

 PDF
PDF Citation
Citation Print
Print


Introduction
Bone formation and maintenance are regulated by bone forming cells, osteoblasts, and bone resorbing cells, osteoclasts (1). An imbalance between osteoblasts and osteoclasts leads to bone diseases or disorders such as osteoporosis, osteopetrosis, and Paget’s disease (1-3). Osteoblasts are bone forming cells that regulate mineralization and synthesis of the bone matrix. Osteoblasts are important regulators in osteoclast differentiation and function (4). Osteogenic differentiation, which plays an important role in osteoblast generation, is a useful therapeutic target for treating bone diseases (5).
Mesenchymal stem cells (MSCs) exist in a variety of tissues, including adipose tissue, skeletal muscle, bone marrow, and the umbilical cord. MSCs have the capacity for self-renewal and differentiate into various cell types including adipocytes, osteoblasts, and chondrocytes. Recently, Adipose-Derived Stromal Cells (ADSCs) have been applied in regenerative medicine to compensate the bone loss in various diseases (6-8). Osteogenesis from MSC is a complex process that is regulated by various stimuli, involving signaling pathways and diverse transcription factors (9).
Horses are valuable in the fields of recreation and sports, and as companions (10, 11). Equine health management is a high-value industry, and bone, muscle, and tendon injuries in particular can be expensive to treat (10-12). Injured cartilage and ligaments, bone loss, and bone fractures are common problems in horses (12, 13). Equine adipose-derived stromal cells (eADSCs) are isolated from adipose tissue, and can be differentiated into osteoblasts, myocytes, chondrocytes, and adipocytes (6, 8). Also, eADSCs are being studied for the treatment of horses, including surface digital flexor tendon (SDFT) damage of horses (14-17).
Flavonoids are found in fruits and vegetables, and are widely consumed by humans (18). Flavonoids have diphenyl propane (C6C3C6) skeleton, and the patterns of hydroxylation in the B ring play an important role in their function (19). Flavonoids have anti-cancer, anti-oxidant, and anti-inflammatory properties (19-23). We have recently verified that flavonoid treatment with 3, 2’-dihydroxyflavone (3, 2’-DHF) up-regulated cell growth and stemness marker expression in embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs). We also demonstrated that 3, 2’-DHF-treated iPSCs promote functional recovery and regeneration when transplanted into rat nerves, which is associated with their neuroprotective properties (24). We also found that 3, 4’-DHF has anti-oxidant, anti-apoptotic, and cell differentiation regulatory properties (25).
Here, we found that treatment with 3, 4’-dihydroxyflavone (3, 4’-DHF) promoted osteogenic differentiation from eADSCs by regulating the Reactive Oxygen Species (ROS). Furthermore, co-treatment with 3, 4’-DHF and U0126 or N-acetyl cysteine (NAC) regulated osteogenesis in eADSCs. We propose that 3, 4’-DHF may be used to regulate osteogenesis for therapeutic applications in humans and animals.
Go to : 
Materials and Methods
Chemicals and antibodies
3, 4’-DHF was purchased from Indofine Chemical Inc. (Hillsborough, NJ, USA) and dissolved in dimethyl sulfoxide (DMSO, Sigma Aldrich, St. Louis, MO, USA). The ROS scavenger, NAC, and the MEK kinase inhibitor, U0126, were obtained from Calbiochem (San Diego, CA, USA). Dexamethasone, β-glycerophosphate, ascorbic acid, alkaline phosphatase kits, and alizarin red s were purchased from Sigma-Aldrich (St. Louis, MO, USA). The reagent for measuring intracellular ROS, 2’, 7’-dichlorodi-hydrofluorescein diacetate (H2DCFDA), was obtained from Molecular Probes (Eugene, OR, USA). Primary antibodies for β-actin, phospho-ERK, ERK, phospho-AKT, and AKT were purchased from Santa Cruz Biotechnology (Dallas, TX, USA), and the peroxidase conjugated secondary antibodies, anti-mouse IgG, anti-rabbit IgG, anti-goat IgG, were obtained from Amersham Bioscience (Piscataway, NJ, USA).
Cell culture and osteogenic differentiation
eADSCs were provided by Kyungpook National University (Daegu, Korea) by Prof. Jeong. eADSCs were cultured in Dulbecco’s modified Eagle’s medium (DMEM; Invitrogen, Carlsbad, CA, USA), supplemented with 10% Fetal Bovine Serum (FBS; GE Healthcare, Chicago, IL, USA) and 0.1% penicillin-streptomycin (Sigma Aldrich, St. Louis, MO, USA). Osteogenesis was induced by osteogenic differentia-tion media containing 0.1 µM dexamethasone (Sigma Aldrich, St. Louis, MO, USA), 10 mM β-glycerophosphate (Sigma-Aldrich, St. Louis, MO, USA), and 50 μg/ml ascorbic acid-2-phosphate (Sigma-Aldrich, St. Louis, MO, USA) for 2 weeks.
Alizarin red S staining
The cells were fixed with 4% paraformaldehyde for 20 min and stained with 2% alizarin red s solution for 10 min. Stained cells were rinsed twice with distilled water and once with Dulbecco’s Phosphate-Buffered Saline (DPBS). Staining was confirmed using a light microscope. The Alizarin red stain was then removed in 10% acetic acid for 30 min and neutralized with 10% ammonium hydroxide. The absorbance was measured at 405 nm using an xMarkTM Microplate Absorbance Spectrophotometer (Bio-Rad Laboratories, Hercules, CA, USA).
Detection of intracellular ROS
Intracellular ROS was measured using H2DCFDA (Molecular Probes, Eugene, OR, USA). Cells were scraped and centrifuged at 3,000 rpm for 1 min. Then cell pellets were washed with DPBS, and stained in the dark with 10 μM H2DCFDA for 30 min. H2DCFDA fluorescence was detected using a flow cytometer (FACS Calibur, Becton Dickinson, Heidelberg, Germany), and data were analyzed using Cell Quest pro software 5.
RNA isolation, cDNA synthesis, and real time RT-PCR analysis
Total RNA was isolated from eADSCs at 4-time points (2, 5, 8, 11 days) during osteogenic induction using Trizol (Sigma-Aldrich, St. Louis, MO, USA). Reverse transcrip-tion was carried out with 2 μg of total RNA using M-MLV reverse transcriptase (Promega, Madison, WI, USA), and quantitative PCR was performed using Fast SYBR Green Master Mix (Applied Biosystems, Stockholm, Sweden). Primers were designed using Primer3 (ver. 4.0) as follows: Equine ALP, F 5’-GACAAGAAGCCCTTCACTGC-3’, R 5’-TGGGGGATGTAGTTCTGCTC-3’; equine OCN, F 5’-GTGCAGAGTCTGGCAGAGGT-3’, R 5’-TCGTCACAGT CTGGGTTGAG-3’, equine OPN, F 5’-CCATGAGGATGA CCAGGACT-3’, R 5’-AACGTCGGTGGAAAAATCAG-3’; equine RUNX2, F 5’-TTACTTACACCCCGCCAGTC-3’, R 5’-GCAGCATTCTGGAAGGAGAC-3’; equine SOD1, F 5’-GATTCCACGTCCACGAGTTT-3’, R 5’-ATGCTTTCC CGAGAGTGAGA-3’; equine SOD2, F 5’-CCCCGACTTG CAGTATGATT-3’, R 5’-TGCAGAGCAATCTGAGCTGT-3’; equine Catalase, F 5’-TACCCGTGAACTGTCCCTTC-3’, R 5’-GGAGAGCACTGGCTTTTACG-3’; equine NOX1, F 5’-TGATCGCAAGCTCAAAACAC-3’, R 5’-AGGATGTCA GTGGCCTTGTC-3’; equine NOX4, F 5’-TTTAGACACC CACCCTCCTG-3’, R 5’-CAGAAAGCCAAAGCCAAGTC-3’, and equine GAPDH, F 5’-ATCACTGCCACCCAGAAGAC-3’, R 5’-GTGAGCTTCCCATTCAGCTC-3’.
Western blotting
Control and differentiated eADSCs were scraped and washed with DPBS. Cell pellets were resuspended in ice-cold lysis buffer, containing 10% glycerol (Junsei Chemical, Tokyo, Japan), 100 mM Tris–HCl pH 7.5 (Sigma-Aldrich, St. Louis, MO, USA), 0,1% Triton X-100 (Amresco, CV, USA), 10 mM NaCl (Sigma-Aldrich, St. Louis, MO, USA), 1 mM sodium orthovanadate (Sigma-Aldrich, St. Louis, MO, USA), 50 mM sodium fluoride (Sigma-Aldrich, St. Louis, MO, USA), 1 mM phenylmethylsulfonyl fluoride (Sigma-Aldrich, St. Louis, MO, USA) and 1 mM p-nitrophenyl phosphate (Sigma-Aldrich, St. Louis, MO, USA). The protein concentration was determined using the Bradford assay reagent (Bio-Rad Laboratories, Hercules, CA, USA). An equal amount of each protein sample was separated on a 10% SDS PAGE gel. Proteins were then electrophoretically transferred onto a nitrocellulose membrane (Whatman International Limited, Kent, UK) and blocked with 5% non-fat milk powder (Amresco, Solon, OH, USA) dissolved in Tris-buffered saline (Sigma-Aldrich, St. Louis, MO, USA). Proteins were detected using the appropriate primary and secondary antibodies, and enhanced chemiluminescence (ECL; Amersham Bioscience, Piscataway, NJ, USA).
Statistical analysis
All experiments were repeated at least three times, and data are presented as the mean±standard deviation (±SD). All statistical comparisons were performed using ANOVA (analysis of variance) or Student’s t-test in MS Excel 2013 (Microsoft, Redmond, WA, USA). Differences between groups were considered to be statistically significant at *p< 0.05.
Go to :
Results
Effects of 3, 4’-DHF on eADSCs during osteogenic differentiation
To investigate effect of 3, 4’-DHF, we treated 3, 4’-DHF during osteogenic differentiation for 2 weeks in eADSCs. After osteogenic differentiation, we analyzed ALP activity, a well-known osteogenesis marker in differentiated eADSCs. ALP activity was significantly increased in eADSCs differentiated with 3, 4’-DHF than in the control (Fig. 1A). In addition, we assessed the extracellular matrix mineralization using alizarin red S staining, and found that 3, 4’-DHF-treated eADSCs had higher amounts of alizarin red S staining compared to that of the control group (Fig. 1B). Next, we measured the calcium content in eADSCs and 3, 4’-DHF exposed-eADSCs during osteogenic differentiation and confirmed that the calcium level also increased in 3, 4’-DHF-treated eADSCs (Fig. 1C). The expression of Osteocalcin (OCN), Osteopontin (OPN), RUNX2, and ALP, which are osteogenic differentiation markers, increased in differentiated eADSCs treated with 3, 4’-DHF compared to that of the control (Fig. 1D).
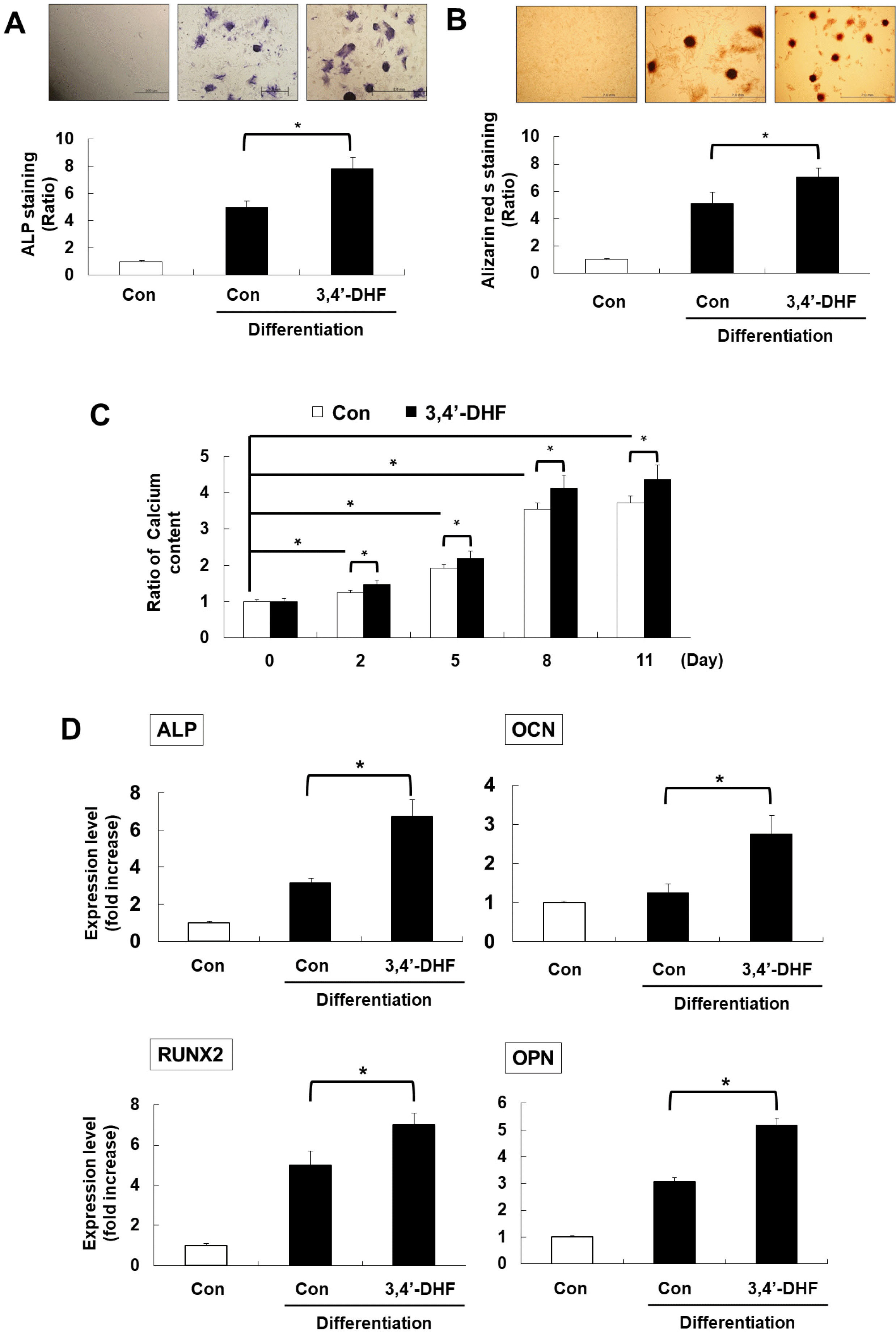 | Fig. 13, 4-’dihydroxyflavone (3, 4’-DHF) enhanced osteogenesis in equine Adipose-Derived Stromal Cells (eADSCs). (A, B) Osteogenic differentiation marker staining of 3, 4’-DHF-treated eADSCs with osteogenic differentiation at day 14 (A) Alkaline phosphatase (ALP) and (B) Alizarin red s. (C) Calcium content ration of 3, 4’-DHF-treated eADSCs with osteogenic differentiation. (D) qRT-PCR analysis of osteogenesis markers (Osteocalcin (OCN), Osteopontin (OPN), RUNX2, and ALP) in eADSCs and 3, 4’-DHF eADSCs. Error bars represent±SD from at least three independent experiments (*p<0.05).
|
ERK phosphorylation increased with 3, 4’-DHF treatment during osteogenic differentiation
AKT and ERK signaling activated during osteogenic differentiation in MSCs. So, we investigated the phosphorylation level of AKT and ERK proteins during osteogenic differentiation with or without 3, 4’-DHF treatment. The phosphorylation level of AKT and ERK increased upon osteogenic differentiation (Fig. 2A). However, AKT phosphorylation was a slightly higher than control differentiation group in 3, 4’-DHF treated eADSCs. In contrast, ERK was significantly phosphorylated in 3, 4’-DHF treated eADSCs during osteogenic differentiation (Fig. 2A). To confirm that ERK is associated with osteogenic differentiation of eADSCs, we treated PD98059, an ERK inhibitor, in eADSCs and 3, 4’-DHF-treated eADSCs. We confirmed that phosphorylated ERK levels are decreased by treatment of PD98059 in eADSCs and 3, 4’-DHF-treated eADSCs (Fig. 2B). Next, we measured the expression of osteogenic differentiation marker genes, ALP and OPN, in eADSCs and 3, 4’-DHF-treated eADSCs with PD98059 (Fig. 2C). Pre-treatment with PD98059 resulted in reduced expression levels in osteogenic related genes in eADSCs and 3, 4’-DHF-treated eADSCs.
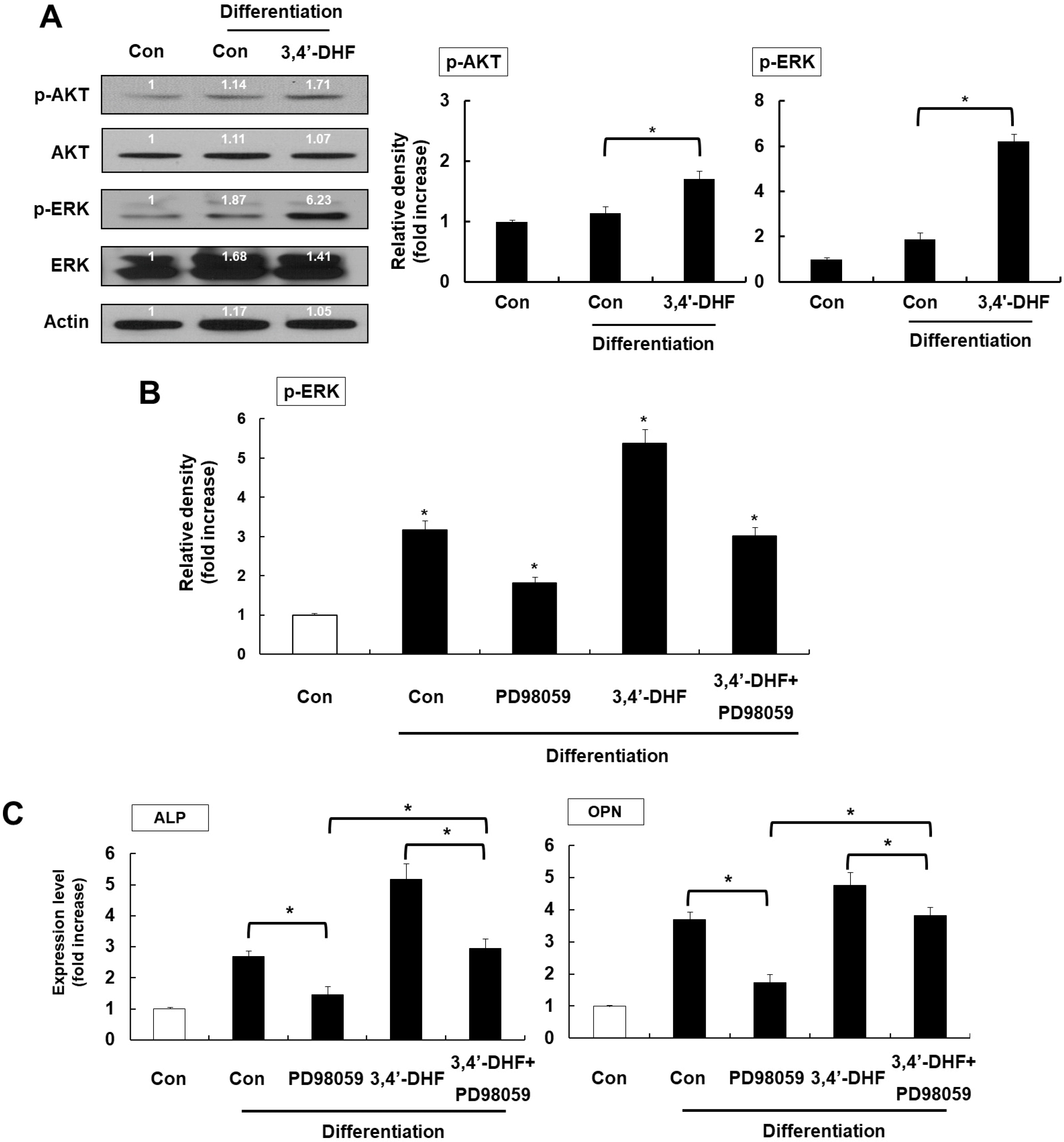 | Fig. 23, 4’-DHF induced ERK activation during osteogenic differentiation. (A) Western blot analysis of ERK and AKT phosphorylation during osteogenic differentiation in the presence or absence of 3, 4’-DHF. (B) Expression level of phosphorylated ERK in the presence or absence of PD98059 or 3, 4’-DHF. (C) qRT-PCR analysis of osteogenic differentiation markers ALP and OPN in eADSCs with or without 3, 4’-DHF-treatment in the presence or absence of PD98059. Each experiment was repeated in triplicate and data are presented as means±standard deviation (p<0.05, denoted by*).
|
U0126 induces osteogenic differentiation via BMP signaling despite EKR inactivation in eADSCs
To confirm ERK inactivation repress osteogenic differentiation in eADSCs, we treated U0126, another ERK inhibitor, during osteogenic differentiation. Surprisingly, U0126 showed slightly increased osteogenic marker expression although inhibited ERK phosphorylation (Fig. 3A and 3B). Recent studies have demonstrated that U0126 increases expression of osteogenic-associated genes by activating BMP signaling pathway transduction despite reduced ERK phosphorylation (26). Similarly, in eADSCs, treatment with U0126 has been shown to induce differentiation through activation of the BMP signaling pathway (Fig. 3C). Therefore, treatment of 3, 4’-DHF during osteogenic differentiation in eADSC induces ERK phosphorylation, but ERK phosphorylation does not induce differentiation absolutely. And U0126 increases osteogenic differentiation through BMP signaling despite inhibiting ERK phosphorylation.
 | Fig. 3Treatment with the ERK inhibitor U0126, led to an increase in osteogenesis in eADSCs and 3, 4’-DHF eADSCs via BMP signaling. (A) Western blot analysis of phosphorylated ERK in eADSCs and 3, 4’-DHF eADSCs in the presence or absence of U0126. (B) qRT-PCR of osteogenesis marker gene expression in eADSCs and 3, 4’-DHF eADSCs treated with U0126. (C) qRT-PCR analysis of BMP2 and BMP4 gene expression in the presence or absence of U0126 and PD98059 in eADSCs and 3, 4’-DHF eADSCs. Error bars represent±SD from at least three independent experiments (*p<0.05).
|
3, 4’-DHF regulates osteogenic differentiation by modulation of ROS signaling
Previously, we demonstrated that 3, 4’-DHF inhibit adipogenic differentiation by modulation of ROS signaling in eADSCs (27). So, we measured the intracellular ROS levels in osteogenic differentiated eADSCs and in 3, 4’-DHF-treated eADSCs. During osteogenesis, 3, 4’-DHF-treated eADSCs further decreased H2DCFDA fluorescence intensity than control differentiation groups (Fig. 4A). To investigate the role of ROS, we pre-treated both eADSCs and 3, 4’-DHF-treated eADSCs with a ROS scavenger, NAC. Pre-treatment with NAC suppressed ROS generation during osteogenic differentiation (Fig. 4B). Moreover, NAC or 3, 4’-DHF treatment significantly elevated the expression of osteogenesis markers (Fig. 4C). We also measured the expression of ROS-related genes in NAC or 3, 4’-DHF treated eADSCs. We found that NAC or 3, 4’-DHF modulate the expression of ROS-related genes, including SOD1 (Cu/Zn superoxide dismutase), SOD2 (Mn superoxide dismutase), NOX1 (superoxide-generating NADPH oxidase enzymes), NOX, and catalase (hydrogen peroxide-decomposing enzyme) (Fig. 4D). Interestingly, SOD2 and catalase expression increased, while NOX1 and NOX4 expression decreased slightly after treatment with NAC or 3, 4’-DHF. SOD1 expression increased immediately after treatment with NAC. In summary, 3, 4’-DHF treatment resulted in ROS modulation via differential regulation of the expression levels of ROS-related genes during osteogenic differentiation.
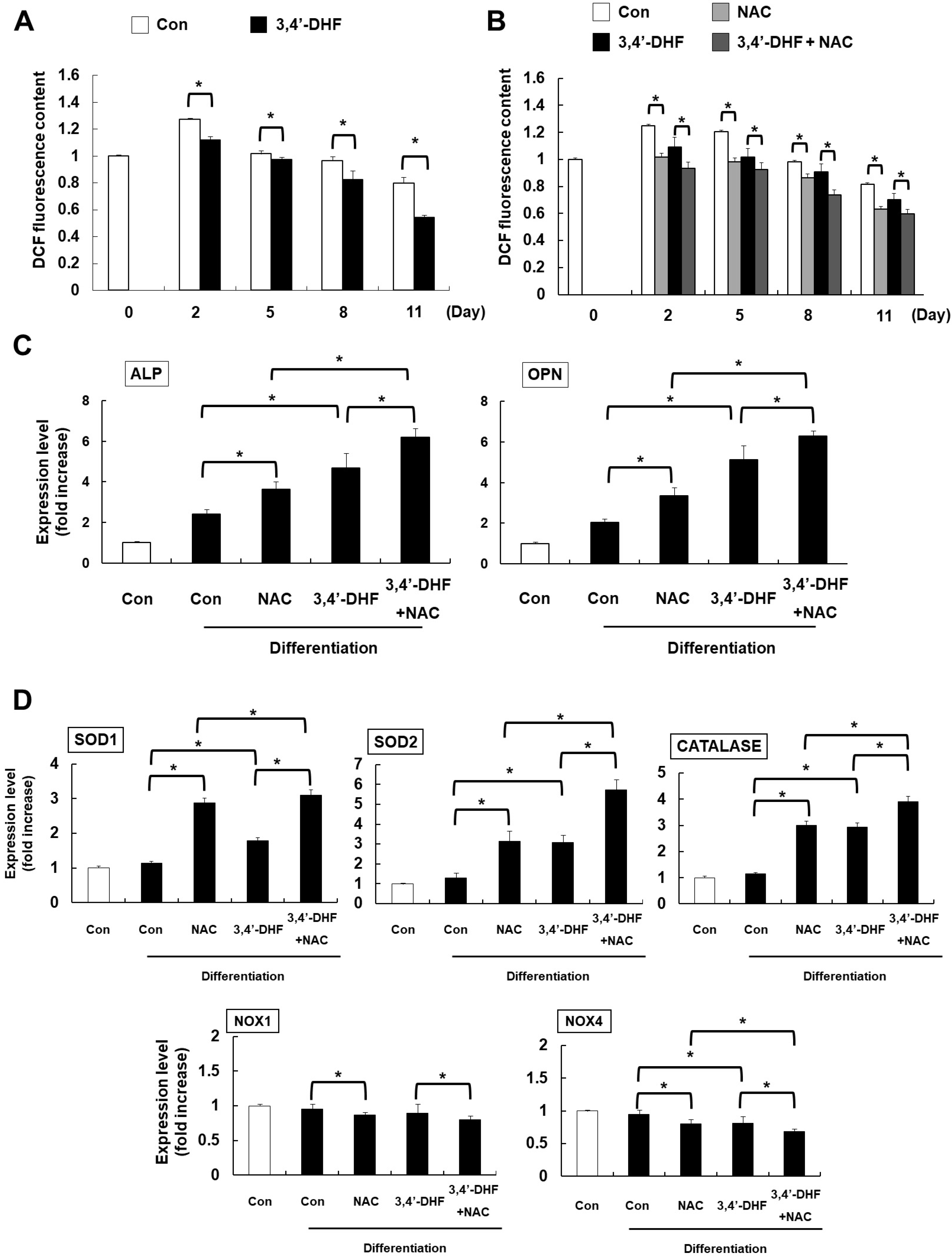 | Fig. 43, 4’-DHF regulates osteogenic differentiation by modulation of Reactive Oxygen Species (ROS) signaling. (A) Intracellular ROS level according to 2’, 7’-dichlorodihydrofluorescein diacetate (H2DCFDA) fluorescence by flow cytometry. (B) The intensity of H2DCFDA fluorescence in N-acetyl cysteine (NAC) treated eADSCs and 3, 4’-DHF eADSCs. (C) Expression of osteogenesis marker genes in NAC treated or untreated eADSCs and 3, 4’-DHF eADSCs according to qRT-PCR analysis. (D) Expression of ROS-related genes in eADSCs and 3, 4ʹ-DHF eADSCs in the presence or absence of NAC treatment. Error bars represent±SD from the mean of three independent experiments (*p<0.05).
|
Co-treatments 3, 4’-DHF, U0126 and NAC enhances osteogenic differentiation in eADSCs
We investigated the effect of U0126 on the intracellular ROS level in eADSCs and 3, 4’-DHF-treated eADSCs during osteogenic differentiation (Fig. 5A). Inhibition of ERK activity by U0126 treatment did not result in a significant difference in ROS generation in eADSCs and 3, 4’-DHF-treated eADSCs during osteogenic differentiation. To characterize the influence of both of ROS and ERK signaling on the osteogenic differentiation of eADSCs and 3, 4’-DHF-treated eADSCs, we treated the cells with NAC and U0126. Intracellular ROS levels decreased in NAC-treated cells, while U0126 had no effect on intracellular ROS levels (Fig. 5B). We previously observed that the expression level of osteogenic differentiation markers increased during treatment with either 3, 4’-DHF, U0126, or NAC. Co-treatment with 3, 4’-DHF, U0126, and/or NAC led to larger increases in the expression levels of osteogenic differentiation markers than treatment with individual compounds (Fig. 5C). We illustrated our results in one representative schematic that shows the link between ERK and ROS signaling during 3, 4’-DHF-enhanced osteogenic expression, and their effects on the expression levels of osteogenic differentiation markers (Fig. 6). Combined treatment with 3, 4’-DHF, U0126, and/or NAC resulted in additive enhancement of the expression of osteogenic differentiation markers.
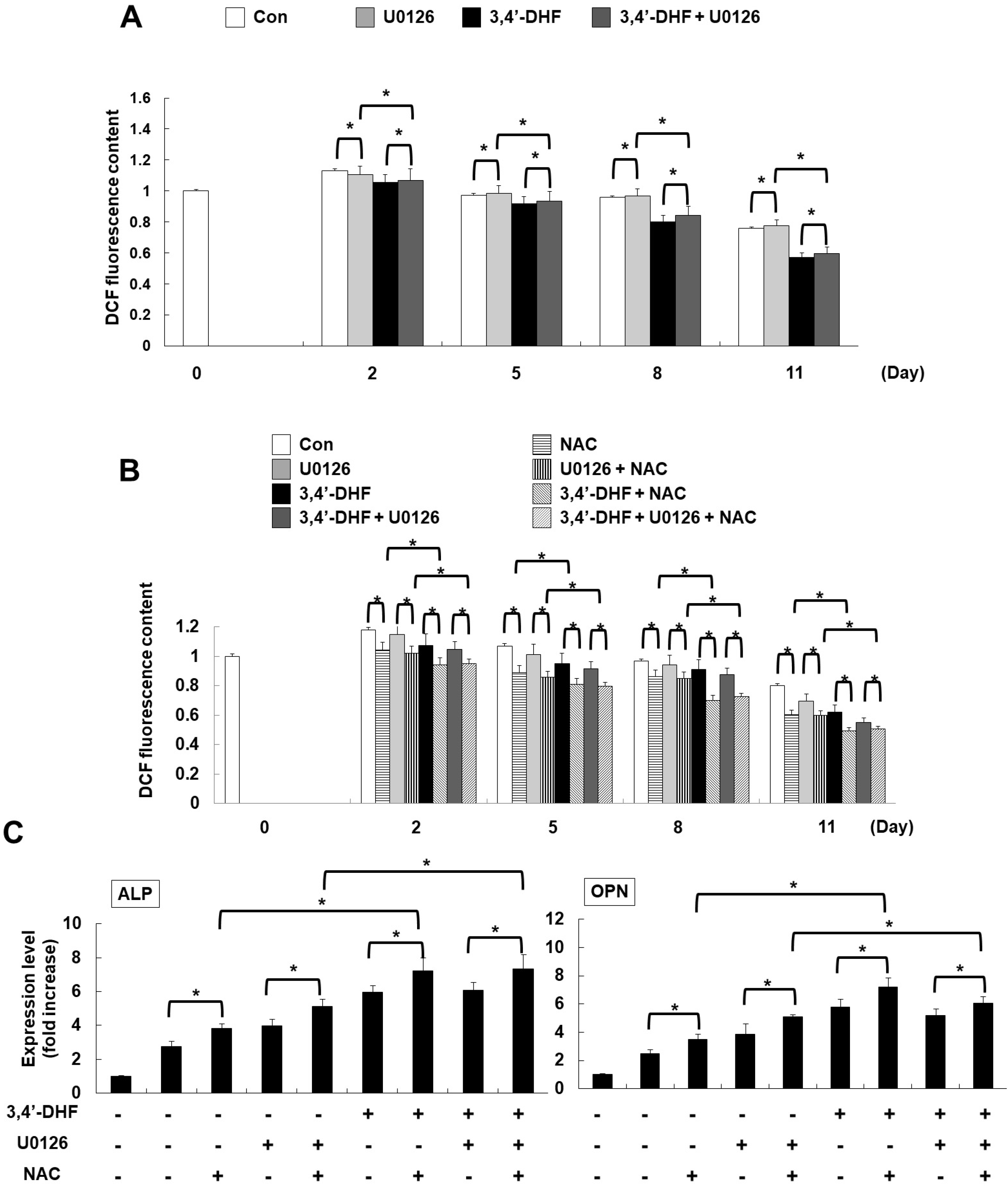 | Fig. 5Co-treatment with 3, 4’-DHF, U0126, and NAC regulated osteogenic differentiation. (A) ROS levels were determined by measuring H2DCFDA fluorescence using a flow cytometer, with or without U0126 treatment in eADSCs or 3, 4’-DHF-treated eADSCs during osteogenic differentiation. (B) Intracellular ROS levels were measured following treatment with U0126 or NAC in eADSCs and 3, 4’-DHF eADSCs. (C) qRT-PCR analysis of osteogenic differentiation marker gene expressions in eADSCs and 3, 4’-DHF eADSCs, in the presence or absence with U0126 or NAC. Each experiment was repeated in triplicate and data are presented as means±SD (*p<0.05).
|
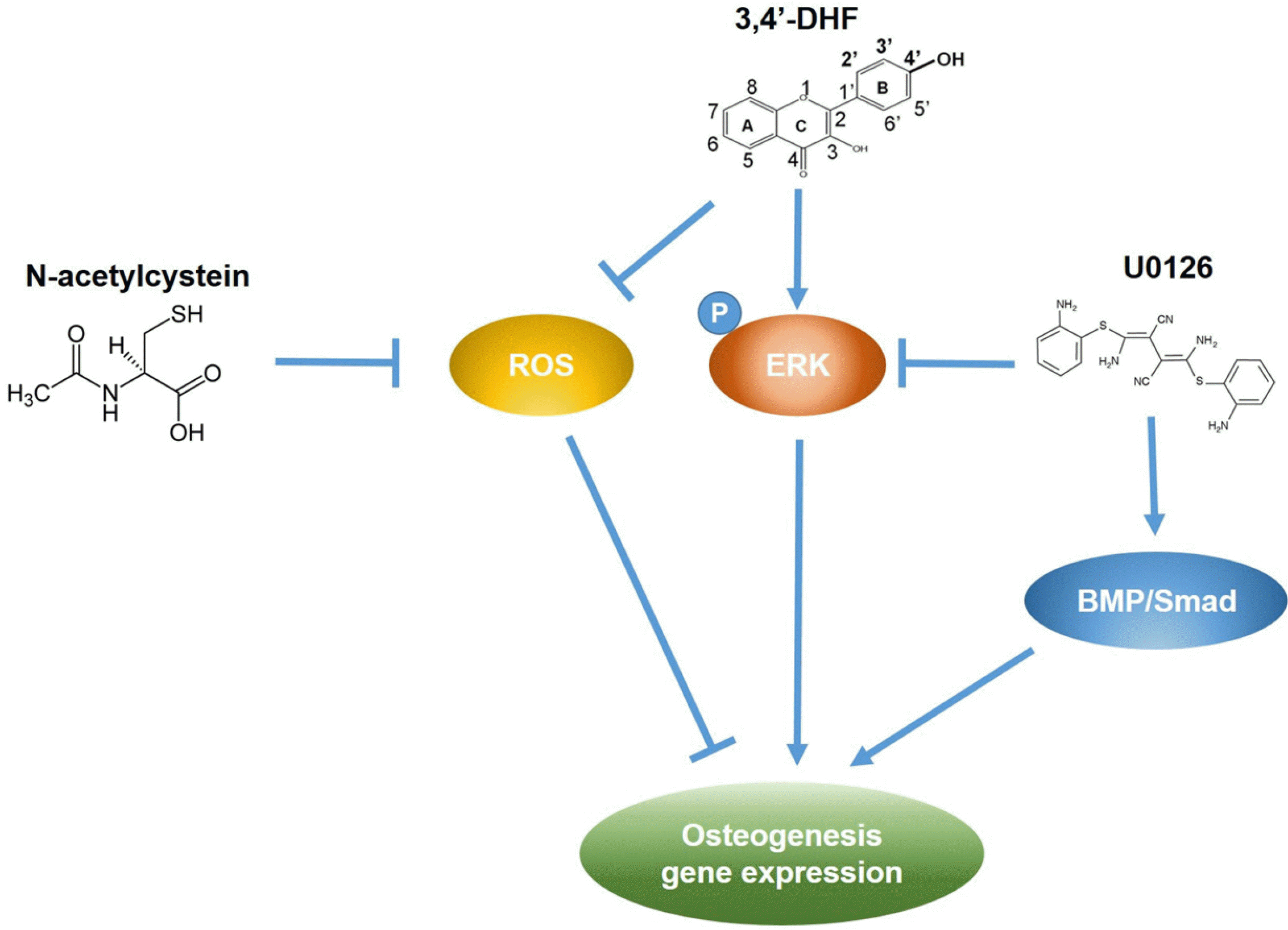
Go to :
Discussion
Flavonoids form the largest group of plant phenols and are found in fruits, vegetables, bark, roots, and grains (28). Flavonoids play a role in controlling various biological and pharmacological activities such as antioxidant, anti-inflammation, antiviral properties (24). Antioxidant activity is the most common function of flavonoids and regulates cell function and differentiation (29). The beneficial function of flavonoids is increasingly used for the prevention and treatment of infections, allergic, inflammatory, and degenerative diseases. In the future, there will be increased interest in the development and application of flavonoid-related drugs, and it can be effectively used for treating diseases. Treatment with 3, 4’-DHF resulted in increased osteogenic differentiation of eADSCs. The inverse relationship between adipogenesis and osteogenesis in ADSCs has been previously reported (9, 30). Osteogenesis is an important process for osteoblast generation, which can be used to treat bone metabolic diseases. Bone contains two distinct cell lineages: osteoblasts and osteoclasts, which are responsible for bone generation and resorption, respectively. Bone maintenance is regulated by these cell lineages, and imbalances between them leads to bone metabolic diseases such as osteoporosis. Osteoporosis is a disorder characterized by a decrease in bone density and bone mass caused by osteoclasts. As osteogenesis plays an important role in osteoblast generation and bone formation, many studies have suggested that osteogenesis is a useful therapeutic strategy for treating osteoporosis (5). Several studies have revealed the relationship between flavonoids and bone development and demonstrated the potential of flavonoids for the treatment of osteoporosis (31, 32). Some flavonoids including genistein, daidzein, icariin, quercetin, rutin, luteolin, kaempferol and naringin, have been investigated for their potential use in the prevention and treatment of osteoporosis. The antioxidant, anti-inflammatory, and bone-conserving properties of flavonoids have recently confirmed to help prevent age-related bone loss and osteoporosis (33).
During osteogenic differentiation, ERK phosphorylation increases (26). IGF-I, EGF, and FGF can regulate osteogenesis via the ERK signaling pathway (34-36). In eADSCs, we observed that ERK activated during osteogenic differentiation. ERK phosphorylation also signifi-cantly increased in the presence of 3, 4’-DHF. Pre-treatment with ERK pathway inhibitors, PD98059 and U0126, significantly suppressed ERK phosphorylation during osteogenic differentiation (Figs. 2 and 3). PD98059 treatment of eADSCs and 3, 4’-DHF-treated eADSCs reduced the amount of ERK phosphorylation levels and the expression of osteogenic differentiation marker genes (Fig. 2). In contrast, although U0126 reduced ERK phosphorylation levels, led to an increase in osteogenic differentiation via BMP signaling pathway (Fig. 3) (26).
Oxidative stress is caused by high levels of ROS production and is associated with apoptosis and damage in a variety of pathological conditions, including aging, neurodegeneration, cancer, and osteoporosis. ROS affect DNA and proteins, cell proliferation, metabolism, and differentiation regulation (1, 37). Previous studies have confirmed that the regulation of ROS production is important for osteogenic differentiation (37). During osteogenesis, ROS levels are down-regulated in a time-dependent manner (Fig. 4A). In the present study, we have shown that ROS levels decreased slightly during osteogenic differentiation, and in cells treated with the ROS inhibitor NAC, we observed an increase in the expression of osteogenic differentiation marker genes (Fig. 4C). Treatment with 3, 4’-DHF resulted in a decrease in intracellular ROS levels during osteogenesis. We also investigated the expression level of the ROS-regulating genes SOD1, SOD2, NOX1, NOX4, and catalase in eADSCs, and found that treatment with NAC or 3, 4’-DHF resulted in differential regulation of gene expression (Fig. 4D), indicating that 3, 4’-DHF modulates ROS-regulating gene expression in eADSCs. When pre-treating with the ERK pathway inhibitor U0126, 3, 4’-DHF-mediated regulation of intracellular ROS generation plays an important role in the 3, 4’-DHF-specific modulation of ERK pathway during osteogenic differentiation. The differential regulation of the ROS-ERK signaling pathway therefore plays an important role in 3, 4’-DHF-mediated differential modulation of osteogenic differentiation in eADSCs (Fig. 5). We investigated the expression of osteogenesis markers during co-treatment of U0126 and NAC with 3, 4’-DHF. We confirmed that 3, 4’-DHF, U0126, and NAC increase the expression of osteogenesis markers, while PD98059 suppresses the expression of osteogenic markers. The highest expression of osteogenic markers was observed with co-treatment with 3, 4’-DHF, U0126, and NAC simultaneously (Fig. 5).
In conclusions, In this study, treatment of 3, 4’-DHF led to enhanced osteogenic differentiation of eADSCs and activation of ERK. ERK inhibitor, U0126, stimulated differentiation by BMP-mediated signaling despite PD98059 inhibited differentiation via ERK inactivation. In addition, 3, 4’-DHF regulated ROS-related genes during osteogenic differentiation. Co-treatment with 3, 4’-DHF, U0126, and NAC cumulatively enhanced osteogenesis in eADSCs. The effect of 3, 4’-DHF treatment during osteogenesis in eADSCs may be important for bone formation in the equine industry. In addition, 3, 4’-DHF and combination with U0126 and NAC may represent a new approach for the treatment of osteoporosis, and for potential therapeutic applications in horse regenerative medicine. Our study findings need to be confirmed using in vivo model in further study.
Go to :
 XML Download
XML Download