as low cost and short turnaround time (<3 hours), IR Biotyper could be a promising tool for strain typing that could make real-time outbreak investigation a reality [7].
For diversity analysis, the SDI and aRI were used in this study. SDI is a measure of diversity that considers the number of species present as well as the relative abundance of each species. As species richness and evenness increase, diversity increases [16]. aRI is the corrected-for-chance version of the RI. This correction establishes a baseline by using the expected similarity of all pairwise comparisons between clusters specified by a random model [17]. In this study, when compared to WGS, IR Biotyper showed compatible diversity, whereas MALDI-TOF MS did not, and the overall concordance with WGS was much better with IR Biotyper than with MALDI-TOF MS. IR Biotyper substantially improved the performance of strain typing compared with MALDI-TOF MS. We speculate that the current library of MALDI-TOF MS might not be sufficient to distinguish polymorphisms within species.
A limitation of this study was that the tested ESBL-EC strains were all isolated from poultry industry-related samples. These isolates could be more diverse than those from outbreaks in healthcare facilities, which could influence the results of IR Biotyper. The transmission of antimicrobial-resistant bacteria from non-human sectors has become an important issue to control the antimicrobial resistance problem based on the “One-Health” concept [20]. Epidemiological analysis of resistant bacteria is essential not only in the situation of an outbreak in a healthcare facility but also for studying the transmission of resistant bacteria. IR Biotyper could be a useful candidate for this purpose, despite its insufficient consistency with WGS.
 XML Download
XML Download