

 PDF
PDF Citation
Citation Print
Print


Introduction
Human induced pluripotent stem cells (hiPSC), similar to embryonic stem cells (ESC) possessing pluripotency and self-renewal capacity, and importantly facing far few medical ethical challenges, have been used extensively in disease modeling, mechanistical studies, cell replacement therapy, and drug screening, among others (1-4). Through manipulating signaling pathways by small molecular drugs, hiPSC can now be very efficiently differentiated to cardiomyocytes (CMs) (5). However, these hiPSC-CMs are a heterogeneous population consisting of ventricular-, atrial- and sinoatrial-CMs and some no-differentiated hiPSC, which significantly limit their clinical applications and cannot be used to model specific cardiovascular diseases such as atrial fibrillation (AF) (6). In addition, accumulating evidence have shown that the contamination of the sinoatrial node-like cells (SANLCs) is one of the most important factors leading to the development of cardiac arrhythmia post hiPSC-CMs transplantation (7, 8). Therefore, it is of great importance to optimize the differentiation protocols to enrich specific CMs subtypes.
RA (retinoic acid) is the active form of vitamin A that through its receptor participates in both cardiac development and maturation (9). Recently, manipulation of RA signaling pathway has emerged as a critical switch in directing cardiomyocyte differentiation (10). Activation of RA signaling pathway biased the differentiation of hiPSC to atrial-like CMs, whereas inhibition of the signaling pathway enriched the ventricular-like CMs (11, 12). However, the timing and dosage of the RA signaling in the differentiation toward SANLC remain to be studied. Embryonic heart development starts from cardiac crescent derived from primitive streak, which then fuses to form primitive heart tube. As development progresses, the primitive heart tube forms anterior and posterior poles. Precursor cells on the anterior pole develop into ventricles, whereas cells on the posterior pole mainly contribute to the formation of atria and sinus venousus, and the latter happens to be where the sinoatrial node cells locate (13, 14). In adult heart, the sinus node is located at the border region between superior vena cava and right atrium. These discoveries suggest that the spatial distribution of the precursor cells of atrial and sinoatrial node CMs is correlated to their different responses to signaling pathway.
In this study, based on the hiPSC-CMs differentiation protocol adopted and optimized (15) from which RA signaling pathway was manipulated during the transition period from cardiac mesoderm to myocardial precursor cells, we found that activation of RA signaling biased the differentiation to atrial-CMs, whereas inhibition of the signaling pathway enriched SANLCs. These discoveries not only improve our understanding of the roles of RA in CMs differentiation but provide an efficient protocol to enrich SANLCs.
Materials and Methods
Materials
hiPSC (HNF-P30-P11), human embryonic stem cell (hESC) line (HN4) and BioCISO medium was obtained from OSINGLAY BIO (Guangzhou, CHINA). Human fibroblast (hF) cells (BJ) were purchased from ATCC. Anti-Ki67 (ab15580), -TBX18 (ab115262) and -TBX3 (ab154828), -SHOX2 (ab55740) were purchased from Abcam. Anti-NANOG (#3580), -OCT4 (#2750), and -TRA-1-60 (#4746) were all from Cell Signaling Techno-logy. Anti-cTNT (MS-295-P1) was from Thermo-Fisher Scientific. Anti-α-actinin (A7811) was from Sigma-Aldrich. Anti-COUPTFII (PP-H7147-00) was from R&D. Anti-NKX2.5 (MAB2444) was from NOVUS. All other chemi-cals, unless stated otherwise, were products of Sigma -Aldrich.
Maintenance and characterization of hiPSC
hiPSC was cultured in BioCISO medium that was refreshed every day, which was then either propagated or differentiated upon reaching around 80% confluence. hiPSC was characterized by proliferation marker Ki67, hiPSC-specific transcription factors, NANOG and OCT4, and iPS-specific membrane protein, TRA-1-60 using both flow cytometry and immunofluorescence (IF).
Cardiomyocyte differentiation from hiPSC
Cardiomyocyte differentiation was performed by small molecule based temporal modulation of Wnt signaling using GSK inhibitor (Gi) and Wnt inhibitor (Wi) (named as GiWi protocol) (16). Briefly, hiPSC was harvested at 80% confluence using 0.5 mM EDTA and resuspended with hiPSC-maintaining medium to 0.5×105 cells per milliliter (ml). Two ml of the cell suspension was added per well in a 12-well Matrigel-coated plate, and this is designated as day minus 4. At day 0, the cells were refreshed with RPMI/B27 medium without insulin (RPMI/B27−) containing 10 μM GSK3 inhibitor, CHIR99021 (S1263, Sigma, USA), and continued to incubate for 24 h. On day 1, the cells were refreshed with RPMI/B27− and continued to incubate for 48 h. On day 3, the cells were refreshed with RPMI/B27− containing 5 μM Wnt inhibitor, IWP2 (3533, Tocris Bioscience, UK) without insulin and continued to incubate for 48 h. On day 5, the cells were refreshed with RPMI/B27− and continued to incubate for 48 h. From day 7, the cells were refreshed with RPMI/B27 medium containing insulin (RPMI/B27+) every 3 days till day 35. The beating cardiomyocytes can be seen as early as on day 8. The differentiated cardiomyocytes were then characterized by immunofluore-scence for the expression of cTNT (MS-295-P1, Thermo Fisher Scientific, USA) and α-actinin (A7811, Sigma, USA), and by flow cytometry for cTNT expression.
Differentiation of hiPSC toward arial-like and SAN-like cardiomyocytes
At the differentiation day 5∼7, equivalent to the cardiac mesodermal stage in vivo, different concentrations of GiWi signaling pathway agonist, RA (0, 0.5, 1, 2, and 5 μM) and antagonist, BMS (0, 1, 2, 5, and 10 μM) were added to the differentiating cardiomyocytes till day 21, and the expression of atrial markers, NPPA, MYL7, COUPTFII, KCNJ5, and CX40 were analyzed by qPCR to determine the optimum concentrations of RA and BMS that regulate the maximum expression of atrial markers. The optimum concentrations of RA and BMS were then added to the differentiating cardiomyocytes on day 5∼7, and hiPSC was continued to differentiate to day 21 for differentiation efficiency evaluation and to day 60 for electrophysiological analysis by patch-clamp, respectively. qPCR was used for the expression of the Sinus node markers, SHOX2, TBX18, TBX3, HCN4, ISL1, CX30.2, CACNB1, CACNA1A, KCNN4, KCNK2, KCND2, and SCN3B; IF was used for atrial marker (COUPTFII) and the sinus node markers (TBX18 and TBX3); and flow cytometry was used to analyze atrial marker (COUPTFII) and markers for identifying sinus node cell, i.e., cTNT+/NKX2.5− and cTNT+/SHOX2+. Whole cell patch-clamp was used to record the action potential and analyze their electrophysiological characteristics of atrial-, pacemaker-, and ventricular-like cardiomyocytes.
Gene expression analysis
Total RNA was isolated using TRIzol (15596026, Invitrogen, USA). One μg total RNA was reversely transcribed in a total volume of 10 μl with ReverTra Ace qPCR RT Master Mix kit (FSQ-201, TOYOBO, Japan) following the vendor’s instructions. The cDNA was diluted 3 times, and 1 μl was used for real-time PCR in a 20-μl reaction using SYBR Green Real Time PCR Mix (204143, Qiagen, Germany). The PCR conditions were 95℃ for 2 min, followed by 40 cycles of 95℃ for 20" and 60℃ for 15". All primers are listed in Supplementary Table S1. The expression of target gene was normalized to that of GAPDH and calculated using the 2−ΔΔCt method.
Immunofluorescence
Single hiPSC cell suspension and induced cardiomyo-cytes were seeded in a μ-Slide 8 well (80827, ibidi) pre-coated with Matrigel at the density of 2×104 cells per well for 48 h. Cells were fixed with 4% (w/v) paraformaldehyde (PFA) for 15 min at room temperature (RT), permeabilized, blocked in 5% (w/v) BSA in PBS for 30 min, and then incubated with primary antibodies as instructed by vendors at 4℃ for overnight. Then, the plates were washed with PBS on a decolorizing shaker for 3 times at 5 minutes each, followed by incubation with species-specific fluorescence-conjugated secondary antibodies at RT for 1 h. The secondary antibodies were: Alexa fluor 488 labeled goat anti-rabbit IgG (A11008, Invitrogen, USA), alexa fluor 594 labeled goat anti-rabbit IgG (R37177, Invitrogen, USA), and alexa fluor 594 labeled goat anti-mouse IgG (A-11005, Invitrogen, USA). The cells were washed 3 times with PBS and then counterstained using 0.5 μg/ml of DAPI (4083, Cell Signaling Technology, USA) for 15 min at RT. The chambers were mounted and visualized under fluorescence microscopy (IX83, Olympus, Japan). The corresponding antibody isotype controls were: mouse IgG (ab205719, abcam, USA), mouse IgG1 (#5415, Cell Signaling Technology, USA), rabbit IgG (ab205718, abcam, USA), and rabbit IgM (ab37424, abcam, USA).
Flow cytometry analysis
The induced cardiomyocytes were digested with 0.25% trypsin/0.5 mM EDTA into single cell suspension, spun down and washed with PBS. Cells were fixed with 4% formaldehyde for 10 min at RT and chilled on ice for 1 min. Permeabilization was performed by adding one tenth of ice-cold 100% methanol slowly to the pre-chilled cells and cells were continued to incubate on ice for 30 min. Cells were then blocked with blocking buffer (0.5% BSA in PBS) for 10 min; incubated with the primary antibody as indicated in the corresponding figures. for 1 h at RT, washed with PBS, and incubated with the corresponding species-specific fluorescence-conjugated secondary antibodies for 30 min at room temperature. The secondary antibodies used were, Alexa fluor 488 labeled goat anti-mouse IgG (A-11029, Invitrogen, USA), and Alexa fluor 647 labeled goat anti-rabbit IgG (A-32733, Invitrogen, USA). Cells were analyzed using a flow cytometer (651155, BD FACS Verse, BD Bioscience, USA) according to the manufacturer’s protocol. The corresponding antibody isotype controls were: mouse IgG (ab205719, abcam, USA), mouse IgG1 (#5415, Cell Signaling Technology, USA), mouse IgG2a (ab18415, abcam, USA), and rabbit IgG (ab205718, abcam, USA).
Action potential (AP) recording
AP recording was performed following El-Battrawy et al.’s protocol with some modifications (17). Briefly, on day 60 after differentiation, induced cardiomyocytes were dissociated into single cell suspension by incubation with type I collagenase (2 mg/ml) for 30 min, followed by 0.25% trypsin without EDTA for 3 min. 1×104 cells were seeded into a 3.5-cm dish containing a lysine-treated glass coverslip and incubated for 3 days. AP was recorded using the whole cell patch-clamp method. Briefly, adherent cells on the coverslip were placed in the recording chamber and perfused with bath solution containing 140 mM NaCl, 1 mM MgCl2, 5 mM KCl, 1.8 mM CaCl2, 5 mM 4-(2-hydro-xyethyl)-1-piperazineethanesulfonic acid (HEPES), and 10 mM glucose (the pH was adjusted to 7.4, and the osmolality to 301±3 mOsm, respectively). The patch pipettes were pulled from borosilicate glass capillaries (7-000-0650-LHC, Drum-mond, USA) by a horizontal puller (PC100, NARI SHIGE, Japan) and had resistances of 1.5∼3 MΩ. Pipette solution consists of 110 mM K-gluconate, 20 mM KCl, 1 mM CaCl2, 1 mM MgCl2, 10 mM HEPES, 5 mM ethylene glycol tetra-acetic acid potassium chloride (EGTA-KOH), 5 mM ATP-Mg2+, and 5 mM Na-phosphocreatine. The pH was adjusted to 7.2 by KOH, and the osmolality to 290±3 mOsm. A Multiclamp 700B amplifier was used to record APs, and data were analyzed using a custom software.
Results
Characterization of hiPSC
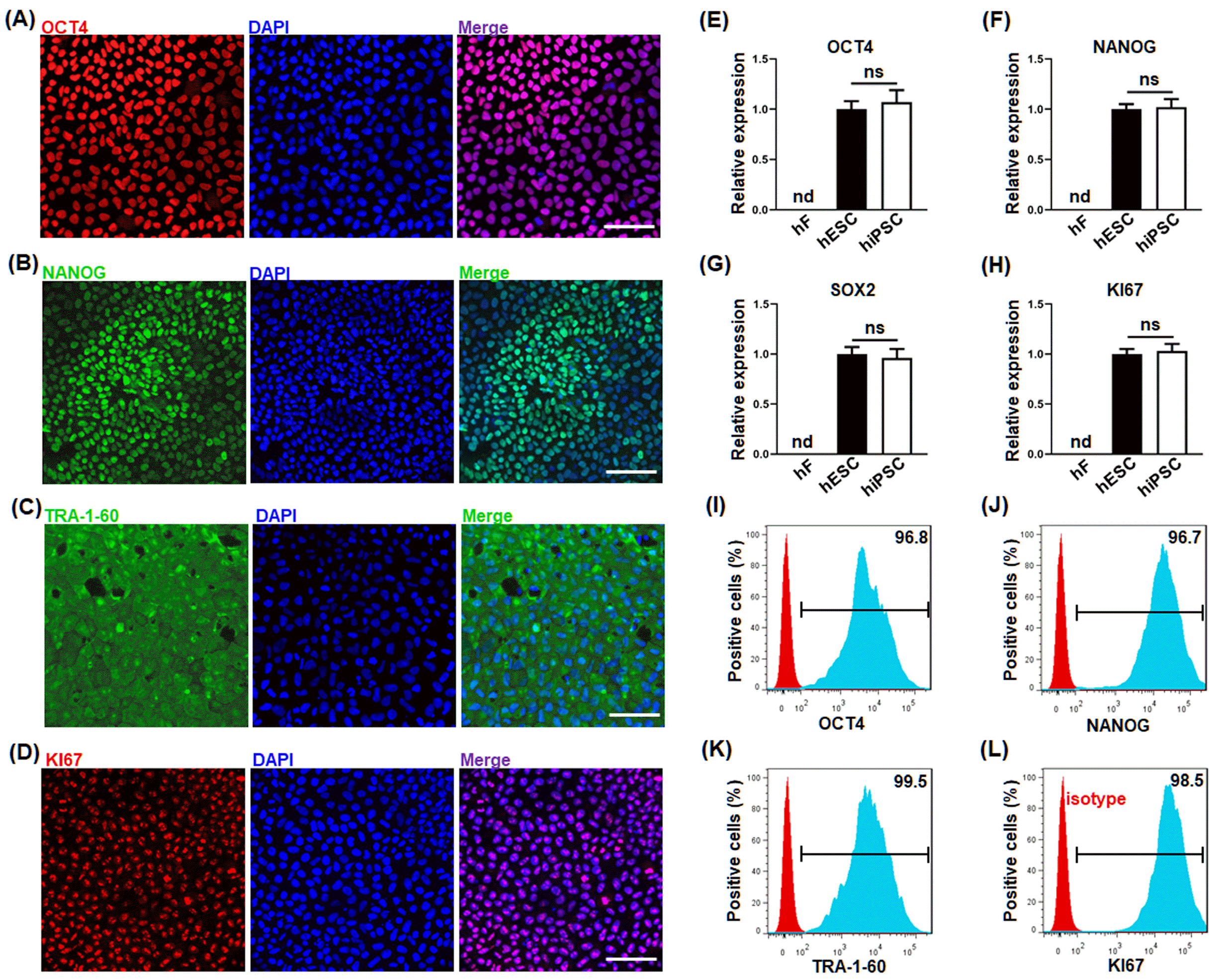
To validate the authenticity of the hiPSC, the expression of the signature molecules of hiPSC (OCT4, NANOG, SOX2 and TRA-1-60) and proliferation marker (Ki67) were evaluated by both IF (A∼D), qPCR (E∼H) and flow cytometry (I∼L). As shown in Fig. 1, the hiPSC highly expressed pluripotent stem cell-specific markers OCT4 (A, E, I), NANOG (B, F, J), SOX2 (G) and TRA-1-60 (C, K), and proliferation marker Ki67 (D, H, L). These results demonstrate that the hiPSC possesses their pluripotency and self-renewal capability.
Characterization of cardiomyocytes derived from hiPSC
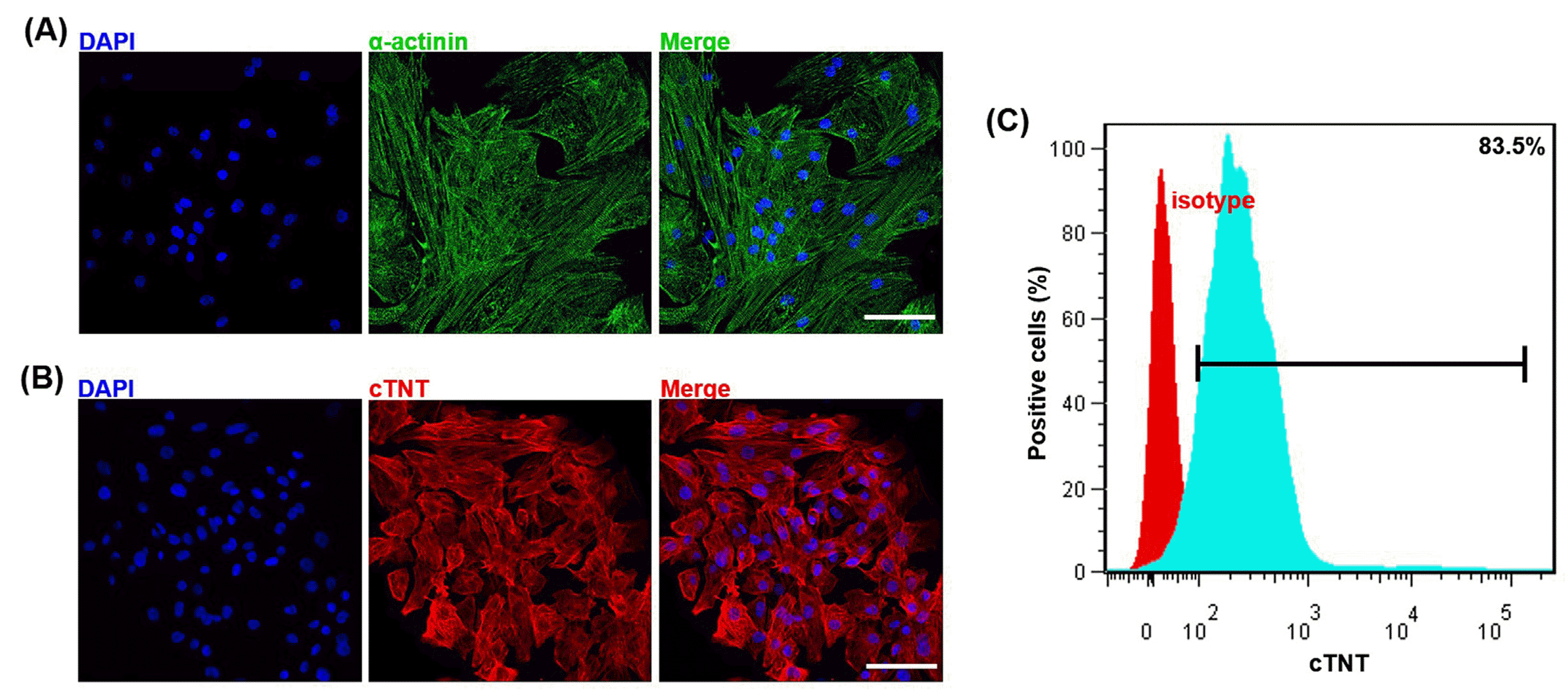
Cardiomyocytes were induced following the GiWi protocol with some modifications (18). On day 35 after the differentiation, the induced cardiomyocytes displayed relatively mature discernable striae and sacromeric structure and were positive for cardiomyocyte-specific markers, α-actinin by IF (Fig. 2A) and cardiac sarcomere proteins, cTNT by both IF (Fig. 2B) and flow cytometry showing that 83.5% of the induced cardiomyocytes were positive for cTNT (Fig. 2C). These results demonstrated the high efficiency of our modified GiWi differentiation protocol.
Activation of RA signaling pathway promotes atrial and simultaneously inhibits Sinoatrial node-like cells differentiation from hiSPC
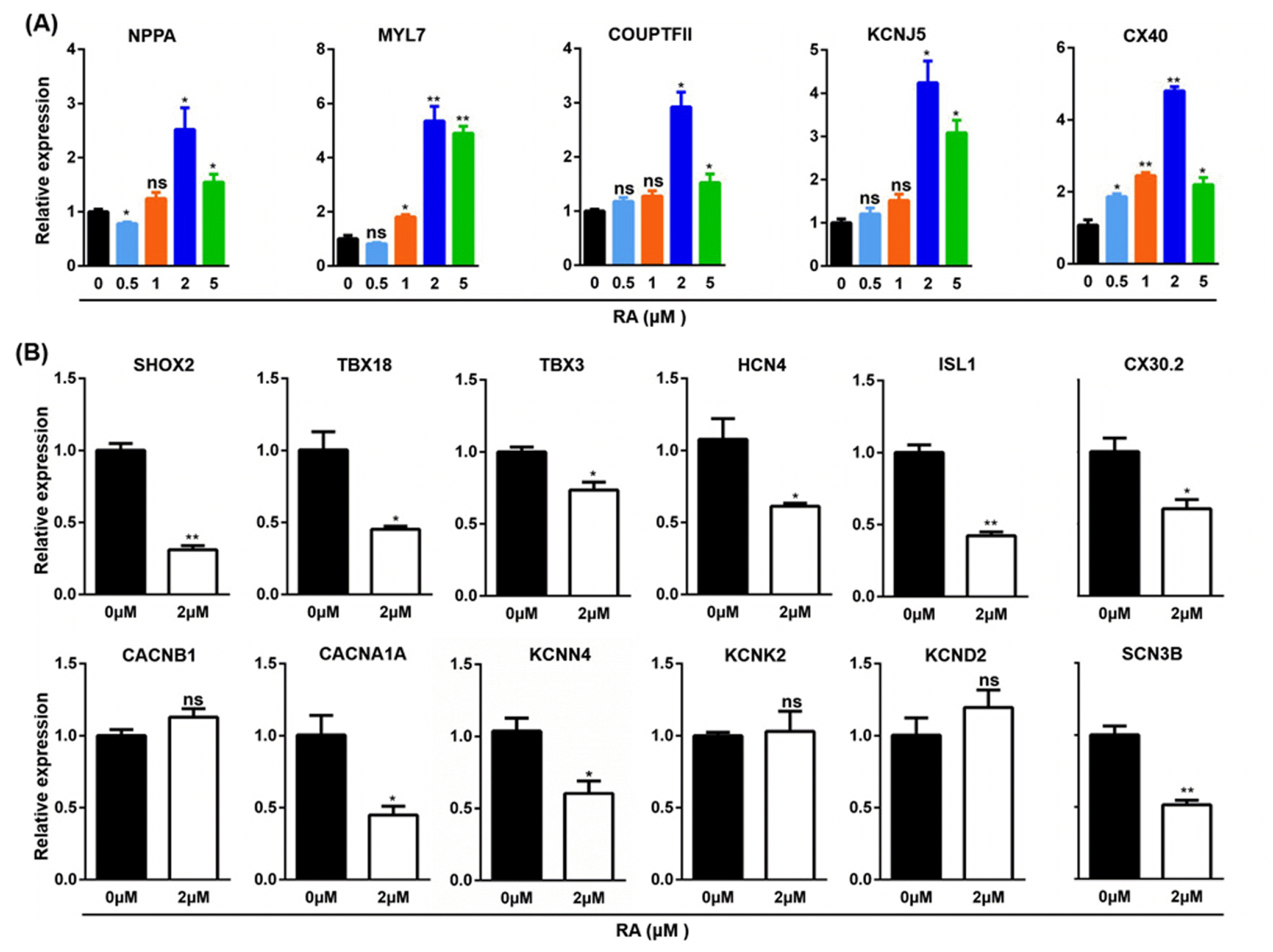
The differentiation of hiPSC to CMs is a spatiotemporally coordinated process. Our previous study showed that day 5 of the GiWi differentiation process was a critical time point for the transition from cardiac mesoderm to cardiac precursor cells (18). RA signaling pathway agonist, RA was introduced into the differentiation on day 5, and atrial cardiomyocyte markers was analyzed on day 21 by qPCR. The results showed that RA at 2 μM significantly induced the expression of natriuretic peptide A (NPPA), myosin light chain 7 (MYL7), COUP transcription factor 2 (COUPTFII), potassium inwardly rectifying channel subfamily J member 5 (KCNJ5), and connexin 40 (CX40) (Fig. 3A). Therefore, 2 μM RA was used to further study the effect of RA on the differentiation of hiPSC to the Sinus node-like cells. When gene expression was analyzed on day 21 (Fig. 3B), RA significantly decreased the expression of sinus node cells-specific transcription factors, SHOX2, TBX18, TBX3 and ISL1, ion channels, HCN4, SCN3B, CACNA1A and KCNN4, and gap junction CX30.2, but no significant effect was observed on the expression of CACNB1, KCNK2 and KCND2.
Inhibition of RA pathway using BMS further increases the efficiency of the differentiation of sinoatrial node-like cells
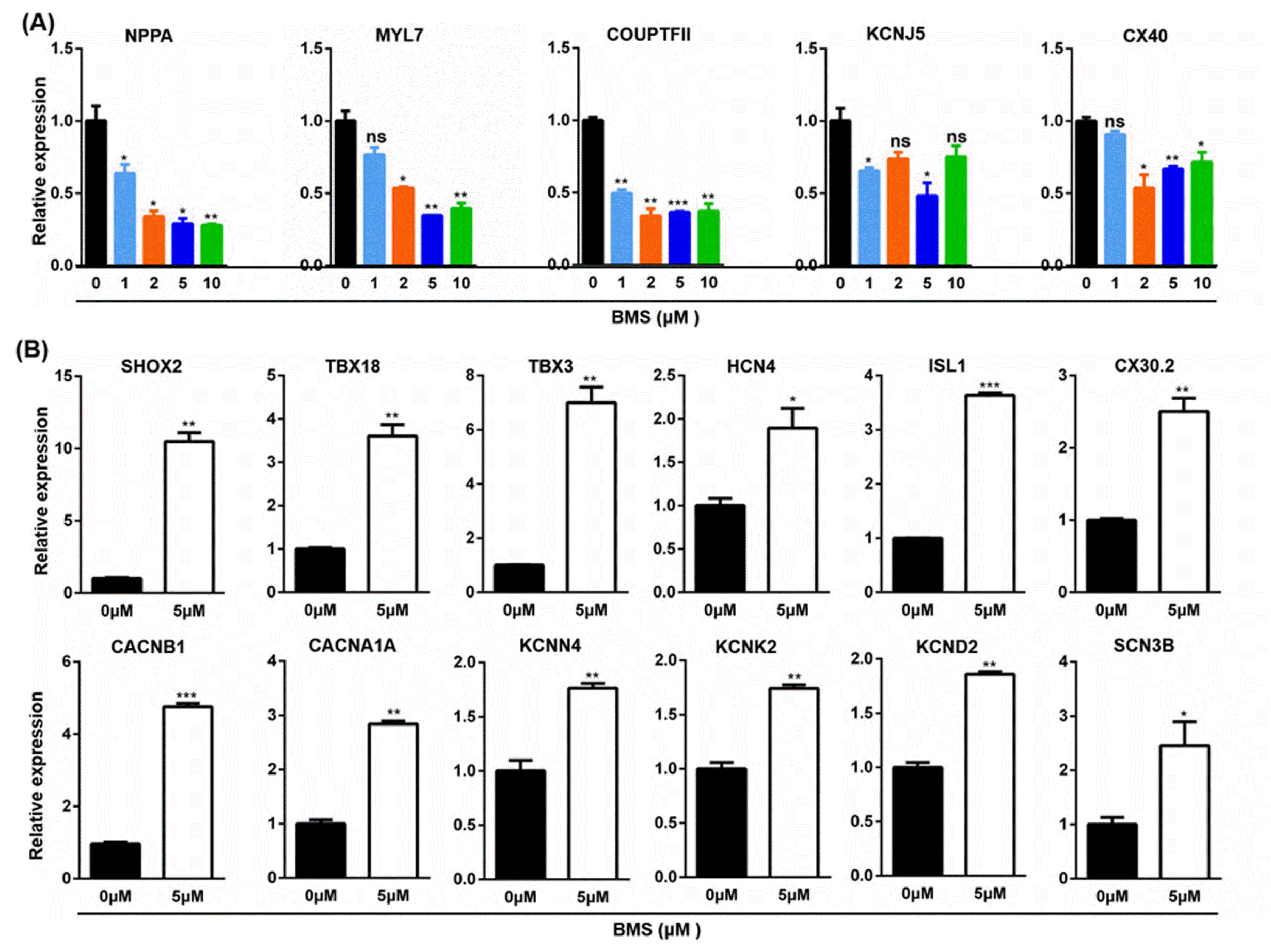
To further investigate the effect of RA signaling pathway on the differentiation of CM subtypes, a RA signaling pathway inhibitor, BMS was introduced on day 5 of the GiWi protocol. When gene expression of atrial cells markers was analyzed on day 21 after the differentiation, BMS dose-dependently inhibited the expression of NPPA, COUPTFII and KCNJ5, and affected the expression of MYL7 and CX40 in a biphasic manner, with 5 and 2 μM inhibited the most on MYL7 and CX40 respectively (Fig. 4A). Therefore, 5 μM was used as the optimum concentration of BMS in the following study. When gene expression was analyzed on day 21, differentiated cardio-myocytes treated with BMS expressed higher levels of sinoatrial node-specific transcription factors, SHOX2, TBX18, TBX3 and ISL1, ion channels, HCN4, CACNB1, CACNA1A, KCNN4, KCNK2, and KCND2, and gap junction CX30.2 (Fig. 4B).
Activation of RA signaling pathway promotes atrial cardiomyocyte differentiation, whereas inhibition RA signaling pathway biases sinoatrial node-like cells differentiation
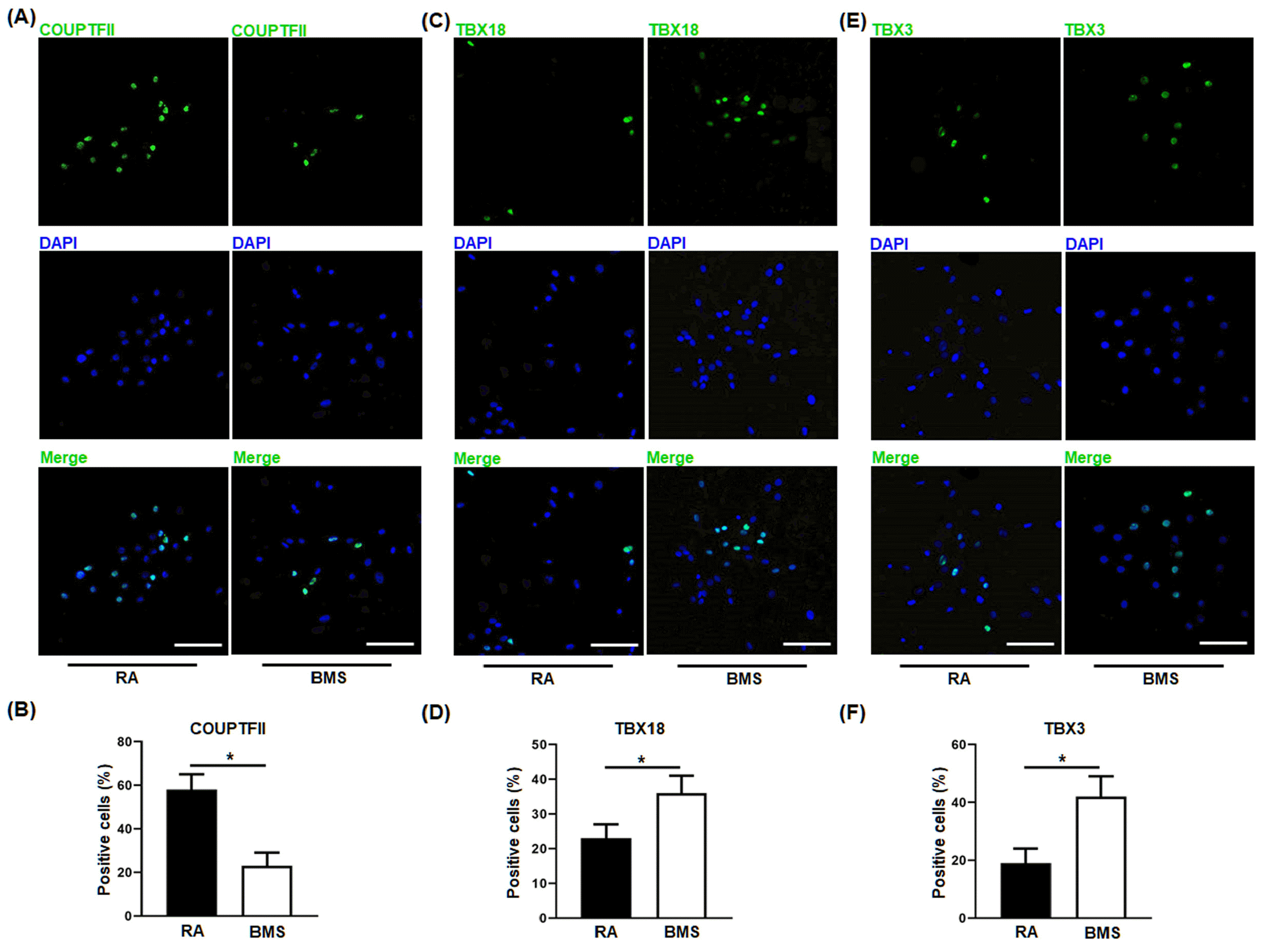
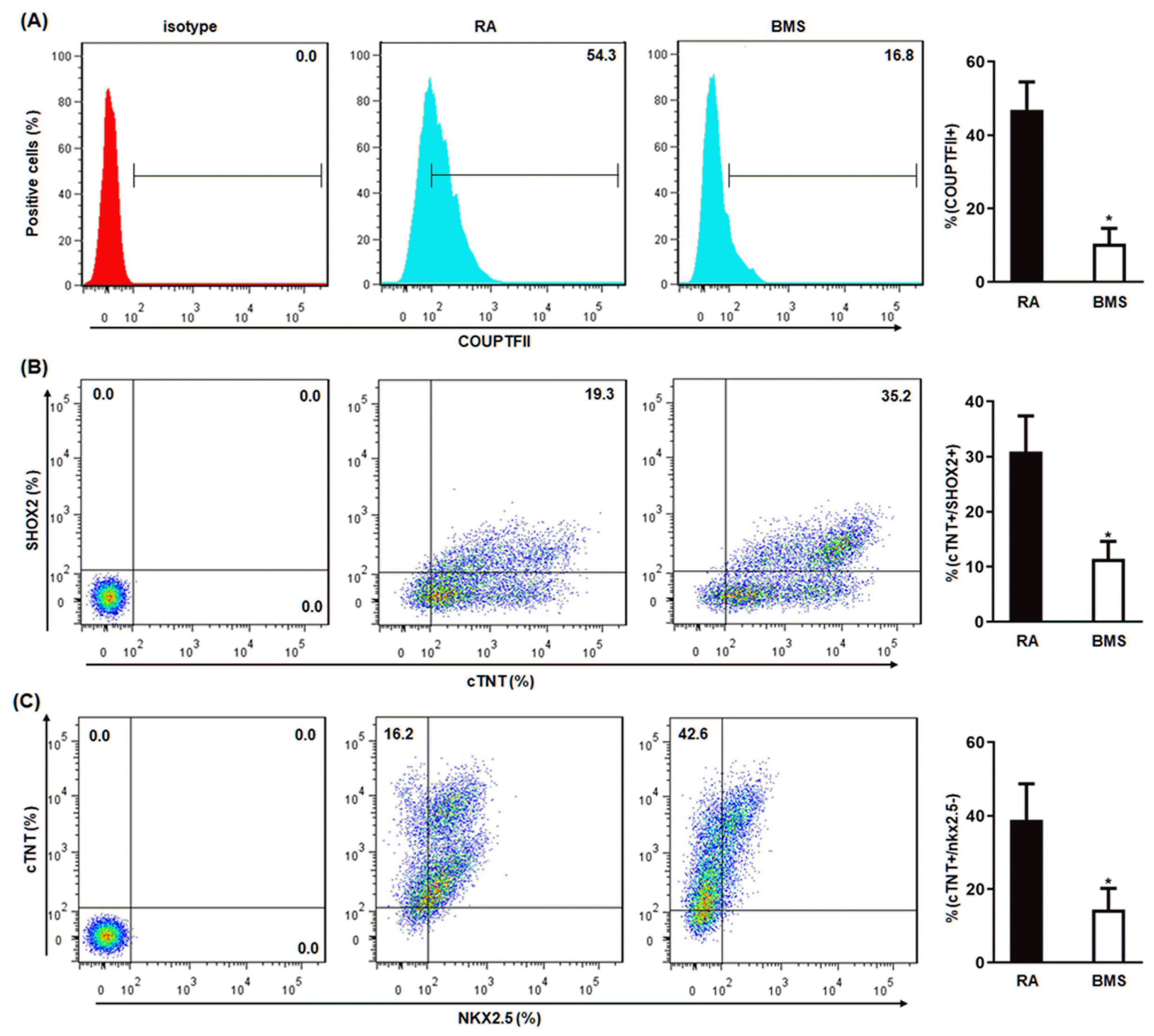
To validate the above findings that RA and BMS promoted the differentiation of atrial and Sinoatrial-like cells respectively, the resultant cardiomyocytes on day 21 were analyzed by IF and flow cytometry, and whole cell patch-clamp was performed on day 60. Compared to RA, BMS decreased the percentage of atrial-specific transcription factor, COUPTFII-positive cells (Fig. 5A, B), whereas increased the percentage of SANLCs-specific transcription factors, TBX18− (Fig. 5C and 5D) and TBX3-po-sitive cells (Fig. 5E and 5F). These results were further confirmed by flow cytometry showing that BMS decreased the percentage of RA-induced COUPTFII positive cells (BMS vs. RA, 10.4±4.2% vs. 46.9±7.6%) (Fig. 6A), and simultaneously increased the percentage of cells expressing SANLC-specific markers, cTNT+/SHOX2+ (BMS vs. RA, 30.9±6.5% vs. 11.4±3.2%) (Fig. 6B), and cTNT+/NKX2.5− MS vs. RA, 38.9±9.8% vs. 14.4±5.8%) (Fig. 6C).
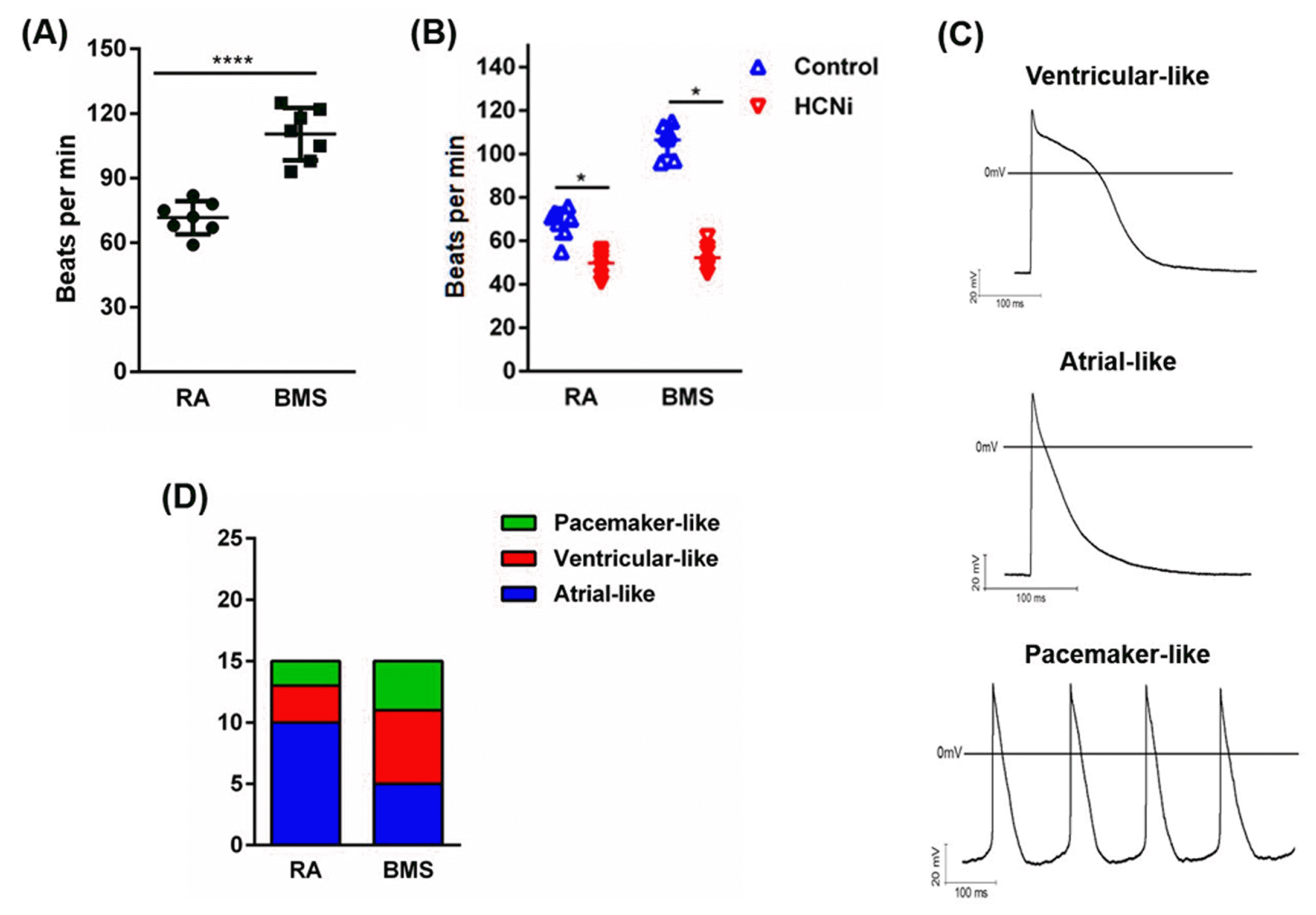
To further characterize the differentiated atrial-like cells and SANLCs, the beating frequency was recorded and action potential (AP) was analyzed by whole cell patch-clamp technique. Compared to RA, BMS significantly increased the beating frequency (times/min) of differentiated cardiomyocytes (RA vs. BMS, 71.57±7.68 vs. 110.43±12.20) (Fig. 7A), and sensitized cardiomyocytes to pacing-related ion channel inhibitor, HCNi (Fig. 7B). AP analysis further showed that the ratio of cardiomyocytes displaying atrial-like AP was significantly higher in RA (10/15) than BMS (5/15) groups, whereas the ratio of cardiomyocytes displaying pacemaker-like AP was significantly higher in BMS (4/15) than RA (2/15) groups (Fig. 7C and 7D).
Discussion
Mounting evidence has shown that RA is one of the most critical signaling molecules orchestrating embryonic development and regulating many crucial steps of the heart morphogenesis, such as differentiation of atria- and ventricular-CMs, and CMs maturation (9, 11). However, how the molecular mechanisms governing the differentiation of sinoatrial-like CMs remains largely unknown, which significantly limits the enrichment of the sinoatrial-like CMs for basic research and regenerative medicine. In this report, the GiWi protocol was optimized to efficiently differentiate hiPSC to pan-CMs, from which the RA signaling pathway was inhibited on day 5 post-differentiation, equivalent to the transition from cardiac mesoderm to the CM precursor cells in vivo (18), and the resultant CMs were analyzed at different time points. We found that after RA signaling pathway inhibition, the expression of atrial-CMs markers at both mRNA and protein levels were significantly suppressed, while that of sinoatrial node-like cells markers were significantly activated on day 21 post-differentiation. When CMs were analyzed on day 60 post-differentiation, the percentage of CMs showing action potential (AP) characteristic of sinoatrial-like CMs were significantly increased, whereas that of atrial CMs were significantly decreased.
Cardiac subtype specification is a precisely and spatiotemporally regulated process and atrial cardiomyocytes could be derived from cardiac mesoderm population via RA activation (11, 15). Consistent with previous reports, we observed that activation of RA during the period from cardiac mesoderm to cardiac progenitor significantly enriched the ratio of atrial-like cells from hiPSC using our previously optimized GiWi protocol (18). In addition to the enrichment effect of atrial CM subtype from human pluripotent stem cells, RA has also been reported to exert inhibitory effect on the specification of the ventricular subtype (11). However, whose effect on the generation sinoatrial node-like cells remains unclear. It was observed in our current study that activating the RA signaling pathway with RA inhibited the expression of sinoatrial CM-specific markers and then the yield of sinoatrial cells. this was further supported by inactivating RA pathway using BMS. As far as our knowledge, it is the first finding that in addition to the pro-atrial and anti-ventricular effect, RA activation also has inhibitory effect on the sinoatrial node direction although the related mechanism remains to be elucidated.
It was evidenced by Lee, et al’s study (11) that human ventricular and atrial cardiomyocytes develop from distinct mesoderm populations that are specified with different concentrations of BMP4 and Activin A and can be identified based on the expression of CD235a (Glycophorin A) and RALDH2, respectively. the RALDH2+, but not the CD235a+, mesoderm population responds to retinol to generate atrial cardiomyocytes. In consistence, our study showed that the cardiac mesoderm cells responding to the RA activator could differentiated into atrial subtype CMs in high-efficiency and vice versa. Moreover, it was also found that the activation and inhibition of RA pathway could also antagonize and promote the generation of sinoatrial node-like cells, respectively. It is therefore speculated that in addition to the ventricular and atrial ancestor sub-population, there also exists the sub-population of sinoatrial node ancestor during the cardiac mesoderm stage whose conversion into sinoatrial node cells is at least depend on the RA modulation. It will be an interesting study in our future work to verify this possible sinoatrial node mesoderm subpopulation and elucidate the involved signaling pathways modulating its differentiation of hiPSC into sinoatrial node CMs.
Currently, there are mainly two strategies to generate sinoatrial CMs. One is directing hiPSC into sinoatrial CMs through genetic engineering of all transcription factors necessary for the development of sinoatrial CMs. This strategy is technically challenging, costly, and potentially tumorigenic, which has limited its application (19-21). The other strategy is manipulating the signaling pathways necessary for the specification of CM subtype through chemical reagents. This strategy circumvents the drawbacks of the former and has attracted much attention lately. For example, at the cardiac mesoderm stage, Protze et al. (22) showed that low concentration of bone morphogenetic protein (BMP) could promote the differentiation toward sinoatrial CMs, Yechikov et al. (23) demonstrated that inhibition of nodal signaling pathway by SB431542 down-regulated the expression of the paired-like homeodomain transcription factor 2C (PITX2c), which promoted the differentiation of pacemaker-like CMs, and we showed previously that coordinated manipulation of BMP, RA and fibroblast growth factor (FGF) signaling pathways at the cardiac mesoderm stage enriched the differentiation of sinoatrial CMs (18). Although manipulation of different signaling pathways achieved high differentiation efficiency of sinoatrial CMs, the permutation of small chemicals required and the complexity of performing the differentiation in individual protocol remain to be optimized.
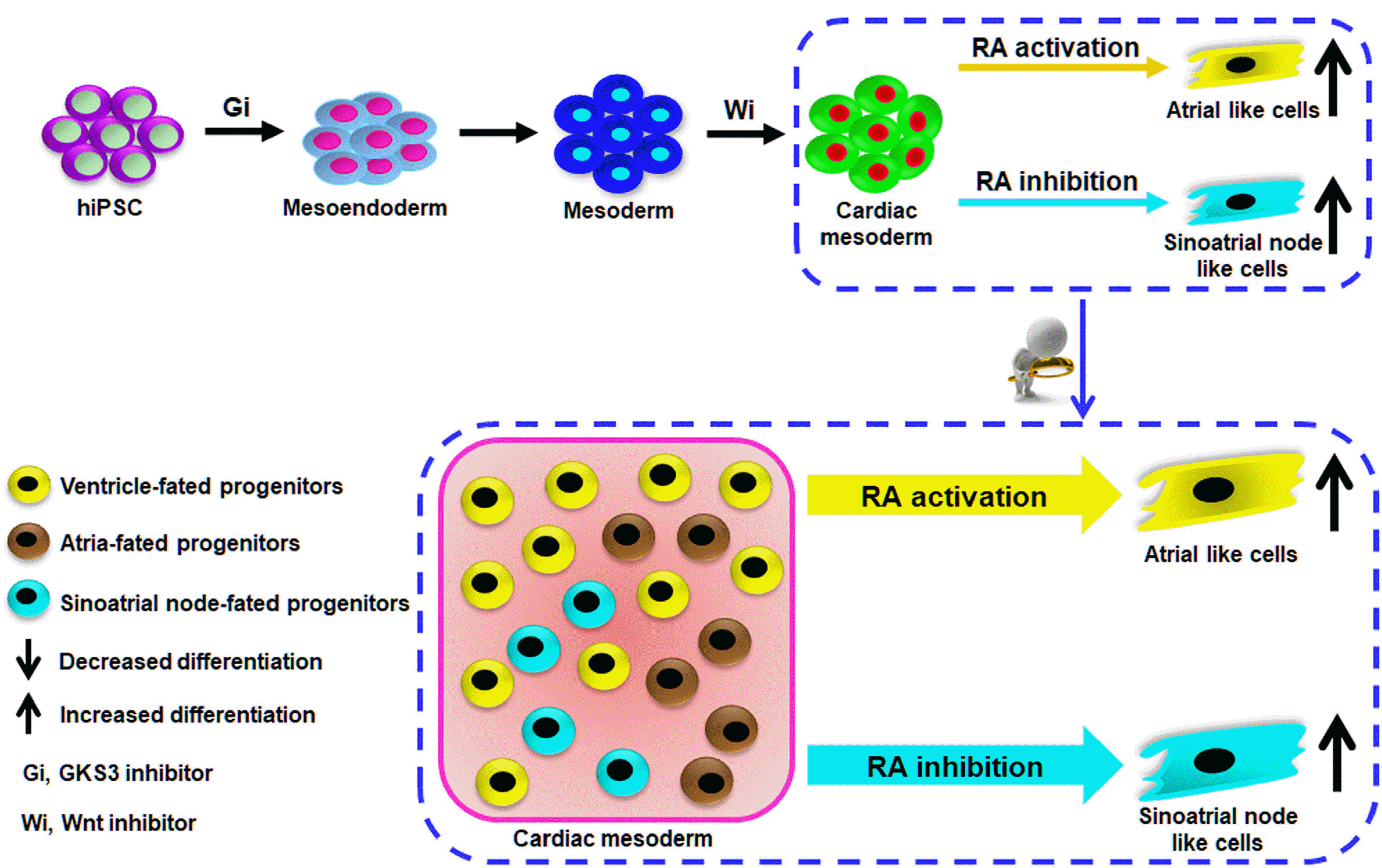
In summary, we developed a de novo differentiation protocol for enrichment of CM subtypes from hiPSC by manipulating the RA signaling pathway during cardiac mesoderm stage of hiPSC differentiation in the biphasic manner, in which activation of RA signaling enabled biased differentiation of atria-like cells as reported before. Moreover, RA pathway inhibition directed the enrichment of sinoatrial node-like cells generation (Fig. 8). It further broadened the understanding of the effect of RA signaling pathway on the embryonic heart development and CM subtypes specification from human pluripotent stem cells.
Supplementary Materials
Supplementary data including one table can be found with this article online at https://doi.org/10.15283/ijsc21148.
 XML Download
XML Download