

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Tinnitus can be defined as the conscious awareness of a tonal and/or noise sound in the milieu of no identifiable corresponding external acoustic source [1]. By contrast, tinnitus disorder is defined as tinnitus with tinnitus-associated suffering, which can consist of emotional distress, cognitive dysfunction and/or autonomic arousal (i.e., stress), leading to functional disability [1]. In other words, tinnitus can be equated to the perception of a phantom sound, and tinnitus disorder is tinnitus accompanied by associated suffering, with or without functional disability. It has been shown that chronic tinnitus (tinnitus with a duration of 3 months or more [1]) is not simply a temporal extension of tinnitus of recent onset but involves distinct mechanisms [2,3]. Tinnitus has a high prevalence, between 12% and 30% [4], increases with age, is more common in men than women, and is expressed more in the left than right ear [4].
The phantom sound is often associated with comorbidities, especially in the auditory domain, such as hearing loss (80% in the elderly) [5] and hyperacusis (7%–70%) [6] being the most common ones. However, tinnitus is also frequently associated with emotional, cognitive, and autonomic problems, thereby qualifying as tinnitus disorder. Indeed, stress is common in tinnitus patients (27%) [7], as are anxiety (26%) and depression (26%). Insomnia is present in 60%–73% of tinnitus patients [8,9]. Tinnitus is also associated with poorer performance across a variety of broad cognitive domains including executive function, cognitive processing speed, short-term memory, and general learning and memory retrieval [10]. This is mainly due to tinnitus-related distress [11]. The natural course of tinnitus is unfavorable. In 18% of patients tinnitus resolves spontaneously in 4 years, while in the other 82% of patients in whom tinnitus persists, it improves in 9% and worsens in 9% [12].
The mainstay of tinnitus treatment is cognitive behavioral therapy (CBT), for which there is meta-analytic evidence of its benefits [13]. Yet, CBT results only in a 10% improvement of the distress, without decreasing the loudness [13]. Furthermore, the meta-analysis could not find a benefit 6 months after CBT treatment [13], and it is highly uncertain whether CBT reduces anxiety, improves health-related quality of life, or reduces negatively biased interpretations of tinnitus [13]. To develop more efficacious tinnitus treatments, a better understanding of the pathophysiological mechanisms that generate and maintain chronic tinnitus is required. Better treatments should treat both the sound and the suffering. Pain and tinnitus share an analogous pathophysiology [14-25], clinical phenomenology [14,19-21,23], and neuromodulatory treatment approaches [19-21,23,26,27].
Chronic tinnitus has been anatomically and symptomatically dissociated into three separable but interacting ascending or descending pathways [16], analogous to what has been described for pain [16,28-33]. These consist of an ascending lateral “sound” pathway, an ascending non-specific medial “suffering” pathway, and a descending noise-canceling pathway [16,34,35]. The medial suffering pathway may overlap with the salience network, analogous to what has been shown for pain [29]. Whereas the lateral auditory and medial salience pathways can explain the loudness and emotional aspects of tinnitus respectively, no correlates have been proposed for the chronification and functional disability encountered in chronic tinnitus. We here propose to extend the current tinnitus networks to the triple cortical network model to fill this gap.
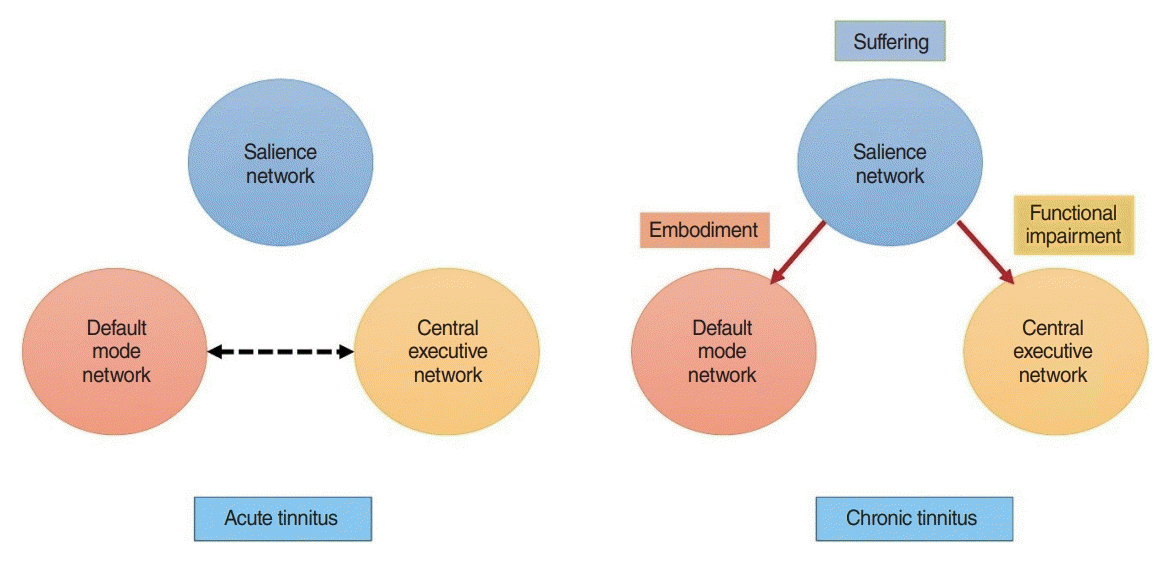
The triple network model is a network science-based approach to find a common framework for understanding cognitive and affective disorders [36]. It proposes that abnormal interactions within and between the three cardinal brain networks underlie neuropsychiatric disorders [36]. These three networks include the self-representational default mode network (i.e., a brain network that is active when a person is not focused on the outside world) [37,38], the behavioral relevance–encoding salience network (i.e., a brain network that selects which stimuli are salient and thus deserving of our attention) [39], and the goal-oriented frontoparietal central executive network (i.e., a brain network crucial for maintaining and processing information in working memory, problem-solving and decision-making) [39,40]. Normally, the central executive network and salience network demonstrate correlated activity, and both networks are anti-correlated to the default mode network [41]. The salience network drives the switch between the anti-correlated default mode network and the central executive network [42-44]. This is in keeping with the proposed functions of the three networks. When the salience network detects a behaviorally relevant external stimulus, it decreases the activity of the self-oriented and mind-wandering default mode network and activates the central executive network to address the salient external stimulus in a goal-oriented fashion. In many brain disorders, such as attention-deficit/hyperactivity disorder, anxiety, depression, bipolar, autism, obsessive-compulsive disorder, posttraumatic stress disorder, and schizophrenia, the functional connectivity (statistical relationships between a pair of brain regions that covary or correlate over time) within and between these three cardinal networks is aberrant [36,45,46].
We propose that in chronic tinnitus, analogous to what has been suggested for pain [47], the three known tinnitus pathways can be linked and extended to the triple network model, which would explain the chronification of tinnitus as well as propose neural correlates for the commonly associated cognitive dysfunction.
THE KNOWN BRAIN ANATOMY OF TINNITUS
A stimulus produces an effect on the different sensory receptors, which is transmitted to the sensory cortex, inducing sensation [14]. Further processing of this sensory stimulation by other brain networks such as the default mode, salience network, and central executive network (i.e., the triple network) permits the sound stimulus to reach consciousness [48-53], and generates an internal representation of the outer and inner world, namely a percept [14]. Tinnitus perception can thus be defined as the act of interpreting and organizing a sound stimulus to produce a meaningful experience of the world and of oneself [14].
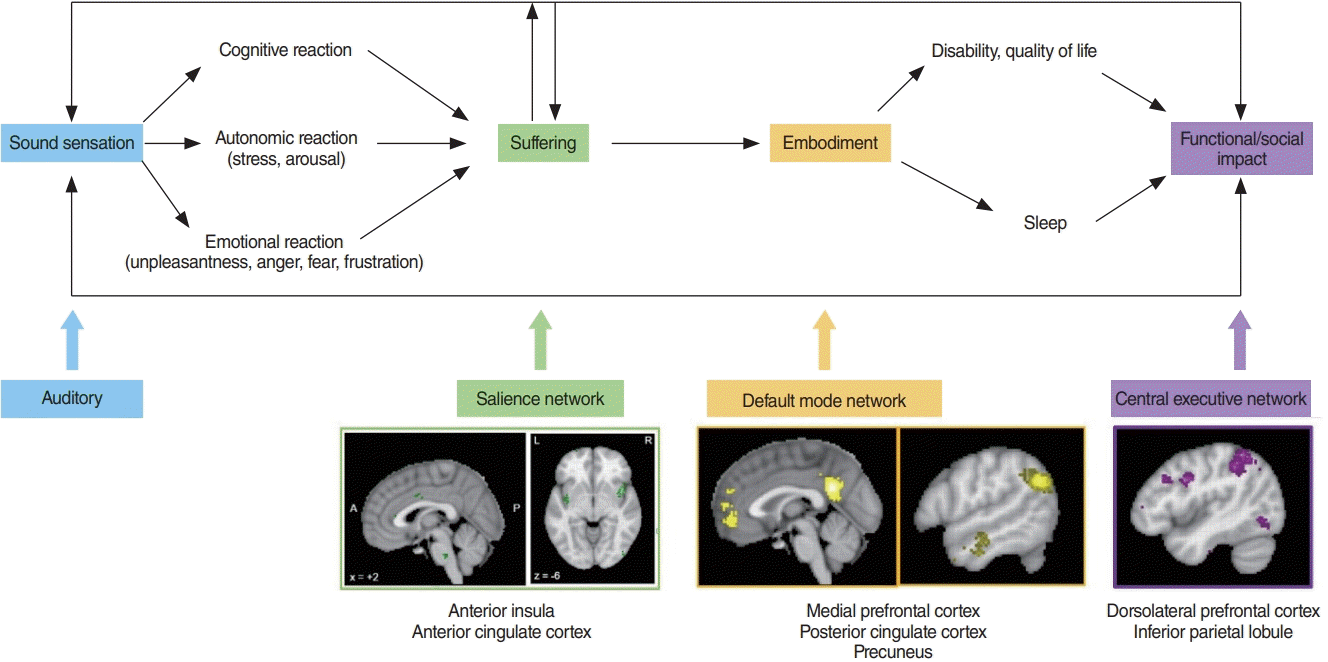
When a patient visits a health care provider and states that he or she has tinnitus, what the person is really conveying is, “I have a certain amount of tinnitus loudness associated with a certain amount of suffering during a certain amount of time.” These three aspects of the unified tinnitus percept can be traced to the three different pathways involved in tinnitus processing (Fig. 1) [16].
As mentioned, the two main tinnitus-activating pathways include the anatomically and functionally separable medial and lateral tinnitus pathways [16]. The medial ascending pathway involves the rostral to dorsal anterior cingulate cortex (rdACC) and anterior insular cortex and processes the affective motivational aspect of tinnitus [14,16,54-56]. The causality has been demonstrated by the fact that cingulotomies could abate negative affect and cognitive control [57,58], as researchers stated: “All the patients except two found the head noises less distressing after the operation but objectively the noises were unchanged” [57].
The lateral ascending pathway involving the auditory cortex encodes the discriminatory components of the tinnitus, such as loudness [59] and tinnitus localization [60]. The two activating tinnitus pathways are balanced by the noise-canceling inhibitory pathway, involving the pregenual anterior cingulate cortex (pgACC) [16,53]. The descending noise-canceling pathway determines the amount of time that tinnitus is consciously perceived [61,62], and therefore reflects the capacity of the brain to suppress acute or ongoing tinnitus [61].
TINNITUS AND SUFFERING ARE DIFFERENT
As above-mentioned, tinnitus can be defined as the conscious awareness of a tonal and/or noise sound for which there is no identifiable corresponding external acoustic source, while tinnitus disorder is defined as tinnitus with suffering [1]. Suffering can be defined as an unpleasant experience associated with negative cognitive, emotional, and autonomic responses to a (tinnitus) stimulus [29]. About 80% of the people with tinnitus are not bothered by the phantom sound, but in 20% it is severely bothersome [63], qualifying as tinnitus disorder [1].
The sensation of a phantom sound can lead to suffering via the associated feeling of (emotional) unpleasantness and (cognitive) catastrophizing. Tinnitus catastrophizing is characterized by (1) a tendency to magnify the threat value of tinnitus, (2) feeling helpless in the context of tinnitus, and (3) a relative inability to inhibit tinnitus-related thoughts (rumination) [64]. Thus, tinnitus catastrophizing acts as an amplifier of unpleasantness and tinnitus loudness by deficient cognitive coping strategies [65]. The combination of the perceived unpleasantness and catastrophizing leads to suffering, which can express in different behaviors such as anger, fear, and frustration [64,65] (Fig. 1).
Tinnitus is often associated with stress [7,66-68]. Physiological stress can be defined as an unpleasant sensory, emotional, and subjective experience that is associated with potential damage of body tissue and bodily threat [69], especially when an environmental demand exceeds the natural regulatory capacity of an organism [70]. Stress results in an immediate adaptive temporary response of the autonomic nervous system and slower protracted stimulation of the hypothalamic-pituitary adrenal endocrine axis [71]. Whereas acute stress responses are adaptive and beneficial to survival by preparing a fight-or-flight response, chronic stress becomes maladaptive, leading to a host of problems including metabolic syndrome, obesity, cancer, mental health disorders, cardiovascular disease, and increased susceptibility to infections [72]. Chronic stress can result in depression [72,73], anxiety [73], and anger [74]—in other words, chronic suffering.
Tinnitus distress correlates with activity in the rdACC [55,56,75]. The increased cortisol level in stress results in unpleasantness via its functional modulation of the rdACC [76]. The neural substrates of physiological stress, as identified by a meta-analysis also involve the rdACC and anterior insula [69]. Unpleasantness (suffering) and sound intensity (loudness) can be modulated independently. This has been known since the 1950s, as following frontal lobotomies performed for tinnitus it was realized that “In fact, of the 19 patients who survived the operation, 11 felt that their head noises were just the same but bothered them less and 8 felt that they had improved” [57]. Similar observations were made by other neurosurgeons [58] and are equivalent to changes noted with electrode implants in the rdACC [54].
In summary, tinnitus consists of a sensory loudness component, encoded by the ascending lateral pathway, and a suffering component, encoded by the ascending medial pathway. Suffering involves a cognitive, emotional, and autonomic component, all encoded by parts of the medial pathway. The medial and lateral pathways are separable, and consequently, one may have tinnitus without suffering and suffering without tinnitus.
TINNITUS CHRONIFICATION, ENERGY EXPENDITURE AND THE DEFAULT MODE NETWORK
Network science is a research field studying complex networks such as computer, economic, biological, social, cognitive, and semantic networks, abstracting the networks to nodes (or vertices) and their connections (or edges). Network science is increasingly used to understand the involvement of resting-state network interactions in brain disorders [77-80], including tinnitus [81,82]. One of its findings is the involvement of the default mode network in tinnitus [83-91]. It can be hypothesized that in chronic tinnitus the default mode network, which controls self-representational processing may become pathologically connected to tinnitus-provoking networks [89]. The significance of this finding is tremendous, as this may be a neurobiological rationale why in chronic tinnitus the sound becomes embodied—that is, an integral part of the self, the new normal default state [92]—thereby making treatments more difficult [93]. Furthermore, not only can tinnitus become an integral part of the self, but when suffering becomes chronic, fear can turn into anxiety and sadness into depression, all common comorbidities in tinnitus.
A question can be raised, “why does this connectivity to the default mode occur?” An evolutionary explanation can be proposed that involves the free energy principle of brain functioning [94,95]. In sum, it posits that an energy-expensive organ, such as the brain, tries to conserve energy in whatever way it can [96,97]. It is of interest that the salience network and the sympathetic central control overlap in the brain [98], and the central component of the brain’s parasympathetic nervous network partially overlaps with the default mode network [99].
The sympathetic nervous system increases intrinsic energy consumption by 15%–35% [100,101]. In recent-onset pain, the daily energy consumption is increased by 60% [102], whereas in more chronic pain, the daily extra energy expenditure is only increased by 15% [103,104]. Similarly, fear increases energy expenditure by 22% [105], whereas chronic anxiety only increases energy expenditure by 6% [106]. By rewiring to connect the tinnitus pathways to the default mode network, which broadly overlaps with the parasympathetic central network, energy expenditure can therefore be saved. Data on energy expenditure in acute tinnitus versus chronic tinnitus have not been published yet, but might hopefully be performed, as to verify whether this concept holds for tinnitus as well.
TINNITUS, COGNITIVE DYSFUNCTION, AND THE CENTRAL EXECUTIVE NETWORK
Chronic tinnitus can lead to a lower quality of life and the development of tinnitus-related disability, especially cognitive disability [10,11,107,108]. Tinnitus loudness, tinnitus distress, and tinnitus duration correlate positively with different cognitive measures (e.g., the trail-making test, Montreal cognitive assessment, Mini-Mental State Examination) [109]. Based on network science principles, each aspect of tinnitus could be the result of connectivity changes between the lateral pathway (i.e., the auditory network), and another resting-state network, such as the salience network (suffering), the default mode network (embodiment), the central executive network (cognitive disability), and motor network (physical disability). Changes in the auditory [110,111], salience [112-116], default mode [83-91], and central executive networks [87,113] have been shown in patients with tinnitus. In a healthy state, the salience network, which overlaps with the ascending medial pathway, and the stress network are anti-correlated with the default mode network [41]. In chronic tinnitus, this anti-correlation is lost, as suggested by increased functional connectivity between the rdACC and precuneus [91]. Furthermore, interference in the goal-oriented central executive network may lead to functional impairment in subjects with tinnitus (Fig. 2).
CONCLUSION
Tinnitus is processed by three distinguishable but interacting networks, each encoding different tinnitus characteristics. The ascending lateral pathway, with the auditory cortex as the main hub, is responsible predominantly for loudness. The ascending medial pathway, with the rdACC and insula as the main hubs, are involved in the suffering component, and the descending noise-canceling pathway, with the pgACC as the main hub, is related to the percentage of the time that the tinnitus is present. When the tinnitus sensation pathways become correlated to the default mode instead of anti-correlated, tinnitus becomes part of the self-percept (i.e., part of who one is, the new normal). This can subsequently lead to functional disability by interfering with the goal-oriented central executive network. Therefore, the three tinnitus pathways previously described need to be extended to incorporate the triple network to explain the full clinical picture of chronic tinnitus.
HIGHLIGHTS
▪ The triple network model is a novel unifying framework common to neuropsychiatric disorders.
▪ The salience network encodes the behavioral relevance of tinnitus.
▪ The default mode network makes tinnitus an intrinsic part of the self-percept.
▪ Tinnitus can lead to functional disability by interfering with the central executive network.
 XML Download
XML Download