

 PDF
PDF Citation
Citation Print
Print



INTRODUCTION
Transforming growth factor-beta (TGF-β) is a multifunctional cytokine that regulates many essential functions in the cell, such as proliferation, migration, adhesion, differentiation, and modification of the cellular microenvironment.1 It also plays a dichotomous role in tumor suppression and tumor progression. TGF-β signatures have been explored in mouse hepatocytes, and they have been reported to predict the clinical outcomes of patients with hepatocellular carcinoma (HCC); patients with HCC exhibiting early TGF-β signatures showed a better prognosis than those with HCC exhibiting late TGF-β signatures.2 The genes involved in the transcriptional activation of cell cycle arrest and apoptosis (GADD45β, etc.), those having liver-specific functions (e.g., glucose and lipid metabolism or detoxification; FBP1, etc.), and those associated with antioxidant responses (catalase and superoxide dismutase 1; CYP1A2 and CYP3A4, etc.) have been reported as the early TGF-β-responsive genes.2 Therefore, early TGF-β signatures may have protective roles against tumor formation and progression.
In contrast, the late responsive TGF-β genes were found to be involved with epithelial-mesenchymal transition (EMT) (Twist and Snail, etc.), metastasis, and angiogenesis-processes related to the aggressive behavior of tumors.2 There is growing evidence that the acquisition of stem cell features in epithelial cells is induced by EMT activation.3,4 Ectopic expression of either Twist or Snail transcription factors, two key molecules of EMT, was shown to generate cancer stem cell (CSC) properties in human breast cancer cells.3 We also reported that HCCs expressing stemness markers showed high expression of EMT and hypoxia markers and were related to poor prognosis5,6, and that cancer stemness features were upregulated during the progression of B virus-associated multistep hepatocarcinogenesis and their expression peaked in HCC.7
Accumulating evidence has demonstrated that the development and progression of human hepatocarcinogenesis is a multistep process transitioning from cirrhosis, to low grade dysplastic nodules (LGDNs), high grade dysplastic nodules (HGDNs), early HCCs (eHCCs), and finally progressed HCCs (pHCCs).7,8 Although TGF-β is known to play a central role in hepatocarcinogenesis, the early response and late responsive genes of TGF-β signatures have not been evaluated in defined lesions of human multistep hepatocarcinogenesis. To evaluate the dual role of TGF-β signaling in human hepatitis B virus (HBV)-related multistep hepatocarcinogenesis, we analyzed the expression levels of TGF-β signaling genes (TGFB1, TGFBR1, TGFBR2 and SMAD4), early response genes (FBP1, GADD45B, CYP1A2, and CYP3A4), and late response genes (TWIST and SNAI1) in cirrhosis, LGDNs, HGDNs, eHCCs, and pHCCs.
METHODS
1. Liver tissue samples and pathological examination
A total of 94 patients who underwent curative resection or transplantation at Severance Hospital, Yonsei University Medical Center, were included in this study; their liver samples consisted of 36 LGDNs, 45 HGDNs, 30 eHCCs, and 51 pHCCs. The patients comprised 72 males and 22 females, aged from 40 to 71 years (54.60±7.22, mean±standard deviation). All patients had HBV-related disease. Representative tissue section from surgical specimens were selected for histological examination and immunohistochemical analyses. All nodular lesions were evaluated according to the criteria proposed by the International Consensus Group for Hepatocellular Neoplasia.9
Liver tissue samples were snap-frozen in liquid nitrogen and stored at -80℃. Fresh frozen liver specimens were provided by the Liver Cancer Specimen Bank and the National Research Resource Bank program of the Korea Science and Engineering Foundation under the Ministry of Science and Technology. This study was approved by the Institutional Review Board of Severance Hospital, Yonsei University College of Medicine and the need for patient consent was waived (2014-0253-004, Seoul, Korea). The Strengthening the Reporting of Observational studies in Epidemiology (STROBE) reporting guidelines were followed (Supplementary Table 1).
2. Total RNA extraction and quantitative real-time polymerase chain reaction (PCR)
The total RNA was extracted, and cDNA was synthesized from 108 snap-frozen liver tissue samples using TRIzol and High Capacity RNA-to-cDNA kit (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. TaqMan Gene Expression Assay kits used in this study were purchased from Applied Biosystems (Foster City, CA, USA); the sequences of their primer/probe sets are summarized in Supplementary Table 2. All experiments were performed in triplicate for each cDNA and the relative expression levels of the target mRNAs were normalized to GAPDH mRNA levels. PCR reactions were performed using gene-specific primers and probes with an ABI PRISM 7700 Sequence Detection System and software (Applied Biosystems) according to the TaqMan protocol. A non-template reaction was included in all experiments as a negative control.
3. Immunohistochemistry
Immunohistochemical staining was performed to detect the expression of CYP1A2, CYP3A4, and FBP1. The antibodies used and the antigen-retrieval conditions are listed in Supplementary Table 3. Representative sections of formalin-fixed, paraffin-embedded tissues from 31 LGDNs, 37 HGDNs, 21 eHCCs, and 42 pHCCs were subjected to immunohistochemical staining using the Ventana automated immunostainer BenchMark XT (Ventana Medical Systems, Tucson, AZ, USA).
The protein expression levels of CYP1A2, CYP3A4, and FBP1 were interpreted in a semi-quantitative manner. The expression of each marker was evaluated as positive when it was detected in ≥5% of tumor epithelial cells with moderate to strong intensity and was graded on a scale of 0-3 (0, <5%; 1, 5-10%; 2, 11-50%; 3, 51-100% of tumor epithelial cells).
4. Statistical analysis
Statistical analyses were performed using SPSS software version 23.0.0 (SPSS Inc., Chicago, IL, USA) or the R package (www.r-project.org). To compare expression of genes according to the differentiation and size of HCCs, we used the Mann-Whitney U test for continuous variables and the χ2 or Fisher exact test for categorical variables. The correlation analysis was performed by calculating the Pearson’s correlation coefficients and p-values. For comparison of gene expression during HBV related multistep hepatocarcinogenesis, we used the linear trend and log-linear model. Significance was set at P<0.05 for all tests.
RESULTS
1. mRNA levels of TGF-β signaling genes in B virus-associated human multistep hepatocarcinogenesis
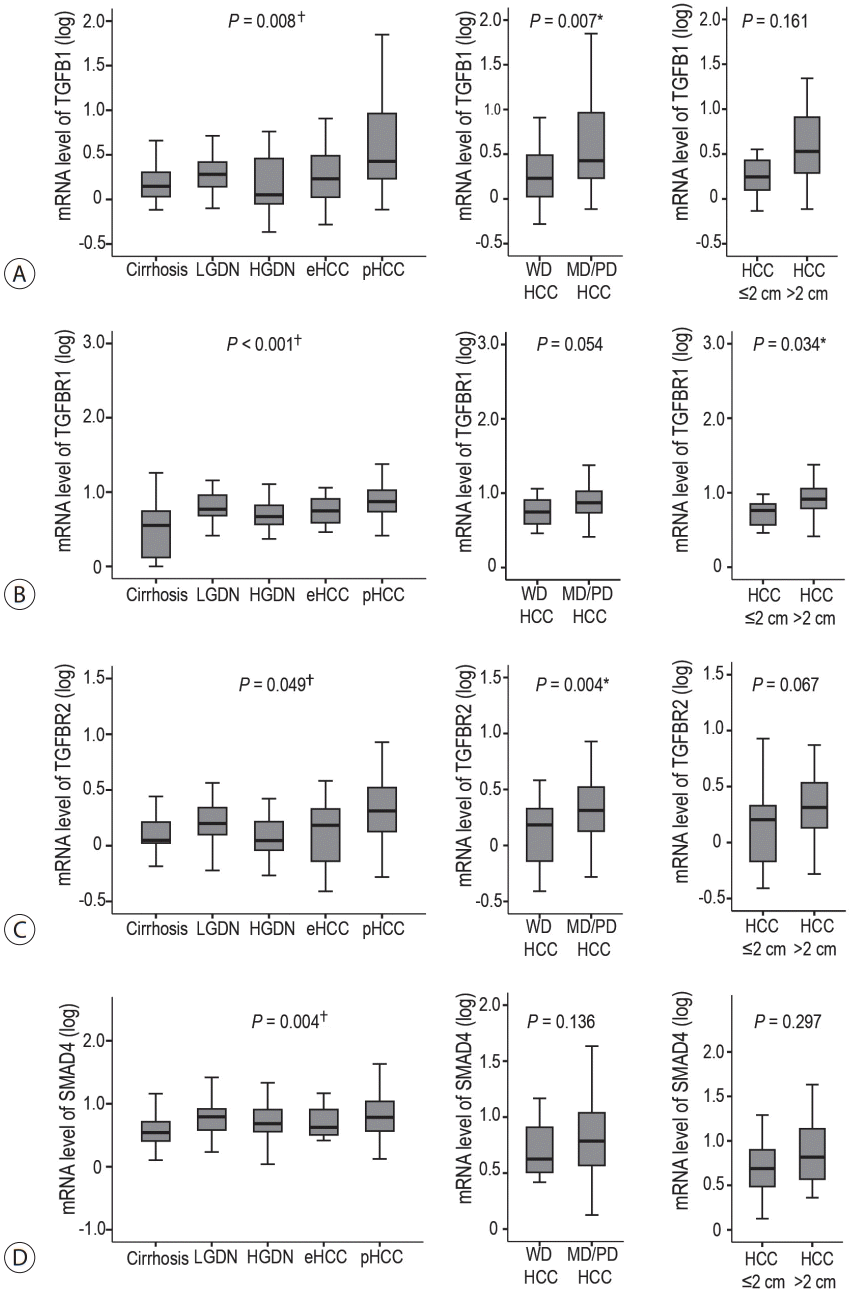
The mRNA levels of TGF-β signaling genes (TGFB1, TGFBR1, TGFBR2, and SMAD4) were evaluated in human HBV-related multistep hepatocarcinogenesis (Fig. 1). The expression of these genes gradually increased as the multistep hepatocarcinogenesis progressed toward HCC, showing the highest levels in pHCCs; this observation was statistically significant (all P<0.05). Additionally, the mRNA expression of TGF-β signatures was evaluated with respect to the differentiation and size of HBV-related HCCs. The mRNA levels of TGFB1 and TGFBR2 were significantly higher in moderately/poorly differentiated HCCs than in well-differentiated HCCs (both P<0.05), whereas the mRNA levels of TGFBR1 and SMAD4 showed no significant difference with respect to differentiation. Depending on the size of HBV-related HCCs, the mRNA levels of TGFBR1 were higher in HCCs >2 cm than those in HCCs ≤2 cm (P=0.034), and the mRNA levels of other TGF-β signaling genes showed no significant difference depending on the size of HCCs.
2. The expression of early and late responsive TGF-β signatures in B virus-associated human multistep hepatocarcinogenesis
Information regarding the early and late response genes was obtained from the reports of Coulouran et al.2; the expression of these genes was investigated in defined lesions of human multistep hepatocarcinogenesis.
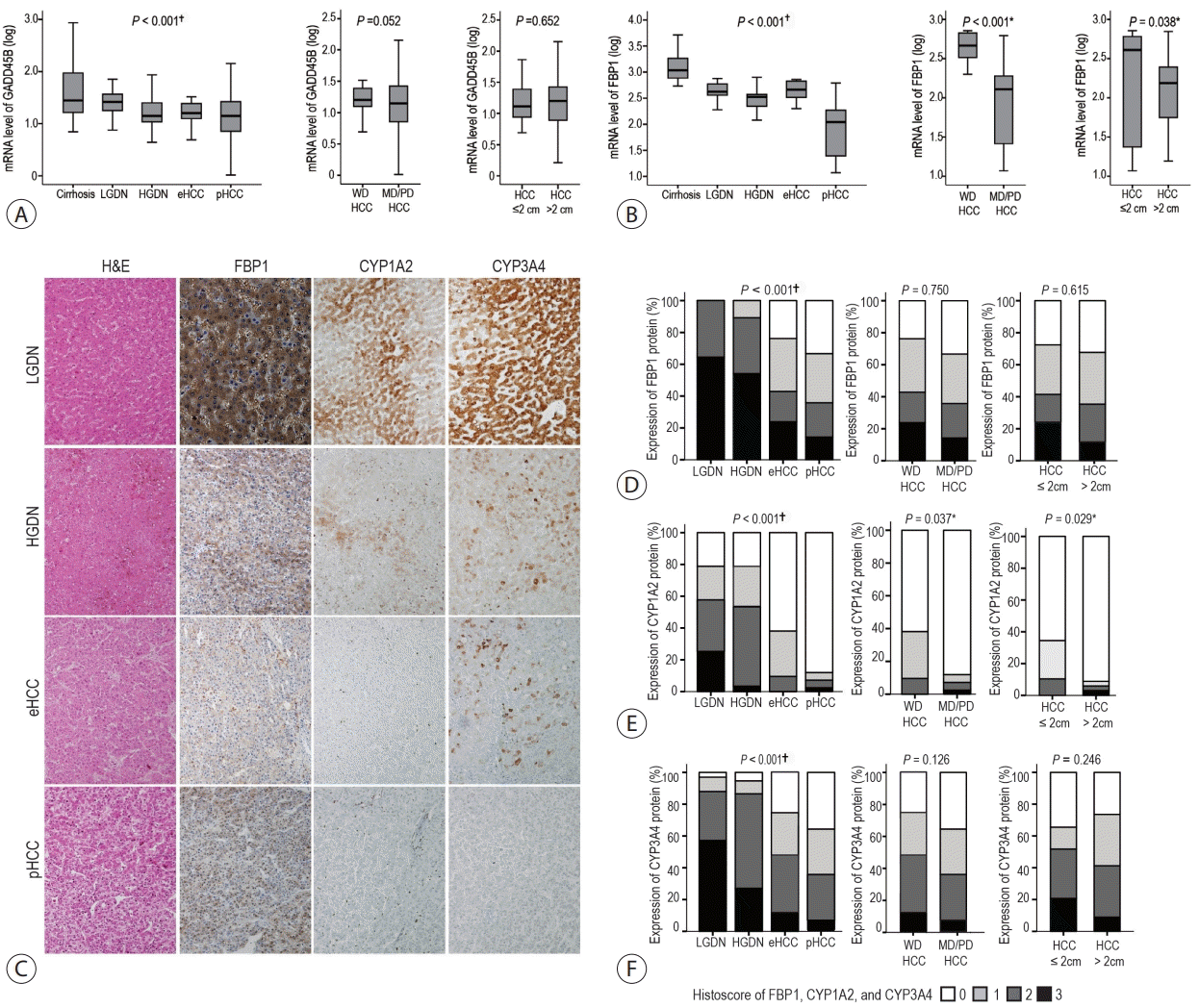
The mRNA levels of the early TGF-β responsive genes (GADD45B and FBP1) gradually decreased with the progression of the HBV-related multistep hepatocarcinogenesis (both P<0.001), and the lowest levels were observed in pHCCs (Fig. 2A, B). The mRNA expression of GADD45B and FBP1 was significantly higher in LGDNs than that in HGDNs (both P<0.001).
In addition, the mRNA levels of GADD45B and FBP1 were evaluated according to the differentiation and size of HBV-related HCCs. The mRNA levels of FBP1 but not of GADD45B, were lower in moderately/poorly differentiated HCCs than those in well differentiated HCCs (Fig. 2A, B). With respect to the size of HBV-related HCCs, the mRNA levels of FBP1 were lower in HCCs >2 cm compared with those in HCCs ≤2 cm (P=0.038) (Fig. 2B); the mRNA levels of GADD45B did not show a significant difference according to the size of HCCs (P=0.652) (Fig. 2A, B).
Next, the protein expression of FBP1, CYP1A2, and CYP3A4, a kind of early responsive TGF-β signature genes2, was evaluated by immunohistochemical staining (Fig. 2C). The expression of FBP1, CYP1A2, and CYP3A4 decreased during multistep hepatocarcinogenesis from the LGDN to pHCC stage, showing a significant linear trend (all P<0.001) (Fig. 2D). Additionally, the histoscores of CYP1A2 and CYP3A4 were significantly higher in LGDNs than in HGDNs (P=0.038 and P=0.023, respectively). Among the pHCCs, the histoscore of CYP1A2 was significantly lower in moderately/poorly differentiated HCCs than that in well differentiated HCCs (P=0.037), and it was significantly lower in large HCCs (>2 cm) than that in small HCCs (≤2 cm) (P=0.029) (Fig. 2E). FBP1 and CYP3A4 expression status showed no significant differences according to HCC differentiation and size (Fig. 2D, F).
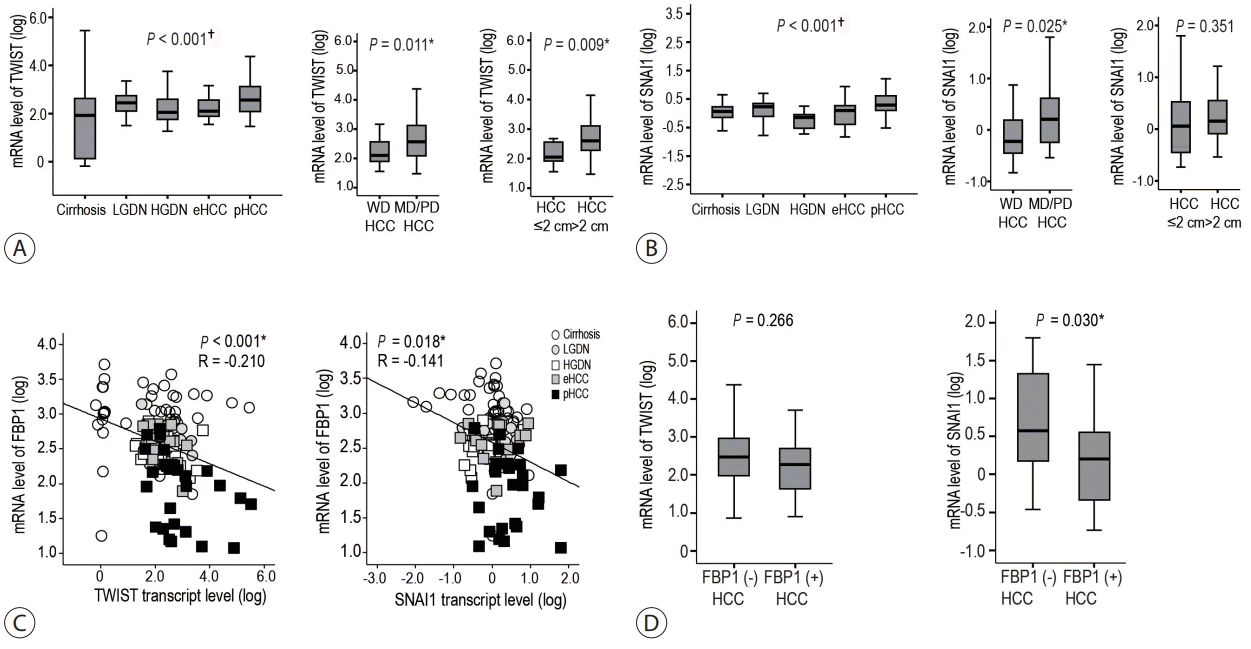
The mRNA levels of TWIST and SNAI1, which are late responsive TGF-β signature genes as well as major regulators of EMT, were evaluated. The transcripts of TWIST and SNAI1 showed a gradual increase in levels as HBV-related multistep hepatocarcinogenesis progressed toward pHCC, showing a significant linear trend (all P<0.05) (Fig. 3A, B). In addition, the mRNA expression of TWIST and SNAI1 was evaluated according to HCC differentiation and size. Transcripts of TWIST and SNAI1 showed significantly higher levels in moderately/poorly differentiated HCCs than those observed in well differentiated HCCs (both P<0.05) (Fig. 3A, B). According to the size of the HCCs, TWIST mRNA levels were higher in HCCs >2 cm than those in HCCs ≤2 cm (P=0.009), but not that in SNAI1 (Fig. 3A, B). Interestingly, FBP1 expression was negatively correlated with TWIST and SNAI1 expression (P<0.05) (Fig. 3C), and SNAI1 expression was higher in FBP1-negative HCCs than that in FBP1-positive HCCs (Fig. 3D).
3. Expression levels of early and late TGF-β gene signatures, according to the expression of stemness-related markers in B virus-associated human multistep hepatocarcinogenesis
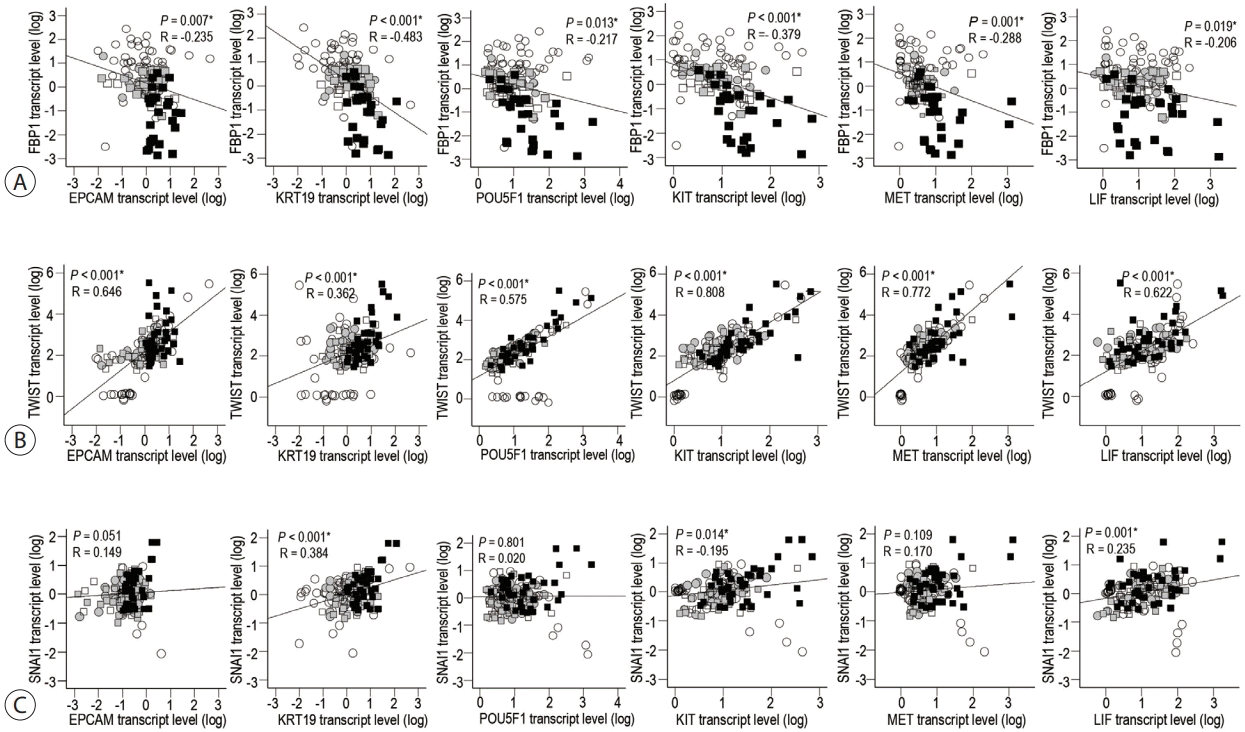
We previously reported that the expression of CSC features is significantly increased during the progression of B virus-associated multistep hepatocarcinogenesis.7 Using previously reported data, we evaluated the correlation between the early and late TGF-β signatures and stemness-related genes. The mRNA levels of TGF-β signatures (TGFB1 , TGFBR1 , TGFBR2, and SMAD4) were positively correlated with those of stemness-related genes (EPCAM, KRT19, POU5F1, MET and LIF) in HBV-related multistep hepatocarcinogenesis (all P<0.05) (Supplementary Fig. 1).
The levels of FBP1 , an early TGF-β-responsive gene, showed a reverse correlation with those of the stemness-related genes during the progression of hepatocarcinogenesis from cirrhosis to pHCC (all P<0.05) (Fig. 4A). As for the late TGF-β signatures, a positive correlation between TWIST mRNA levels and those of stemness-related genes was found in HBV-related hepatocarcinogenesis (all P<0.05) (Fig. 4B). SNAI1 mRNA levels showed a significant positive correlation with those of KRT19, KIT, and LIF in HBV-related stepwise hepatocarcinogenesis (all P<0.05) (Fig. 4C).
DISCUSSION
The purpose of this study was to investigate the expression of early and late TGF-β signatures during multistep hepatocarcinogenesis in order to gain a better understanding of the dual functions of TGF-β, including its tumor-suppressive and oncogenic roles.10,11 In this study, the transcript levels of TGF-β signatures, including TGFB1, TGFBR1, TGFBR2 and SMAD4 gradually increased with the progression of B virus-associated human multistep hepatocarcinogenesis and showed the highest levels in pHCCs. Among the HCCs, these genes were more highly expressed in advanced HCCs showing less differentiation and a large size.
GADD45B is an early response gene with a TGF-β signature and induced by TGF-β in a SMAD-dependent manner. It has been identified as a positive regulator of TGF-β-induced apoptosis; therefore, activation of GADD45β is considered to prevent the propagation of damaged cells by initiating cell growth arrest and subsequent apoptosis after exposure to genotoxins.12 In our study, GADD45B mRNA levels gradually decreased with the progression of HBV-related multistep hepatocarcinogenesis with the highest level in cirrhosis. It was actually higher in LGDNs than in HGDNs. These results indicate that GADD45β may have a tumor-suppressive or -protective role by preventing the malignant progression of DNs in the early stages of hepatocarcinogenesis.
FBP1 is one of early response genes with a TGF-β signature and is associated with liver-specific functions, such as glucose and lipid metabolism or detoxification.2 In this study, protein and mRNA levels significantly decreased according to the progression of multistep hepatocarcinogenesis, and the FBP1 transcript levels were lower in more advanced HCCs with less differentiation and larger size. Accordingly, downregulation of FBP1 has been reported to be strongly correlated with aggressive characteristics and poor prognosis in HCCs.13
CYP1A2 and CYP3A4 are early response genes with a TGF-β signature and are related to antioxidant responses. CYP1A2 and CYP3A4 protein expression significantly decreased with the progression of multistep hepatocarcinogenesis, and the histoscores of CYP1A2 and CYP3A4 were significantly higher in LGDNs than in HGDNs. Among the HCCs, CYP1A2 expression was lower in more advanced HCCs, with less differentiation and a large size. Previously, CYP1A2 expression was reported to be lower in HCC than that in the adjacent liver14,15, and CYP1A2 was found to suppress HCC via the conversion of E2 to 2-ME that induces cell cycle arrest and apoptosis of HCCs.15,16 Additionally, CYP1A2 expression is inhibited by TGF-β1 in human and rat hepatocytes.17,18 Taken together, the early responsive TGF-β genes, such as GADD45B, FBP1, CYP1A2, and CYP3A4, are considered to have tumor-suppressive roles in the early stages of multistep hepatocarcinogenesis. In addition, a followup study of histologically proven LGDNs and HGDNs revealed that the hazard ratios for transformation to HCC were 2.96 and 16.80, respectively.19 Only a minority of LGDNs became malignant, and those transforming were mostly HGDN.20 This is considered to be related to the tumor-protective roles of early responsive genes of the TGF-β signature that are highly expressed in LGDNs compared to that in HGDNs.
Concerning the late response genes of TGF-β gene signatures, the mRNA levels of TWIST and SNAI1 gradually increased with the progression of hepatocarcinogenesis, showing the highest levels in pHCCs. Among HCCs, the mRNA levels of TWIST and SNAI1 were higher in the less differentiated and large HCCs. They were also positively correlated with the mRNA levels of TGF-β signaling genes in this study. Additionally, overexpression of Twist has been reported to be correlated with the invasive and metastatic abilities of HCCs through the induction of EMT.21 Snail expression was previously reported to be associated with matrix metalloproteinases expression and HCC invasiveness.22 Therefore, the late TGF-β signature and EMT are considered to be more involved in the late stages of multistep hepatocarcinogenesis.
The aggressive biological behavior and poor prognosis of HCC have been reported in those expressing stemness markers such as K19 and EpCAM.5,23,24 Previous studies have revealed that the activation of EMT in epithelial cells leads to the acquisition of stemness properties, which may serve the generation of CSCs in the context of cancer.3,4 We have previously reported that stemness is enriched in the progression of multistep hepatocarcinogenesis, along with stromal alterations.7 In addition, this study showed that stemness marker expression was well correlated with that of TWIST and SNAI1, which were upregulated by TGF-β signatures in the late stage of B virus-associated human multistep hepatocarcinogenesis.
Unlike the late-responsive TGF-β genes, the expression of FBP1 is inversely correlated with the expression of stemness markers in HBV-related multistep hepatocarcinogenesis. Overexpression of FBP1 in basal-like breast cancer was reported to inhibit tumor sphere formation and reduce the percentage of CD44high/CD24low/EpCAM-positive population, whereas knockdown of FBP1 in luminal cell lines resulted in the opposite phenotypes.25 FBP1 is downregulated by methylation of the promoter region by Snail or Zeb1.25,26 Accordingly, in this study, the expression of FBP1 was inversely correlated with the expression of SNAI1, and SNAI1 levels were lower in FBP1-positive HCCs when compared to FBP1-negative HCCs. Taken together, the reduced expression of FBP1 might be associated with EMT and stemness features.
This study focused on HBV-related multistep hepatocarcinogenesis. There is molecular diversity in HCCs according to their etiology. HCCs with a progenitor cell phenotype, including K19-positive HCCs, are more prevalent in HBV-related HCCs and it has been reported that in these HCCs, there is an increased TGF-β signaling.27 Therefore, our study samples of HBV-related hepatocarcinogenesis were considered a good cohort to evaluate the dual role of TGF-β signatures. Further studies are needed to evaluate its role in multistep hepatocarcinogenesis related to other etiologies, including hepatitis C virus, alcohol, and metabolic diseases.
In conclusion, TGF-β signatures and the late response genes of TGF-β are upregulated, whereas the expression of the early response genes of TGF-β are gradually decreased during HBV-related multistep hepatocarcinogenesis. Thus, enrichment of the late responsive signatures of TGF-β with the induction of stemness is considered to be involved in the progression of multistep hepatocarcinogenesis. In contrast, the early responsive genes of TGF-β might have tumor-suppressive roles in the early stages of multistep hepatocarcinogenesis.
 XML Download
XML Download