

 PDF
PDF Citation
Citation Print
Print



Introduction
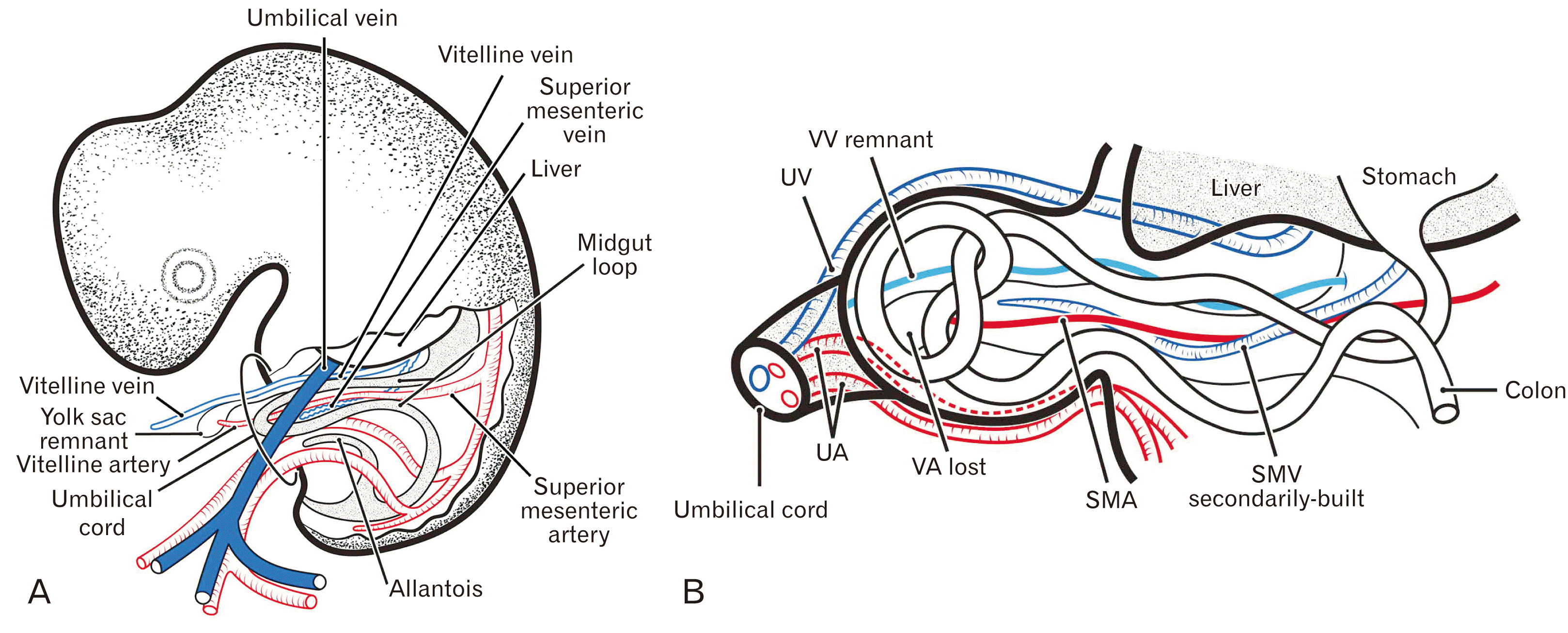
In contrast to the superior mesenteric vein (SMV), which develops secondarily in the midgut mesentery without contribution from the vitelline vein (VV), the vitelline or omphalomesenteric artery is thought to directly transform into the superior mesenteric artery (SMA) [1, 2]. Although the development and growth of the midgut loop and its blood supply and drainage have been major interests in embryology [3-5], less is known about the upstream or extra-embryonic portion of the vitelline artery (VA) and VV. Similarly, there have been few photographic evaluations of yolk sac morphology in human embryos. A digitized series of two digitally reproduced embryonic morphology (DREM) embryos of Carnegie stages (CS) 13–18 (http://virtualhumanembryo.lsuhsc.edu/) show both the VA and VV running freely outside the mesentery in the extra-embryonic coelom or herniation sac. However, the posterior termini of the candidate VA and VV were not clearly visible in these virtual embryos.
Micro-computed tomography has shown that, following “regression of the omphaloenteric duct” in rat embryos at ED12, a combination of the VA and VV maintains its long course through the yolk stalk in the extra-embryonic coelom until ED13 or later [6]. In addition, that study showed a switching of midgut venous drainage in the extra-embryonic coelom from short tributaries of the VV (ED12) to an adult-like long SMV (ED13). Notably, the contents of the yolk stalk seemed to differ between mouse and human embryos at the same stage. An excellent series of 3D reconstructions of human embryos has shown that the yolk stalk contains the omphaloenteric duct as well as the vitelline vessels at CS 14–18 (approximately 4–6 weeks) [2]. Therefore, at CS 19–20 or approximately 6 weeks, the human VA seemed to transform into the SMA without a distal course outside the midgut mesentery.
The human yolk sac seems to be much smaller than that of experimental animals at the same stage [7, 8]. To our knowledge, to date there has been no histological demonstration of the human yolk sac supplied with the VA and VV. Because of the extreme difficulty of finding histological sections of human embryos at 3–4 weeks, we assessed 20 specimens of CS 15–19 (approximately 5–6 weeks) to provide photographic evidence of 1) candidates for the yolk sac remnant, 2) a regressing VA and a persistent VV in the extra- and intra-embryonic coelom, and 3) the topographical anatomy of the most anterior portion or apex of the midgut loop (midgut apex), including the omphaloenteric duct.
Materials and Methods
This study was performed in accordance with the Declaration of Helsinki 1995 (as revised in 2013). Paraffin-embedded histological sections were obtained from 20 embryos of stages 15–19 [9] and crown-rump length (CRL) 7.5–20 mm (approximately 5–6 weeks). The sectional planes of 14 specimens were horizontal, whereas the planes of the other six specimens were sagittal. All sections were part of the large collection maintained by the Department of Anatomy of the Universidad Complutense, Madrid, Spain, with these embryos being the products of miscarriages and ectopic pregnancies obtained from the Department of Obstetrics of the University. No information was available on the genetic background of the embryos and/or the cause of abortion. The sections were stained with hematoxylin and eosin (H&E) or Azan. This study was approved by the Ethics Committee of Complutense University (B08/374). All photographs were taken with a Nikon Eclipse 80 (Nikon, Tokyo, Japan). The identification of the intestines on each section was based on our previous studies [10-12].
Results
Figs. 1–7 are arranged in order from smaller to larger specimens, although this order did not always correspond to the stage; for example, the embryo in Fig. 2D–E is younger than that in Fig. 2A–C according to the morphologies of the heart and liver. Since all seven specimens shown in Figs. 1–6 were at 5 weeks of gestation, a figure-to-figure explanation is necessary at the head of this section for introduction of the individual difference in morphology. Regardless of whether the umbilical cord containing the extra-embryonic coelom protruded superiorly or inferiorly, the upstream portion near the placenta was described as the “distal” portion. Thus, the VA and VV, along with the umbilical vessels, protruded distally beyond the midgut apex. When the dorsal, intramesenteric course of the VA was called “SMA”, the VA was regarded as a free distal artery outside the mesentery. The sagittal sections of six specimens (Table 1) were unlikely to contain a distal part of the VA, possibly due to tissue injury during the histological procedure.
Fig. 1 may be most difficult to understand the topographical anatomy because abdominal structures are packed in an extremely narrow space. However, as a diagram summarizes (Fig. 1K), a small vacuole-like yolk sac is surrounded by a VV and two branches of the VA. Fig. 2, sagittal sections containing two embryos, exhibits a fact that the yolk sac remnant is likely to exist near the heart and liver or in the dorsal site in the intra-embryonic celom. Fig. 3 may be easy to read because of the large celomic cavity. The yolk sac remnant is seen in the extra-embryonic celom (Fig. 3A–C) and the VV runs freely in the celom to approach the duodenum (Fig. 3E, F). Although Fig. 4 is composed of many panels, a half of them (Fig. 4A–F) shows details of a close relation, even a communication, between the VV and yolk sac (summarized in Fig. 4L). Although the yolk sac remnant is absent, Fig. 5 is characterized by 1) the dilated veins (Fig. 5B–E) and 2) a rarely long, longitudinal cut of the VA (Fig. 5G). Fig. 6 focuses the VV and VA in the extra-embryonic celom: they have free ends in the celom without any yolk stalk-like structure (Fig. 6F). Finally, Fig. 7 shows a limited 6 weeks’ embryo that had a yolk sac remnant in spite of the large body size (summarized in Fig. 7A).
In all specimens, the midgut apex reached the extra-embryonic coelom in or adjacent to the umbilical cord. Candidate yolk sac remnants were found in 13 of the 20 specimens, although these remnants in horizontal sections were usually small, round vacuole-like structures. The yolk sac remnant often communicated with (Fig. 4E) or opened to (Fig. 7F) the VV or SMV. In smaller specimens (CRL <12 mm), the yolk sac remnant was present on the distal side of the midgut apex, but, in larger specimens (CRL >16 mm), the remnant was located on the proximal or posterior side of the apex. In sections containing a yolk sac remnant, the yolk stalk was thick and easy to identify (Figs. 3A, 4A). However, the yolk stalk on sagittal sections was often unclear, possibly due to oblique cutting (Fig. 2A–C). The SMV was always present, usually as a series of irregularly-shaped slits of the mesentery tissue (Figs. 1J, 3G, and 5B). Thus, the luminal surface of the SMV was not smooth. We did not found candidates of the epithelial-lined omphaloenteric duct originating from the midgut apex and extending distally along or near the VA and/or VV.
The VA in the extra-embryonic coelom of the umbilical cord usually ran distally along the yolk stalk on the distal side of the midgut apex, with or without the concomitant VV (Figs. 1C and 3B, 4C, 5A–D). In larger specimens, however, the VA was absent, being replaced by the SMA within the mesentery (Fig. 7D, G, Table 1). In the larger specimens, strictly, the VA regressed in the distal side of the midgut apex and the proximal long portion remained as a SMA. The VV was present in all specimens, irrespective of whether its course was partial or complete, with the posterior ending of the VV at the mesentery near the duodenum being present in all of the smaller specimens (Figs. 3F, 4J and 5D and 7G). Therefore, the VV was likely to regress in the posterior or proximal course. Conversely, in the extra-embryonic coelom, the VA started to regress earlier than the VV. Branches of the VA and tributaries of the VV were rarely observed near the yolk sac remnant in the yolk stalk (Fig. 1A). Sagittal sections showed that the VV curved around the duodenum, resulting in a connection between the pre- and post-duodenal portal veins (Fig. 2B, C).
Overall, a vacuole-like remnant of the yolk sac was consistently present in embryos smaller than 10 mm CRL, but was absent from 7 of the 11 larger embryos (11–20 mm CRL). The VA disappeared in the distal side of the midgut apex in larger embryos (>16 mm CRL), but the VV was likely to still remain. Fig. 8A is a diagram showing a whole embryo at 5 weeks with a set of the vitelline vessels and yolk sac remnant: the sac was thinner than the umbilical artery and vein and it had lost a capillary network for the vascular supply. At and until 6 weeks, The VA and yolk sac were almost always lost, while the VV still ran freely through the extra-embryonic celom or herniation sac (Fig. 8B). Therefore, Fig. 7 exhibits a rare specimen containing the yolk sac remnant even at 6 weeks.
Discussion
Although the number of specimens examined was limited, the present study was likely the first systematic examination of the upstream portions of the VA and VV in human embryos. The yolk sac remnant was strikingly small and sandwiched between the VA and VV at the distal end of the yolk stalk. This was quite different from the classical image of a bulky yolk sac surrounding blood capillaries at 5–6 weeks in textbooks of embryology. In their excellent 3D-diagrams, Hikspoors et al. [4] showed multiple tributaries of the VV surrounding and draining the yolk sac at CS 11 and 12. According to them, however, any of such tributaries seemed to disappear suddenly at CS 13 or within approximately 5 weeks. To our regret, we have no available histological information before and at the drastic change. In addition, we ensured a fact that the superior mesenteric vein develops secondarily as a sequence of tissue slits in the mesentery.
The present study also demonstrated a considerable difference in topographical anatomy of the VA, VV and yolk sac remnant in embryos at 5 weeks. In short, this seemed to be a result of extremely rapid changes of the stage. In spite of the almost same size, some embryos were characterized by long courses of the VV and VA, while the others had the vessels and midgut loop packed tightly in a narrow celomic cavity. Likewise, whether the vessels were thin or dilated also provided different morphologies. Thus, in addition to a great contribution of the increased length of intestines, the vascular dilation might increase a volume of the extra-embryonic celomic cavity. A key factor for understanding the different anatomy seemed to be the yolk sac: the regression occurs first in the yolk sac, and subsequently the VA seemed to disappear from the distal end. Growth of the midgut loop might facilitate the VA regression.
The secondary yolk sac originates from membrane folds of the primary, large sac [13, 14] and is followed by remodeling of the vitelline vessels [15]. In human embryos, this remodeling may drastically reduce vascular distribution around the secondary yolk sac. Being different from the initial sac opening to the midgut lumen [4, 16], the secondary yolk sac membrane is not continuous with the midgut epithelium [17]. This may explain the absence of the omphaloenteric duct in the present specimens. The so-called omphaloenteric duct in humans was most likely to disappear at 4 weeks. However, our observations could not rule out the possibility of “another” connection after 6 weeks between the midgut apex and umbilical cord. We suspected the usual explanation of a famous Meckel’s diverticulum of intestine that should be derived from the initial omphaloenteric duct.
The VA was found to disappear earlier than the VV on the distal side of the midgut apex. We have a series of unpublished data showing a free VV outside of the mesentery even at 9–10 weeks. Actually, a persisting VV has been shown to form a strangulating loop around the intestine [18]. Although tributaries of the VV were not evident in the present specimens, left and right VVs are likely to co-exist. Similarly, because branches of the VA supply the initial yolk sac, multiple VAs are likely to be present even after regression of the sac. These branches, however, seemed not to correspond to multisegmental vitelline VAs of the type often shown in reviews [19] and textbooks [1]. Photographic evaluation of mouse embryos showed the presence of multisegmental arterial roots from the aorta at E 9.5–10.5 [20].
Many recent studies have shown persistent vitelline vessels running along the umbilical cord surface at birth. However, it is difficult to determine whether these thin vessels are actual remnants or develop secondarily as collaterals of the embryonic umbilical vein [19, 21, 22]. These veins in adults, have sometimes been called umbilical veins [23], resulting in serious confusion between embryonic umbilical and vitelline veins. Most of these adult veins seem to correspond to paraumbilical veins [24, 25].
Finally, we considered small numbers of sagittal sections at 5 weeks as the greatest limitation of this study. Although the numbers were limited, the present sagittal sections suggested, within 5 weeks, the yolk sac was likely to exist in the intra-embryonic cavity or even in the dorsal site below the liver and heart, not in the extra-embryonic celom in the ventral side of the future umbilicus. However, the present sagittal sections did not demonstrate the ventral abdomen because of the tissue injury.



 XML Download
XML Download