

 PDF
PDF Citation
Citation Print
Print



Introduction
Tramadol is a synthetic opioid 4-phenylpiperidine codeine analog, it can bind with μ receptors, but its binding ability with δ- and κ-opioid receptors is low [1]. Tramadol is recommended to treat pain with different degrees of severity [2].
As various tramadol formulations are available, it is increasingly prescribed worldwide as an alternative for high-affinity opioid medication in acute and chronic pain treatment (especially acute perioperative pain) [3]. Routes of tramadol administration are oral, rectal, or parenteral (intravenous, intramuscular, and subcutaneous) routes. Dose titration should be followed according to pain intensity and individual response [4] Tramadol’s efficacy to overcome moderate to extreme postoperative pain has been proven in both day surgery and hospitalized patients. Moreover, it has been demonstrated that using tramadol (orally or parentally) is as effective as a painkiller during perioperative care [5].
Although it is a good painkiller, poisoning can be developed due to tramadol abuse that happens in overdosage, use for an extended period, or interaction with other drugs [6]. Furthermore, severe neurological toxicity involving seizures, respiratory failure, coma, and mild tachycardia with hypertension may result from tramadol overdosage [7]. In addition, many authors indicated that opioids addiction or their consumption for an extended period, such as tramadol and morphine, can cause a lot of adverse effects such as decreasing in sexual activity, dropping in the production of follicle-stimulating hormone (FSH), and testosterone, and hypothalamic-pituitary function abnormalities [8, 9]. It was proved that low-safe and controlled doses of tramadol could be an effective treatment for early or premature ejaculation in mild and severe cases [10].
Although many scientific works studied the side effects of tramadol administration, the impact of tramadol on testicular tissues from a histopathological view needs more investigation. So, this work was designed to investigate histopathological and morphological testicular changes that occur with tramadol administration and compare these testicular changes between adult and juvenile male mice.
Materials and Methods
Drugs and chemicals
Tramadol hydrochloride (125 mg tablets) was supplied from MINA Pharm, 10th of Ramadan City, Cairo Governorate, Egypt, by official request to the dean of the Faculty of Medicine through the pharmacy of Assiut University Hospital.
Animals
From Animal House of Assiut University, a total number of 40 healthy male albino mice were purchased. Animals were maintained under controlled conditions of humidity and temperature. Light:dark cycle was carefully arranged. Standard food and fresh, clean water were supplied ad libitum. The procedures of the experiment were carried out according to the internationally accepted guidelines for the Care and Use of Laboratory Animals and were approved by the Ethics Committee at the Faculty of Medicine, Aswan University, Aswan, Egypt (123-4-17).
Experimental design
Forty mice were divided equally into two major groups, and each one was subdivided into two other groups:
Group I (juvenile group): twenty mice weighting 10–15 g, aged three weeks, were subdivided into:
Group Ia: control group (n=10) received isotonic saline.
Group Ib: tramadol treated group (n=10) received 40 mg/kg/d tramadol, as tablets were smashed and dissolved in normal physiological saline (0.9% orally one time/d for 30 days).
Group II (adult group): twenty mice weighting 25–30 g, aged two months, were subdivided into:
Group IIa: control group (n=10) received isotonic saline.
Group IIb: tramadol treated group: (n=10) received 40 mg/kg/d tramadol, tablets were treated as in group Ib.
Histological examination
Light microscope examination
Testis specimens were collected from all groups, fixed in bouin for 16 to 18 hours, washed with 70% alcohol, dehydrated through ascending graded alcohols and embedded in paraffin, microtome sectioned at 5-μm thickness [11], and stained with the following stains:
H&E stain: for general histological examination.
Masson’s trichrome: for collagen fibers demonstration.
All staining methods were done following the steps described by Bancroft and Layton [12].
Electron microscope examination
Testis specimens were immersed in glutaraldehyde (4%) in cacodylate buffer (pH 7.4) for 24 hours and postfixed for 2 hours in osmium tetroxide (1%) in phosphate buffer solution (PBS). Tissues were rinsed with the same buffer, dehydrated with alcohol, cleared with propylene oxide, and embedded in Epon-812 substitute. Semi-thin sections (0.5–1 μm) were cut with glass knives on the ultratome, and toluidine blue (1%, pH 7.3) was used for staining to be examined on a light microscope. Ultrathin sections (50–80 nm) were cut on a Reichert ultratome, placed on copper grids, and contrasted with uranyl acetate and lead citrate. A transmission electron microscope (Jeol E.M.-100 CX11; Japanese Electron Optics Laboratory Co., Ltd, Tokyo, Japan) was used to examine these sections. Photographing was done at 80 kV in Assiut Electron Microscopic Unit, Assiut University [13].
Immunohistochemical staining
Testis paraffin sections with rabbit anti-mouse caspase-3 polyclonal antibody 1:100 (Chongqing Biospes Co., Ltd., Chongqing, China) were used to demonstrate apoptotic cells. According to O’Callaghan and Jensen [14], the avidin-Biotin peroxidase technique was used. Sections were incubated with primary antibodies diluted; at 1:100 in PBS and left overnight at room temperature. After that, they were stained with an avidin-biotin-peroxidase system using diaminobenzidine as the chromogen (horseradish peroxidase [HRP]; rabbit/mouse/goat [DAB+] code no. K0679, DakoCytomation; Dako, Glostrup, Denmark) in conformity with the staining procedure instruction included in Dako LSAB+ System-HRP (Dako). Sections were washed in distilled water and counterstained with hematoxylin.
Morphometric studies
Morphometric studies were done using a Java-based open-source image processing package, ImageJ (National Institutes of Health, Bethesda, MD, USA). The required parameters were measured in five fields/five non-overlapped sections from three mice from each group. The measured parameters were: the thickness of the capsule, the thickness of the germinal epithelium, the number of positive (+) caspase immune-stained cells and number of distorted seminiferous tubules. Images of stained sections captured at 400× magnification were used for the thickness of the capsule, the thickness of the germinal epithelium and the number of positive (+) caspase immune-stained while for counting number of distorted seminiferous tubules, we used 100× magnification
Statistical analysis
Data were expressed as mean±standard error of the mean. Statistical analysis was performed by multiple t-tests, one-way and two-way repeated-measures ANOVA, followed by post hoc Tukey’s test when appropriate. All statistical tests were directed using GraphPad Prism 7 software (GraphPad Software Inc., San Diego, CA, USA). The difference among groups was considered significant at P<0.05.
Results
Light microscopy results
Hematoxylin and eosin-stained sections results
Tramadol administration (40 mg/kg body weight [BW]/d for 30 days) showed morphological changes in the seminiferous tubules and interstitial tissue in both juvenile and adult groups. The changes were more severe in the juvenile group (Ib) than in the adult group (IIb). Our results showed that group Ia (juvenile control group) showed that the transverse sections in seminiferous tubules appeared regular rounded or oval, separated by interstitial connective tissue and covered by a connective tissue capsule (Fig. 1A, B). The seminiferous tubules were lined by germinal epithelium and surrounded by a basement membrane and a layer of peritubular myoid cells which are flat cells with flat nuclei. The germinal epithelium was formed of spermatogenic cells and supporting Sertoli cells. The spermatogenic cells included spermatogonia, primary spermatocytes, rounded spermatids, and sperms. In between tubules, the interstitial cells of Leydig with rounded nuclei and acidophilic cytoplasm were noticed surrounding the blood capillaries (Fig. 1A, C).
The spermatogonia were arranged as one raw next to the basement membrane with oval nuclei. The primary spermatocytes were represented in the next row of germinal epithelium cells, usually two or three layers. They were the largest of the whole series, having rounded large nuclei with different stages of meiotic division (Fig. 1C). The rounded or early spermatids revealed rounded nuclei and presented in multiple rows (Fig. 1C).
In group Ib (juvenile tramadol-treated group), sections-stained with H&E stain showed severe distortion in most seminiferous tubules; they were irregular in shape, widely separated, and degenerated. The thickness of the germinal epithelium was reduced significantly compared to the control group with irregular empty spaces. Few or no sperms were found. However, few tubules appeared nearly similar to the control group (Fig. 1D, E). Shrinkage with increased acidophilia was observed in some tubules (Fig. 1D, F), while others were severely shrunken up to appear star-shaped (Fig. 1G). In addition, fusion occurred between some tubules with swelling of some cells and irregular vacuoles or empty spaces between cells (Fig. 1F). Congestion of blood vessels was noticed (Fig. 1D, E).
In some sections, exfoliation, vacuolation, and pyknosis of germ cells were prominent features (Fig. 1G–I). The basement membrane showed irregularity with increased thickness (Fig. 1F). The interstitium showed transudation, increased interstitial cellular mass, and congested blood vessels (Fig. 1D, E). Leydig cells were seen with darkly stained nuclei and deeply acidophilic cytoplasm (Fig. 1H, I).
Group IIa (adult control group), the same as group Ia, rounded or oval regular seminiferous tubules enclosed by a connective tissue capsule and surrounded by interstitial connective tissue were detected (Fig. 2A). The germinal epithelium was seen lining the tubules and basement membrane surrounding them. The germinal epithelium was formed of spermatogenic cells (spermatogonia, primary spermatocytes, rounded spermatids, and sperms) and supporting Sertoli cells (Fig. 2A, B). Interstitial cells with round nuclei and acidophilic cytoplasm were observed (Fig. 2B).
One raw next to the basement membrane, spermatogonia with ovoid nuclei were arranged. Primary spermatocytes were found in the following raw germinal epithelium cells (usually two or three layers). They were the largest of the whole series, having rounded large nuclei with different stages of meiotic division (Fig. 2B). The rounded spermatids revealed rounded nuclei and presented in multiple rows. At the same time, elongated spermatids had condensed elongated nuclei and tails that protrude to the lumen (Fig. 2A). The Sertoli cells had large pale nuclei with prominent nucleoli resting on the basement membrane (Fig. 2A). Large Leydig cells were found in the interstitium with round nuclei and light acidophilic cytoplasm adjacent to a blood vessel (Fig. 2A).
Group IIb (adult tramadol-treated group), sections stained with H&E stain showed distortion in most seminiferous tubules. They were widely separated with irregularity. Furthermore, a thick, irregular testicular capsule was observed (Fig. 2C). Few numbers of tubules were more or less the same as in the control group (Fig. 2C). Exfoliation of dense acidophilic germ cells with vacuolation, increased mitotic activity, and irregularity of basement membrane were detected (Fig. 2C–E). Transudation, congestion of blood vessels, and increased interstitial cellular mass were found in the interstitium. Deeply acidophilic cytoplasm and darkly stained nuclei were detected in Leydig cells (Fig. 2E, F).
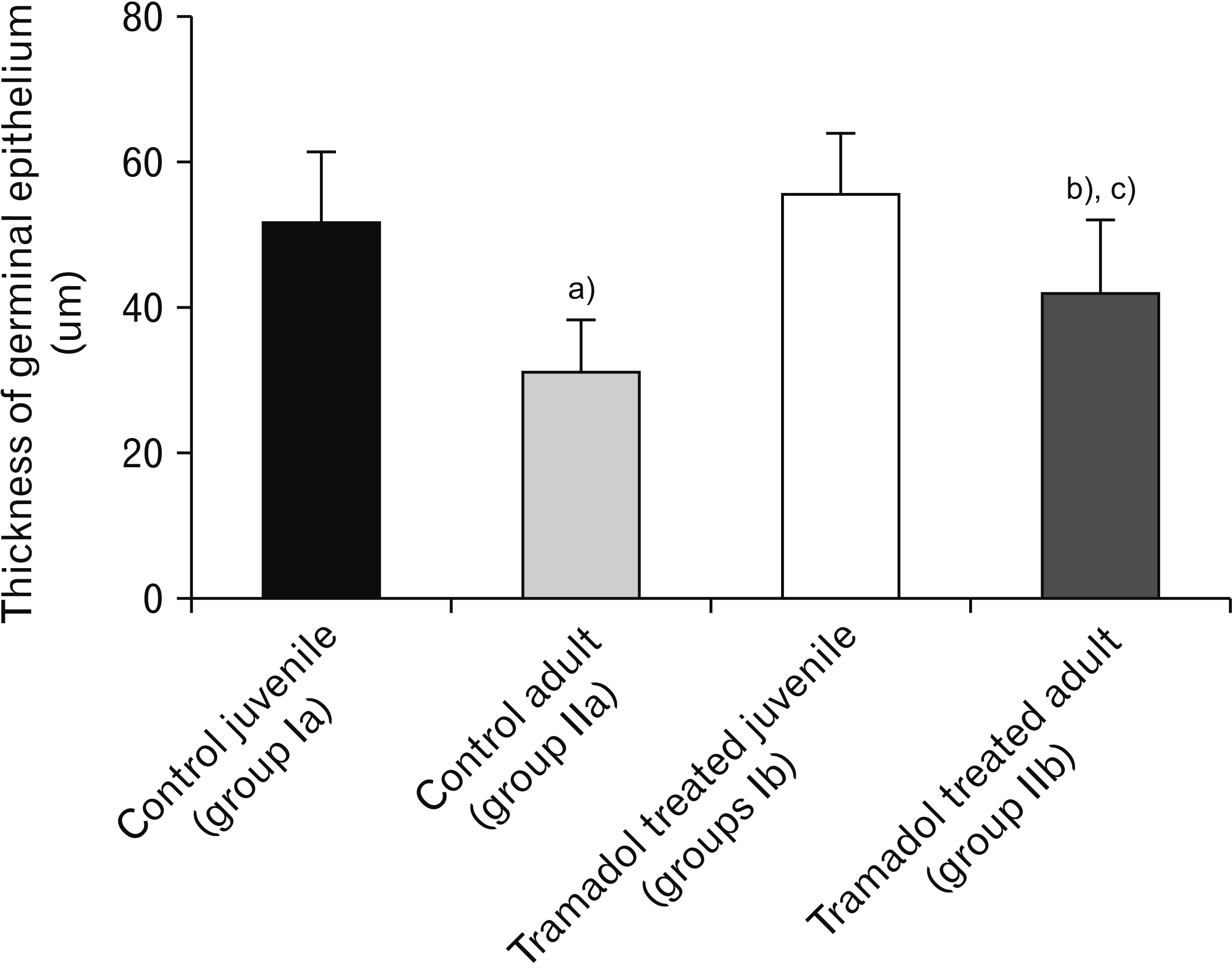
The thickness of germinal epithelium decreased significantly in Tramadol-treated juvenile group when compared with the control juvenile group (P<0.001). The tramadol-treated adult group showed a significantly reduced thickness of germinal epithelium versus the control adult group (P<0.001). The thickness of germinal epithelium was reduced considerably in tramadol treated juveniles than Tramadol treated adult groups (P<0.001; Fig. 3).
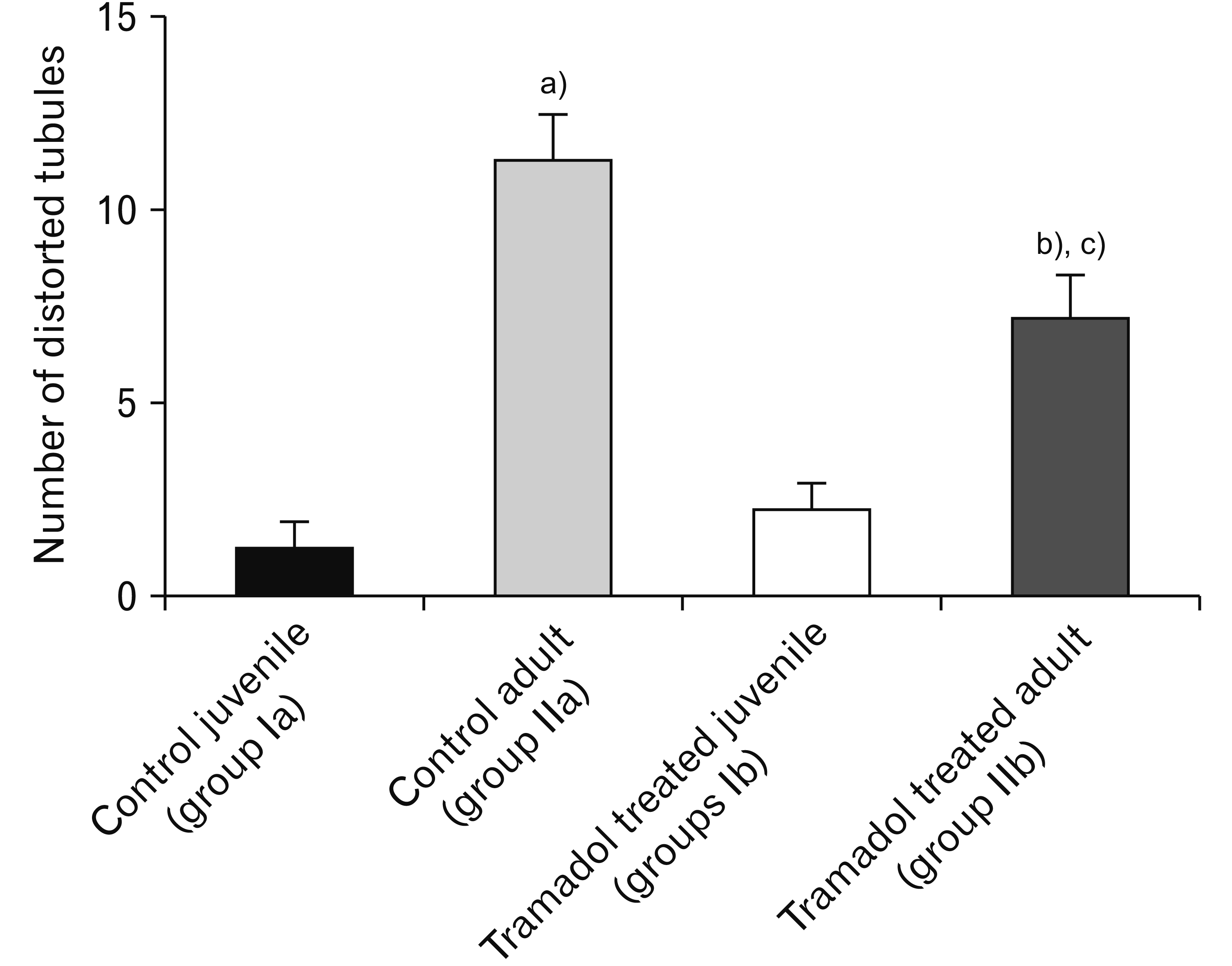
The number of distorted tubules was increased significantly in Tramadol treated juvenile group in comparison with the control juvenile group (P<0.001). Tramadol treated adult group has a significantly increased number of distorted tubules than the control adult group (P<0.001). Tramadol treated juvenile group showed a significantly higher number of distorted tubules than the Tramadol treated adult group (P<0.001; Fig. 4).
Masson’s trichrome stained sections
In group Ia, regular distribution of collagen fibers in the capsule with few depositions around blood vessels was detected during Masson trichrome stained sections examination (Fig. 5A). Group Ib, most of the seminiferous tubules showed dense deposition of the collagen fibers, causing thickening in the capsule and around blood vessels, as well as the basement membrane (Fig. 5B). Group IIa, in the capsule, collagen fibers showed normal distribution, while around blood vessels and basement membranes, few depositions of collagen fibers were found (Fig. 5C). Group IIb, collagen fibers showed dense deposition surrounding basements membrane, blood vessels, and in the capsule (Fig. 5D).
The capsule thickness was increased significantly in Tramadol treated juvenile group in comparison with the control juvenile group (P<0.001). Tramadol treated adult group has a significantly increased capsule thickness versus the control adult group (P<0.05). An insignificant difference was obtained between the Tramadol-treated juvenile and Tramadol-treated adult groups (Fig. 6E).
Semithin section stained with toluidine blue
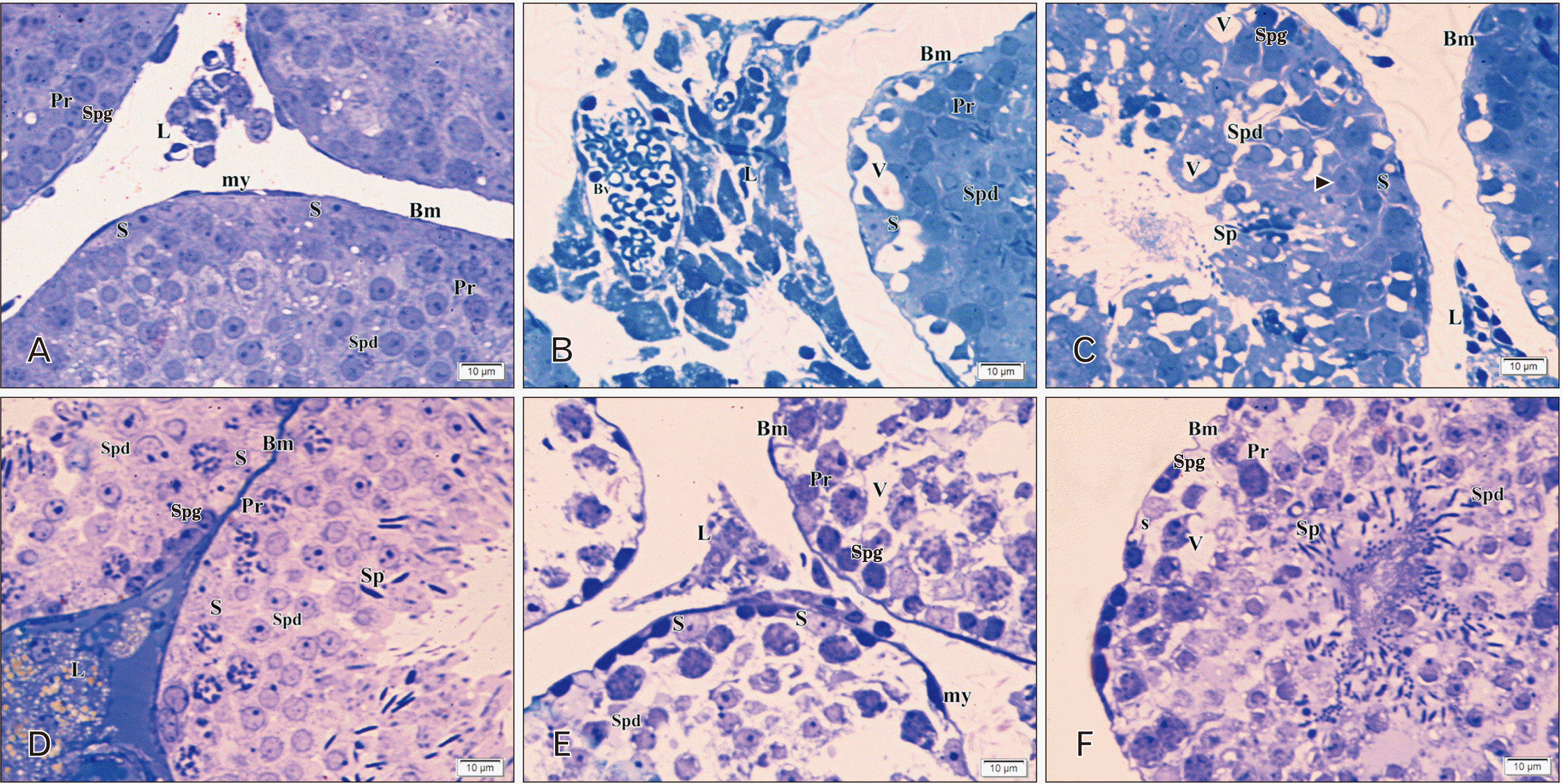
In group Ia, the seminiferous tubules appeared separated by interstitial connective tissue. A thin regular basement membrane surrounded them with myoid cells which were flat cells with flat nuclei. Spermatogonia had ovoid-shaped nuclei resting on the basement membrane. Primary spermatocytes had rounded large central nuclei in different stages of meiosis. Early or rounded spermatids showed acrosomal caps covering the rounded nuclei. The Sertoli cells were characterized mainly by their vesicular nuclei and resting on the basement membrane between spermatogenic cells. Leydig cells with spherical vesicular nuclei were seen between seminiferous tubules (Fig. 7A). Group Ib semithin sections revealed distorted irregular seminiferous tubules separated by wide interstitial spaces with a marked increase in interstitial cell mass, and deeply stained nuclei of interstitial cells were frequently observed in comparison to control group with hazard arranged germ cells with multiple irregular empty spaces. Some Sertoli cells are detached from the basement membrane with depletion of sperms (Fig. 7B, C).
In group IIa, rounded and oval seminiferous tubules were separated by interstitial connective tissue. A thin regular basement membrane surrounded each seminiferous tubule with myoid cells, which appeared flat. Spermatogonia appeared with ovoid-shaped nuclei resting on the basement membrane. Primary spermatocytes had rounded large central nuclei in different stages of meiotic division. Early spermatids showed acrosomal caps covering the rounded nuclei. Sertoli cells appeared with vesicular nuclei, resting on the basement membrane in between spermatogenic cells. Sperms were seen in the lumen attached to Sertoli cells. Leydig cells with spherical vesicular nuclei and vacuolated cytoplasm were wedged in between seminiferous tubules (Fig. 7D). In group IIb, the seminiferous tubules appeared distorted, irregular, and separated by wide interstitial spaces compared to the control group. Some tubules showed hazard arranged germ cells with multiple irregular empty spaces, and irregular shrunken Sertoli cells with irregular basement membranes (Fig. 7B, C).
To explore whether treatment with tramadol can induce apoptosis, we evaluated the expression of the apoptotic parameter caspase-3 in testicular tissues using the immunohistochemistry technique.
Group Ia: Immunostaining with caspase-3 showed scattered distribution of weakly positive apoptotic cells, especially in spermatogonia (Fig. 6A). In group Ib a significantly increased frequency of strongly positive apoptotic cells in spermatogonia and interstitial cells of Leydig (Fig. 6B). In group IIa, few weak positive caspase-3 immunostained cells were observed (Fig. 6C). Group IIb, significantly increased the frequency of moderately positive apoptotic cells in nearly all spermatogenic cells and interstitial cells of Leydig (Fig. 6D).
The tramadol-treated juvenile group showed significantly higher caspase-3 positive cells than the control juvenile group (P<0.001). Also, the Tramadol-treated adult group has a significantly increased number of caspase-3 positive cells than the control adult group (P<0.001). An insignificant difference was obtained between the Tramadol-treated juvenile and Tramadol-treated adult groups (Fig. 6E).
Electron microscopy results
Group Ia
Electron microscopic examination for this control group showed the presence of a regular thin basement membrane around the seminiferous tubule with myoid cells (with flat nuclei), while spermatogenic cells and Sertoli cells were lining them (Fig. 8A). Settling on the basement membrane, Sertoli cells were found with large basally located nuclei and prominent nucleoli with the fine distribution of granular chromatin. Their cytoplasm had smooth endoplasmic reticulum, several mitochondria, and lysosomes (Fig. 8A, B). Spermatogonia with round nuclei were found in the basal compartment (Fig. 8B), and intercellular junctions were intact (Fig. 8A). Primary spermatocytes showed large rounded nuclei with finely granular chromatin and clumps of heterochromatin, and their cytoplasm was rich in mitochondria (Fig. 8A). Rounded spermatids were seen with spherical nuclei and finely granulated chromatin distribution. The nuclear membrane was thickened in a particular area and covered by the acrosomal cap and vesicle. The acrosomal cap was seen over one pole of the nucleus. The cytoplasm contained characteristic peripherally situated mitochondria with clear matrix and dilated cristae (Fig. 8C). Clumps of Leydig cells were present in the interstitium and had round or oval euchromatic nuclei, with several mitochondria and smooth endoplasmic reticulum in the cytoplasm (Fig. 8D).
Group Ib
The germinal epithelium of most seminiferous tubules, showed cellular loss, severe distortion, and extreme vacuolation in the cytoplasm. The intercellular junctions were not observed. A thickened irregular basement membrane surrounded them with myoid cells (Fig. 8E, F).
In some tubules, Sertoli cells showed irregular nuclei with the cytoplasm appearing rarified with empty spaces, devoid of organelles apart from abnormally shaped mitochondria. The spermatogenic cells appeared dark with various signs of degeneration varying from irregular dark nuclei and rarified cytoplasm. Degenerated elongated spermatids with irregular elongated acrosomal caps and irregularly arranged mitochondria were detected. The interstitial cells of Leydig appeared with rounded electron-dense nuclei, dilated smooth endoplasmic reticulum, and disrupted cell membrane (Fig. 8E–G).
Group IIa
The seminiferous tubules of the control adult testis were lined by spermatogenic cells and the supporting Sertoli cells. Sertoli cells were rested on the basement membrane, and had large basally located nuclei with prominent central nucleoli and finely granular chromatin distribution. The Sertoli cell cytoplasm was studied with numerous mitochondria and smooth endoplasmic reticulum. Primary spermatocytes showed large rounded nuclei with finely granular chromatin and clumps of heterochromatin (Fig. 9A, B). The rounded spermatids were seen with spherical nuclei with finely granular chromatin distribution. The cytoplasm contained characteristic peripherally situated mitochondria (Fig. 9C). The interstitium showed Leydig cells with oval to rounded euchromatic nuclei. The cytoplasm revealed numerous smooth endoplasmic reticulum (Fig. 9D).
Group IIb
In this group, most of the seminiferous tubules showed a predominance of Sertoli cells with loss of most spermatogenic cells. The spermatogenic epithelium was severely distorted with excessive cytoplasmic vacuolation and cellular loss. An irregular basement membrane surrounds them with contracted dark myoid cells. Sertoli cells showed small nuclei in some tubules, and the cytoplasm was rarified with empty spaces (Fig. 9E). The spermatogenic cells showed various signs of degeneration with irregular nuclei (Fig. 9F).
Degenerated rounded spermatids with dark nuclei and abnormal chromatin distribution (Fig. 9F, G). A cluster of electron-dense Leydig cells was present with dark nuclei, and dilated smooth endoplasmic reticulum and lysosomes could be observed (Fig. 9H).
Discussion
Because of its high availability with low price, tramadol is relatively popular among Egyptian youth for alleged uses in premature ejaculation treatment and its ability to treat different pain degrees (moderate to severe, acute, and chronic pain) [10, 15, 16].
The objective of this study was to investigate light and electron microscopic changes observed in seminiferous tubules and interstitial cells after tramadol administration in juvenile and adult mice.
Light microscopic examination of control groups (Ia and IIa) revealed a typical histological picture of seminiferous tubules; they appeared regularly with normal spermatogenesis and interstitial tissue. In addition, spermatogenic cells appeared normal and regularly organized. Moreover, the germinal epithelium thickness was greater than treated groups, as indicated by morphometrical studies. These results were in accordance with several studies that reported normal spermatogenesis and normal intertubular tissue [17, 18].
Tramadol administration (40 mg/kg BW/d for 30 days) showed structural changes in the seminiferous tubules and interstitial tissue in both juvenile and adult groups. These changes were more severe and prominent in the juvenile group (Ib) than in the adult group (IIb) that received tramadol for the same duration. In some animals, seminiferous tubules were largely separated from each other with loss of their normal contour. Some tubules’ depletion of spermatogenic cells with excessive vacuolations was observed, while shrinkage of tubules, lumen obliteration, and increased acidophilia were found in other tubules. The number of distorded tubules was higher in treated group compared to control, with more increase in juvenile group. These findings were in line with the results obtained by Abdellatief et al. (2015) [17], they investigated the testicular changes in rats affected by long-term tramadol administration and attributed this to the deposition of homogenous hyaline acidophilic Periodic acid-Schiff-positive material. In contrast, Salama et al. (2003) [19] explained the wide separation between the seminiferous tubules. It may be related to the degeneration of lymphatic vessels leading to the oozing of excess lymphatic exudate, resulting from the accumulation of free radicals and reactive oxygen species (ROS). Dilated congested intertubular blood vessels and an increase in collagen fibers deposition were also detected. The thickness of the capsule in both treated groups was significantly increased compared to the control groups, and this was in agreement with the observations obtained by Awadalla and Salah-Eldin (2005) [20] and Essam Hafez et al. (2015) [21]. They investigated the hepatorenal toxicity in adult albino male rats under the tramadol effect. Moreover, Elkhateeb et al. (2015) [22] indicated that a significant decrease in glutathione peroxidase and a significant increase in hepatic and renal malondialdehydes under tramadol administration might result in oxidative stress and hepatorenal toxicity in adult albino rats.
The wide intercellular spaces noticed among spermatogenic cells may be due to apoptosis and shrinkage of the cells. These findings were similar to the results obtained by Mohamed et al. (2014) [23], who examined cadmium impact on testicular tissues in adult male albino rats. They indicated that widening of intercellular spaces is associated with disrupting the blood-testis barrier, which allows passage of toxic agents among the cells under the effect of ROS on spermatogenic cells. Furthermore, Sertoli cells and most the spermatogenic cells showed cytoplasmic vacuolations. These changes most likely developed from endoplasmic reticulum vesiculation and dilatation as well as mitochondrial swelling. Nolte et al. (1995) [24] reported that significantly larger vacuoles are phagocytotic vacuoles that continue to remain after necrotic germ cells digestion.
Marked depletion of spermatogenic population and many spermatogenic cells are replaced by vacuoles. Focal disorganization and shrunken or atrophy of seminiferous tubules detected in the present study were consistent with the findings of Caju et al. (2011) [25] and Abdellatief et al. (2015) [17], who suggested that these changes were related to endocrine and paracrine functions disturbance through disturbing luteinizing hormone (LH), estradiol, somatotropin, somatostatin, prolactin, and gonadotrophin-releasing hormone levels acted on the hypothalamus or pituitary gland. Moreover, Ahmed and Kurkar (2014) [26] stated that tramadol causes a significant decrease in the plasma levels of LH, FSH, testosterone, and total cholesterol, whereas it causes an increase in the levels of prolactin and estradiol.
Electron microscopic examination of tramadol treated groups (Ib and IIb) showed that the basement membrane was irregular, discontinuous, and surrounded by scattered collagen fibers. The same observations were recorded by Ahmed and Kurkar (2014) [26]. They explained that oxidative stress and overproduction of nitric oxide result from a reduction of scavenger antioxidant enzymes, caused by tramadol treatment.
Many studies have explained the relationship between oxidative stress and tramadol-induced reproductive toxicity, as tramadol interacts with molecular oxygen and can initiate a cascade of reactions producing numerous free radicals, such as superoxide and hydrogen peroxide, by which induction of lipid peroxidation and reduction of antioxidant enzymes in different tissues occurred [27].
Berruti (2006) [28] stated that basement membrane thickness which is observed in this study may be related to increased glycosaminoglycans and proteoglycans production. He considered the collagen type IV in the basement membrane as a defense mechanism against free radicals’ damaging activity. Moreover, Lombardo et al. (2005) [29] reported that certain stimulants might induce myoid cells to produce more collagen and extracellular matrix, responsible for the thickening of the basal lamina. Also, decreasing the proteolysis rate in the extracellular matrix or increasing collagen production from myoid cells may be the reason for the deposition of collagen into the basement membrane [30].
In this study, sloughing of germinal epithelium and seminiferous tubules atrophy were primarily detected in the juvenile treated group. Heidari et al. (2012) [31] indicated that the effect of opioids on the pituitary or hypothalamus leads to inhibiting LH release, which causes a decrease in testosterone. Furthermore, Sorge and Stewart (2006) [32] reported that testicular metabolism impairment followed by hypoplasia of germinal cells and atrophy of tubules could occur due to increased basement membrane thickness.
Coinciding with our results, it was found that tramadol induced suppression of FSH and LH [17, 25]. This could lead to a reduction in tubular diameter and degeneration of spermatogenic cells. Consequently, this would lead to widening of the peritubular tissue and thickening in the basement membrane observed in Masson’s trichrome-stained sections of the present study. This observation was in line with the results of [33], who accounted for the widening of peritubular tissue due to the withdrawal of gonadotrophic stimulation after hypophysectomy. Some authors explained germ cell sloughing because affected Sertoli cells induced a decrease in seminiferous tubule fluid secretion, which further resulted in apical sloughing or shedding and germ cell death. Others attributed the sloughing of germ cells to the organization of germ cells held in place by a close association between their membranes and specialized junctions of cell membranes of Sertoli cells [33, 34]. In the present study, ultrastructural analysis of testicular tissue revealed extensive degenerative changes in spermatogenic and Sertoli cells in some seminiferous tubules up to the complete depletion of cells. These features agreed with the findings of Jezek et al. (1993) [35]. They explained Sertoli cell vacuolation due to metabolic disturbance. It is well known that Sertoli cells are influenced mainly by testosterone, producing androgen-binding protein and other products under the control of FSH and androgens [36]. We postulated that tramadol administration disturbed this link and induced morphological alteration due to testosterone deprivation.
In this study, our result showed that tramadol triggered atrophy and apoptosis by the stimulation of apoptotic molecules via increased activation of caspase-3 in testes which was revealed by immunohistochemistry staining, as tramadol led to the increase of the number and intensity of caspase-3 positive cells in both tramadol treated groups compared to control groups. The damage to the mitochondria may explain this. Mohamed and Mahmoud (2019) [37] reported that chronic tramadol consumption induced oxidative damage leading to inflammation, and apoptosis. This may be explained by activation and expression of the inflammation and apoptosis markers and a decrease in the anti-apoptotic proteins. Another explanation of apoptosis is the increased DNA fragmentation induced by exogenous opioids in men [38].
The sperms and spermatids were infrequently seen in the present work. This may be explained by the release of cytotoxic reactive oxygen species that cause tissue injury, especially in the late phase of differentiation of germ cells [39]. Moreover, Eddy (1999) [40] mentioned that the spermatocytes arrested in prophase meiosis were then eliminated at the late pachytene phase by apoptosis, resulting in the absence of spermatids.
Furthermore, electron microscopic examination of the testes of treated groups (Ib and IIb) showed some spermatogenic cells rupture with small dark nuclei. The same observations were detected by Newton et al. (2005) [41] and Tamura et al. (2012) [42], who said that the free radicals facilitate the release of lysosomal enzymes into cytosol with subsequent oxidation of the protein components of the cell, causing their fragmentation.
Leydig cells nuclei were irregular with dilated smooth endoplasmic reticulum and many vacuoles in the cytoplasm in both tramadol-treated groups. These findings agreed with the results obtained by Ghoneim et al. (2014) [9]. In addition, Ahmed and Kurkar (2014) [26] detected a reduction in Leydig cells numbers with their malformation. An extreme increase in testicular nitric oxide level occurred with repeated tramadol administration in rats for several weeks. These results could suggest Leydig cells’ hypofunction and a decrease in testosterone production [43-45]. Recently, Bassiony et al. (2020) [46] stated that patients treated with tramadol were more likely to have lower free testosterone levels and lower sperm counts and vitality. This was accompanied by higher prolactin levels and abnormal forms of sperm compared with controls. These findings may be explained by Popovic et al. (2009) [47], who related Leydig cells degeneration to oxidative stress that generates ROS reacting with protein, nucleic acids, and lipids, causing function and structure changes and finally damage and even death of the cells. Moreover, El-Gaafarawi (2006) [48] reported that tramadol directly affected the hypothalamic-pituitary axis inhibiting the secretion of both FSH and LH, which are responsible for normal spermatogenesis in male rats, and reduction of testosterone secretion, which could be involved in the involution of the seminiferous epithelium.
In conclusion, tramadol has a deleterious effect on testicular tissues of both juvenile and adult male albino mice, but tramadol impact was more evident in juvenile mice. Tramadol abuse, especially in the juvenile treated group, may solve the association between its use and unexplained late fertility, such as damage to testicular tissues, apoptosis, lipid peroxidation, induced oxidative stress reducing sperm production. So, tramadol abuse should be stopped and used only with a medical prescription, to avoid its harmful effect.
Attention from the Egyptian Institutes of education and health, together with families, is required to prohibit tramadol overuse, reduce its popularity, and decrease its addiction among young people.





 XML Download
XML Download