

 PDF
PDF Citation
Citation Print
Print


INTRODUCTION
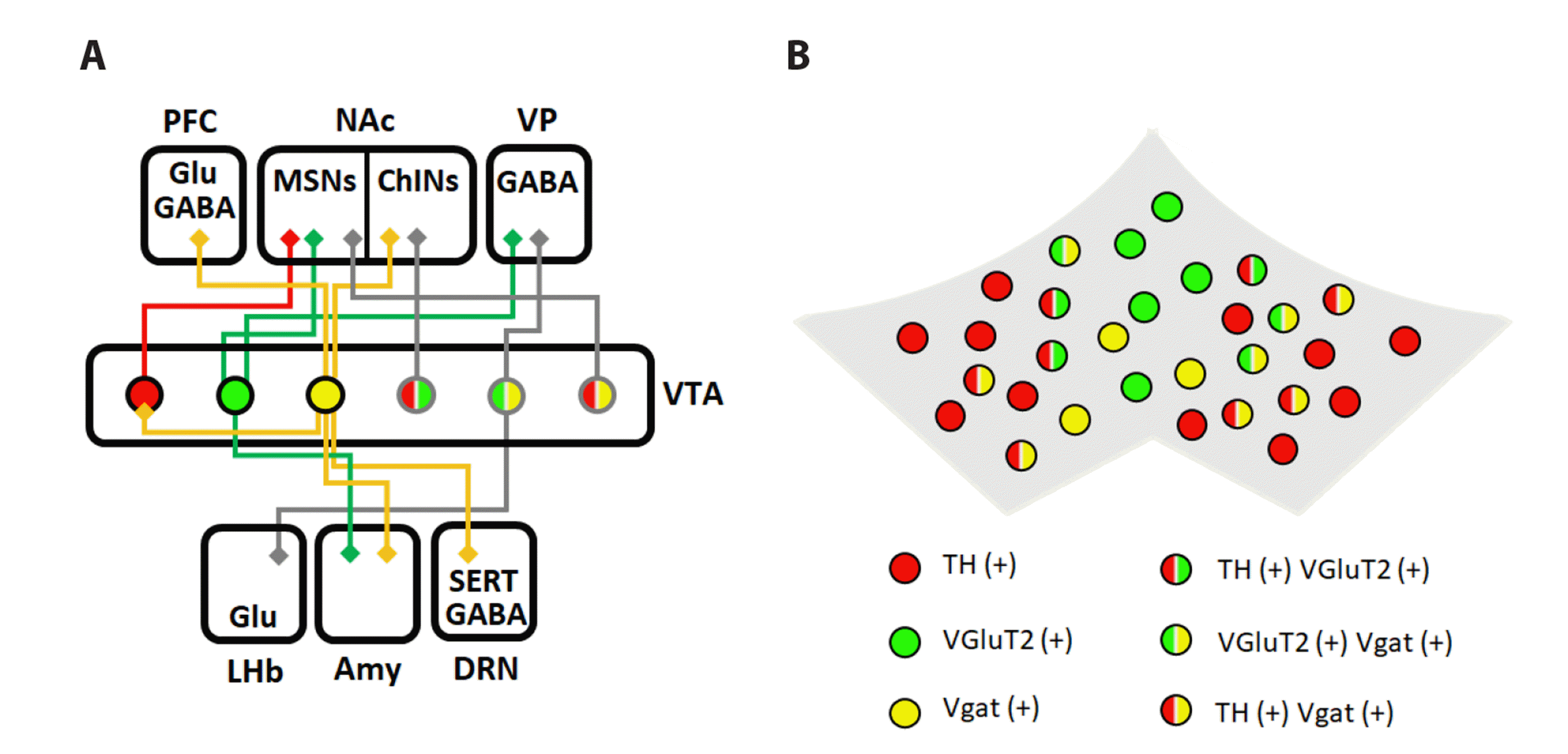
Dopamine transmissions from the midbrain have been characterized to mediate motivated behavior. In the mammalian brain, dopamine is released from ventral tegmental area (VTA) and substantia nigra (SN). Depending on the pre- and post-regions of dopamine, four distinct pathways exist: the mesolimbic, mesocortical, nigrostriatal, and tuberoinfundibular pathways (Fig. 1) [1-3]. The mesolimbic pathway represents dopamine traveling from the VTA to the nucleus accumbens (NAc). It encodes reward-associated memory and reinforcement learning [4]. The mesocortical pathway represents a dopaminergic projection from the VTA to the prefrontal cortex (PFC). This projection is related to cognition and executive behavior [5]. The nigrostriatal pathway is a dopaminergic projection from the SN to the striatum or basal ganglia, which mediates motor control [2,6]. Finally, the tuberoinfundibular pathway releases dopamine from the hypothalamus to the anterior pituitary [3].
Historically, the biological significance of dopamine in reward-seeking behavior has been controversial. There have been two theories regarding the role of dopamine. First, dopamine release reflects motivation [7]; specifically, Pavlovian motivation, as mice show increased movement to get the previously-encountered reward. Zhang et al. [8] first provided a computational model of this theory in 2009. Zhang et al. [8] defined motivation as the integration of the current state and associated cues during reward-associated training. Several studies have reported that the activity of dopamine neurons in the VTA and direct dopamine release at the axon terminal are correlated with motivated behaviors [9-11].
The second theory suggests that dopamine encodes reward prediction error (RPE). The RPE theory originated from value prediction after repetitive training. During the training, mice learn that they can get a positive stimulus, the unconditioned stimulus (US) when they encounter a specific cue, the conditioned stimulus (CS) [12,13]. This theory is supported by the dopamine dynamics in the brain region during exposure to CS and US. A rapid firing of dopaminergic neurons is observed upon exposing mice to an unexpected reward [14]. However, dopamine firing becomes below the basal level when anticipated stimulus is not given [15,16]. Furthermore, dopaminergic neurons show sharp firing to US during the initial learning. In well-trained animals, they often show a higher response to CS than US [17,18]. This finding supports that dopamine dynamics reflect the prediction error.
Whereas dopaminergic pathways and dopamine theories are well-known, non-dopaminergic neurons in the VTA have been underestimated. The contributions of non-dopaminergic, glutamatergic, or GABAergic neurons in the VTA have recently been emphasized regarding motivated behavior. In this article, we review distinct cell populations of the VTA that contribute to motivated behavior through projections and molecular pathways. Moreover, we suggest a role of neurons that release two neurotransmitters compared to that of neurons that release single neurotransmitters.
DOPAMINERGIC NEURONS OF THE VTA
Dopamine neurons account for more than 70% of the total population of VTA neurons. These neurons activate when mice encounter positive stimuli such as food, sugar, water, or addictive drugs [19]. The two different firing patterns for dopamine neurons are phasic and tonic. Well-developed dopamine dynamics in the mammalian brain show that dopamine leaks to the post-region under basal conditions, whereas dopamine release increases after the stimulus [20]. Dopamine-releasing patterns, both phasic and tonic, can change the occupancy of the receptors in post-regions such as the NAc [21].
VTA dopaminergic neurons are highly connected to the ventral striatum [22]. In particular, the projection from the VTA dopaminergic neurons to the medium spiny neurons (MSNs) of the NAc is relevant to motivated behavior. MSNs can be divided into two populations: dopamine receptor subunit 1 expressing MSNs (D1-MSNs) and dopamine receptor subunit 2 expressing MSNs (D2-MSNs) [23]. From the classical perspective, the two different MSN subpopulations have distinct roles [24-26]. D1-MSNs encode reward/positive stimuli and D2-MSNs convey aversive/negative responses [27]. However, both types of MSNs function concurrently in reward-associated behavior [28].
When mice encounter a positive or aversive stimuli, dopamine is released from the axon terminals of the VTA dopaminergic neurons to the dendrites of NAc MSNs [29]. Two types of MSNs respond differentially according to the concentration of dopamine and have contrasting downstream signaling pathways. They display different projections as well [21]. For a classical understanding of projections from D1- and D2-MSNs, D1-MSNs directly inhibit the ventral mesencephalon (VM), which inhibits the thalamus (direct pathway). However, D2-MSNs disinhibit the VM by suppressing the ventral pallidum (VP) (indirect pathway) [27,30]. Distinct pathways have been upended by the finding that D1-MSNs convey an indirect pathway and that D2-MSNs pass through a direct pathway [30]. Based on this novel finding, the optogenetic activation of D1- and D2-MSNs respectively modulate self-stimulation, although the extent differs in each case [31].
Distinct populations of MSNs exhibit different intracellular calcium signaling cascades in response to dopamine [32]. After transient dopamine exposure, inositol 1,4,5-triphosphate (IP3)-mediated calcium signals are activated by D1 receptors (D1Rs), whereas IP3-evoked calcium signals are suppressed by D2 receptors (D2Rs) [33]. To summarize, intracellular calcium activity is enhanced in D1-MSNs but decreased in D2-MSNs, in response to dopamine. This calcium signaling pathway varies according to the concentration of dopamine [32,34]. When MSNs are exposed to high concentrations of dopamine, D1R-MSNs are active through enhancement of the PKA pathway, while D2-MSNs are inactive. On the other hand, once dopamine concentration is low, D2-MSNs become active through adenosine A2A receptor (A2AR) activation, which increases intracellular calcium levels [34].
GLUTAMATERGIC NEURONS OF VTA
Recent research suggests that VTA glutamatergic neurons also modulate motivated behavior [35,36]. Approximately 30% of VTA neurons are type 2 vesicular glutamate transporter (VGLUT2)-positive neurons [37,38]. They are specifically localized in the medial section of the VTA, such as the parabrachial pigmented nucleus and the interfascicular nucleus. Interestingly, the current study has shown that optogenetic activation of VTA glutamatergic neurons is sufficient to induce reward reinforcement behaviors [39]. Conditional knockout of VTA glutamatergic neurons causes reduced motivated behavior. Moreover, the fact that VTA VGLUT2 neurons become active during self-administration, drug-seeking behaviors, and reinforcement has been discovered [39-41]. VGLUT2-positive neurons in the VTA project to the NAc medial shell, VP, and lateral habenula (LHb). VTA VGLUT2-positive neurons excite NAc and VP neurons directly, whereas they suppress LHb neurons [42]. Once glutamate is released in the axon terminal, it binds to N-methyl-D-aspartate receptors (NMDA-Rs), and leads to calcium influx, which activates the pERK signaling pathway [43].
VTA VGLUT2 neurons are differentiated into two types: the VGLUT2-only-expressing neurons (VGLUT only neurons) and both VGLUT2 and dopamine-expressing neurons (VGLUT2-DA neurons) [44]. VGLUT2-DA neurons co-release glutamate and dopamine at the axon terminals. These neurons release glutamate through asymmetric synapses and dopamine through symmetric synapses. This new finding is contradictory to the classical viewpoint of Dale’s principle, which states that a single neuron releases a single type of neurotransmitter. VGLUT2-DA neurons increase the survivability and axonal arborization of VTA dopamine neurons [45,46]. In addition, this population projects to the cholinergic neurons in the NAc medial shell, and contributes to the switching of behavioral response during cued reinforcement tasks [38,45]. However, the studies regarding VGLUT2-DA neurons cannot suggest how these neurons act differentially from VGLUT2-only or DA-only neurons.
VTA VGLUT2 neurons also co-release the inhibitory neurotransmitter, GABA [47]. VGLUT2-GABA neurons are distinct from those of VGLUT-only neurons at the circuit level. VGLUT2-GABA neurons are highly connected to the LHb, whereas most VTA VGLUT2 neurons project to the NAc shell [42,48]. These two populations exhibit different activities for specific behaviors. In classical conditioning, VGLUT2-GABA neurons become active during exposure to a reward or electric shock. VGLUT2 only neurons are also active in response to the reward and electric shock. However, only this population shows active firing in response to the conditioned stimulus associated with the reward (CS+) [48]. To summarize, VGLUT2-GABA neurons encode the valence itself.
GABAERGIC NEURONS OF VTA
The GABAergic neurons form the largest population of VTA accounting for approximately 30% of the total VTA neuronal population [49]. Efferents of VTA GABAergic neurons project to the NAc, PFC, central amygdala, LHb, and dorsal raphe nucleus (DRN). The molecular functions of GABAergic synapses are distinct according to the type of GABA receptors. GABAA receptors cause hyperpolarization of post-neurons by the influx of chloride ions [50]. In contrast, GABAB receptors also induce hyperpolarization by suppressing acetyl cyclase and voltage-gated calcium channels (VGCC) [51].
GABAergic neurons have two distinct roles: inhibition of dopaminergic neurons in the VTA and inhibition of distal brain regions [49]. The function of VTA GABAergic neurons is quite complicated. Previous findings on the role of VTA GABAergic neurons have focused on the aversion and interruption of reward [52,53]. These experimental results imply that the activation of VTA GABAergic neurons activates aversion behavior and inactivates reward-consuming behavior [49]. However, VTA GABAergic neurons play a significant role in cue responsiveness and reward-associated learning [53,54]. VTA GABAergic neurons become active and suppress VTA DA neurons when mice expect a reward, such as sucrose or cocaine after they are exposed to a stimulus that is not associated with the reward [48,55,56]. This means that VTA GABAergic neurons interrupt reward consumption and disruption of responsiveness by gating dopaminergic activity in the VTA [57]. Moreover, mice show specific place preference, if the projection from caudal VTA GABAergic neurons to DRN serotonergic neurons is activated [58].
GABAergic neurons in the VTA also co-release dopamine or glutamate (GABA-DA neurons or VGLUT2-GABA neurons). GABA-dopamine co-releasing neurons highly target the NAc, whereas VGLUT2-GABA neurons project to the LHb and VP [49]. Interestingly, neurotransmitter release from these neurons is mediated by the uptake mechanism instead of the classical GAD1/2 pathway [59]. VTA DA neurons synthesize GABA through aldehyde dehydrogenase 1A1 [60]. GABA vesicle loading is mediated by a vesicular monoamine transporter instead of the vesicular GABA transporter (VGAT) [61]. However, the environmental stimuli that can activate GABA-DA neurons remain unknown.
CONCLUSIONS AND PERSPECTIVES
This review summarizes that distinct populations of the VTA, which participate in motivated behavior by extending the classical view of the dopamine-mediated reward system. There are three different cell types, dopaminergic, glutamatergic, and GABAergic neuronal populations in the VTA. Each population affects motivated behavior through distinct mechanisms. Dopaminergic neurons project to the MSNs in the NAc. Released dopamine causes opposite downstream signaling, according to the type of dopaminergic receptors expressed in MSNs. VTA glutamatergic neurons also project to MSNs of the NAc shell and encode a reward-predicting cue (CS+). VTA GABAergic neurons mediate motivated behavior by suppressing VTA dopaminergic neurons or modulating distal regions, such as the LHb and DRN.
However, these three populations are not distinct. VGLUT2-DA, GABA-DA, and VGLUT2-GABA neurons exist, and their functions compared to those of single-expressing neurons are unclear. In accordance with current studies on the function of VTA neurons, they play a central role in motivated behavior as well as aversive experience [62-64]. Further research is required to understand how the functions of VTA neurons that release two other neurotransmitters are different from those of single-expressing neurons.
 XML Download
XML Download